

Is the Phytotoxic Effect of Digestive Fluid of Nepenthes x ventrata on Tomato Related to Reactive Oxygen Species?

Abstract
:1. Introduction
2. Results
2.1. Electrical Conductivity
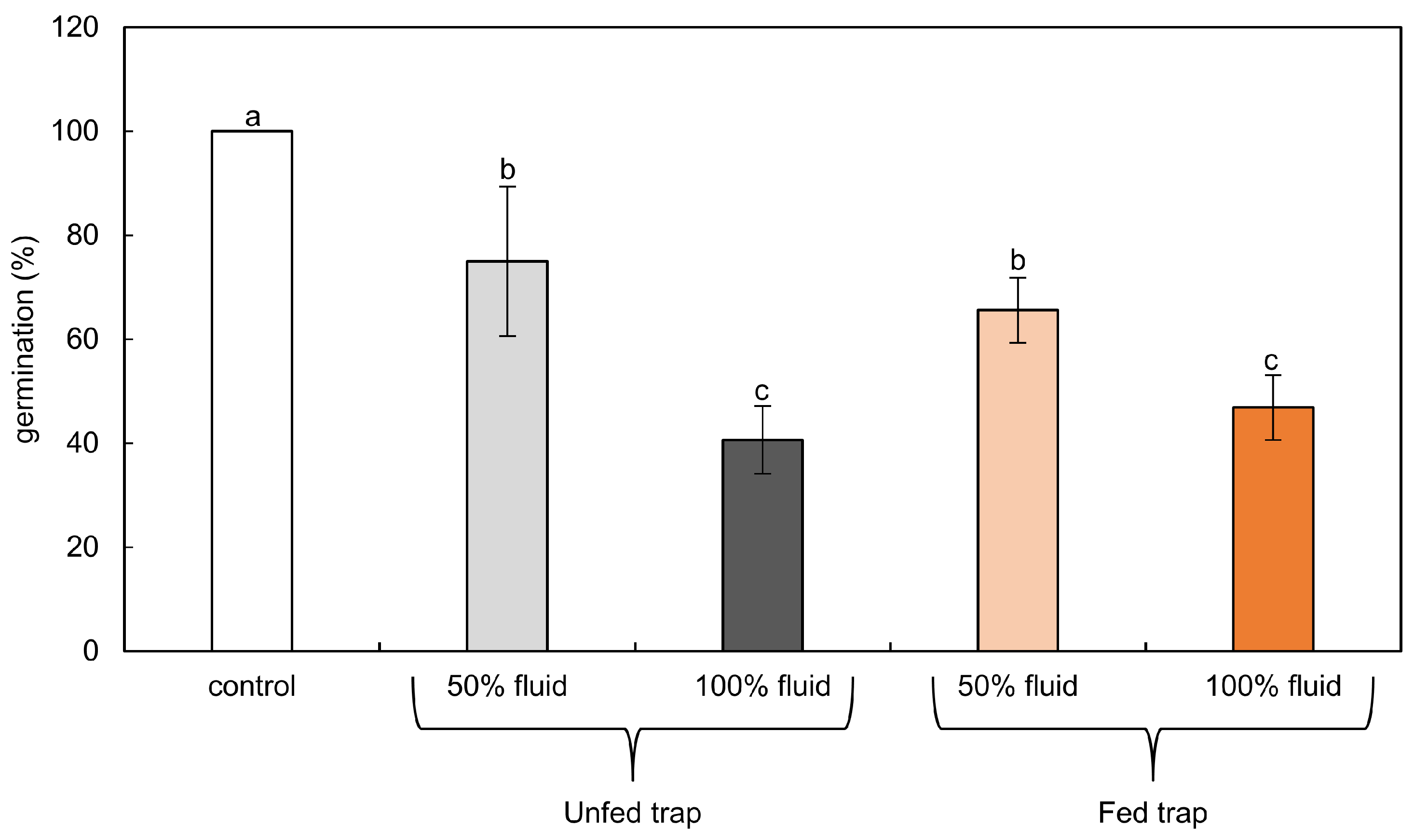
2.2. Germination Tests
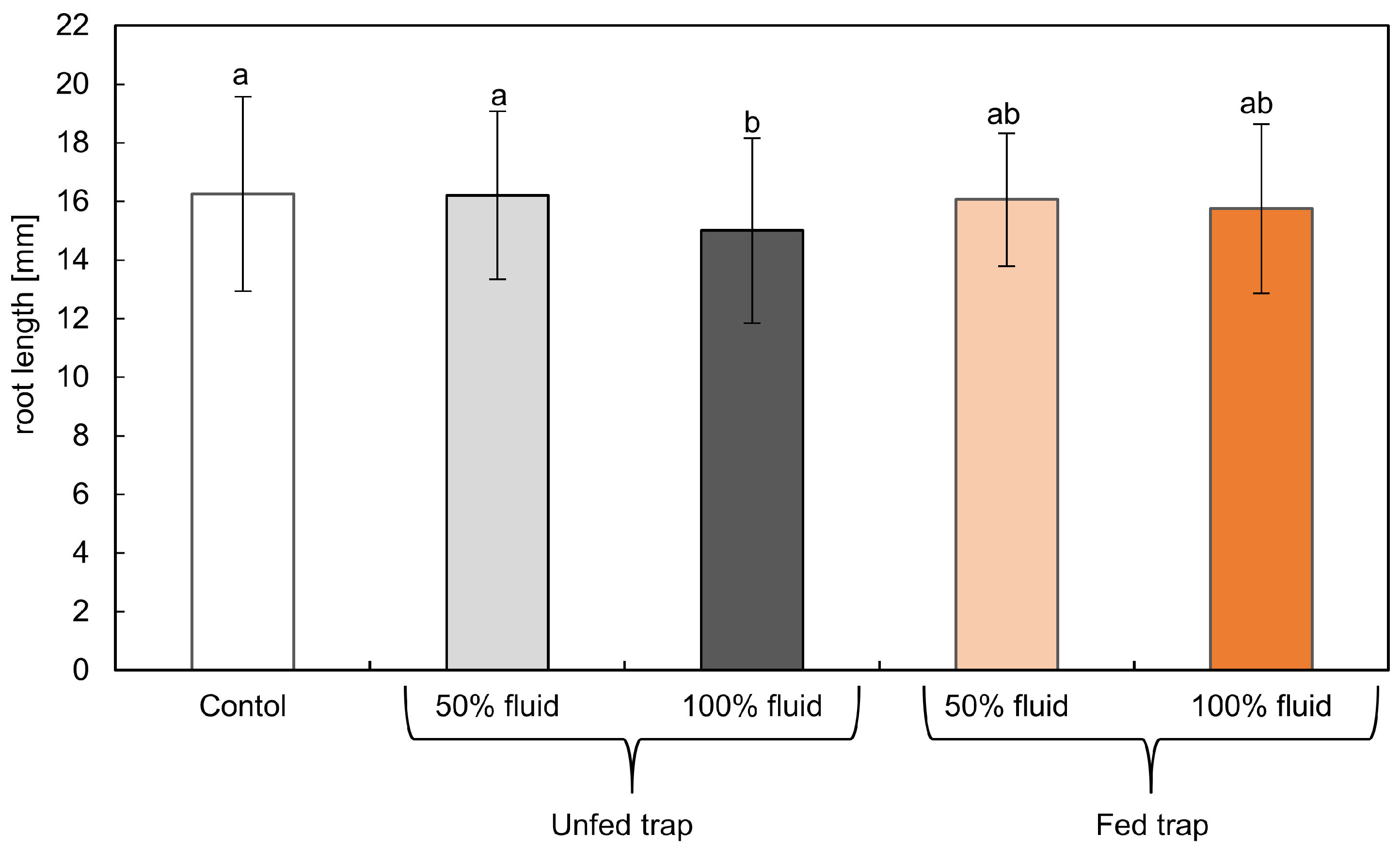
2.3. Root Growth Biotest
2.4. Test of Root Cell Viability
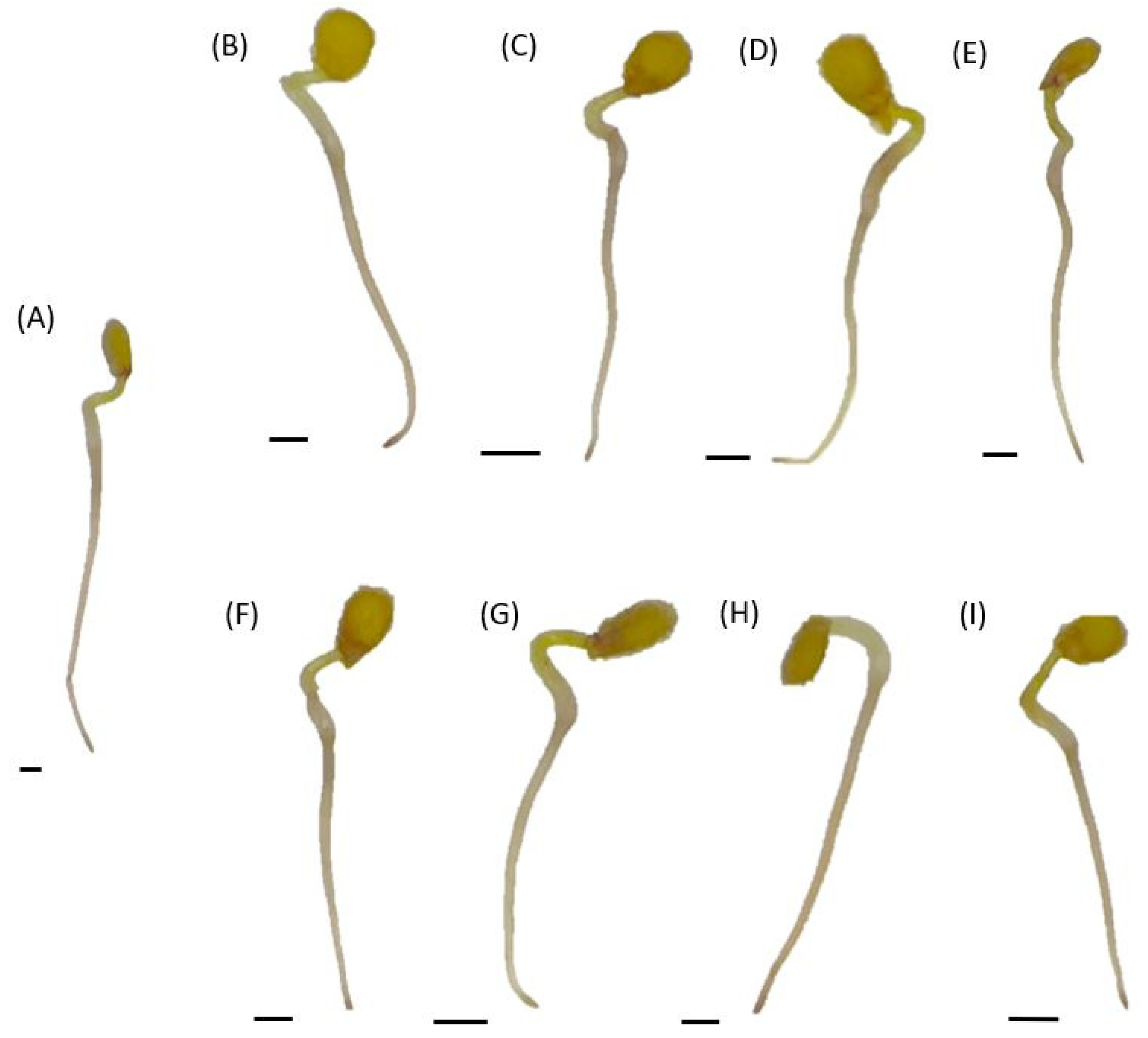
2.5. In Situ H2O2 Localization in Tomato Seedlings Cultured
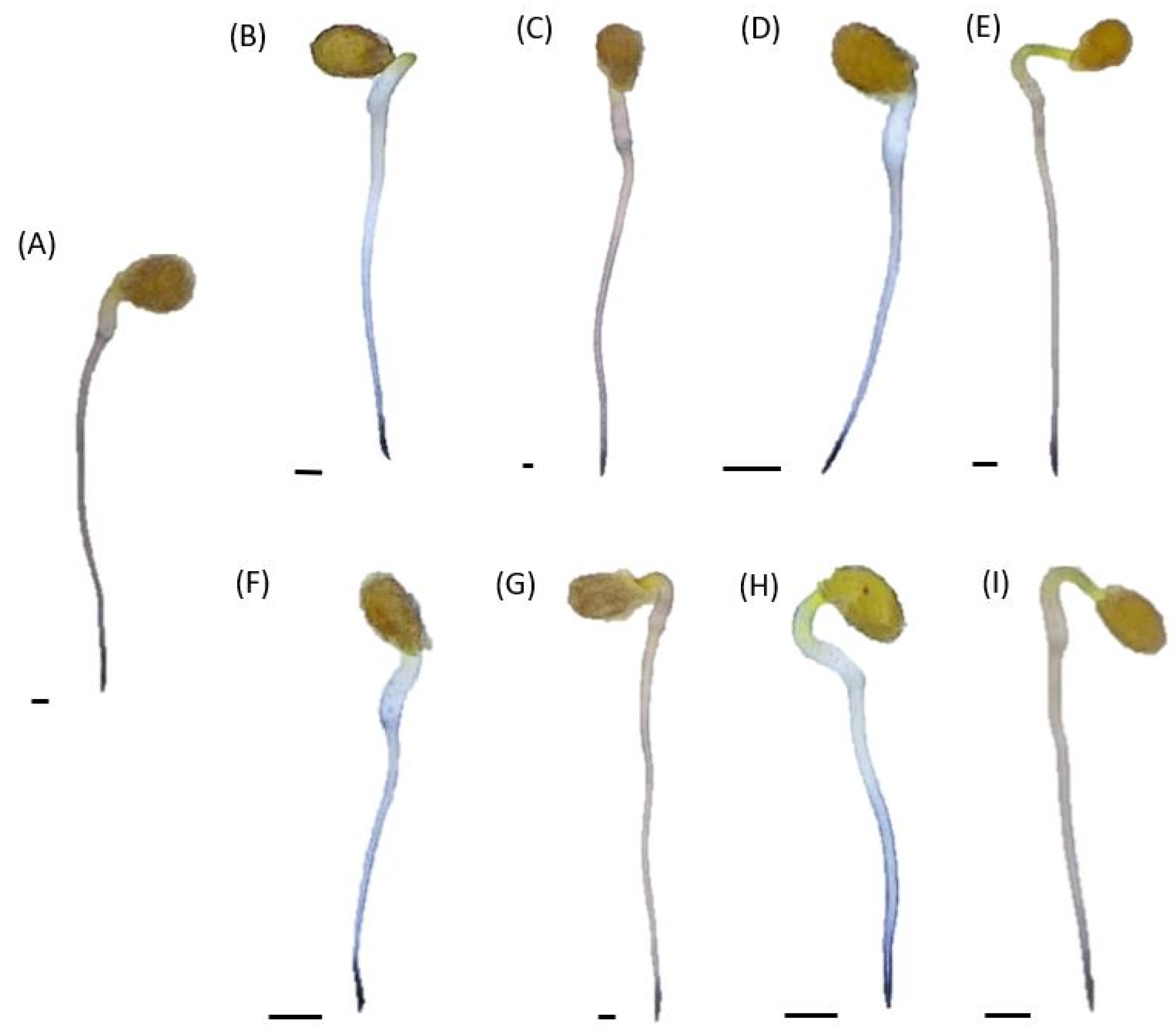
2.6. In Situ O2•− Localization
3. Discussion
4. Materials and Methods
4.1. Experimental Model
4.2. Germination Tests
4.3. Test of Root Cell Viability
4.4. In Situ H2O2 Localization
4.5. In Situ O2•− Localization
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clarke, C. Nepenthes of Sumatra and Peninsular Malaysia; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2001. [Google Scholar]
- Adlassnig, W.; Peroutka, M.; Lendl, T. Traps of Carnivorous Pitcher Plants as a Habitat: Composition of the Fluid, Biodiversity and Mutualistic Activities. Ann. Bot. 2011, 107, 181–194. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Fukusaki, E.I.; Kobayashi, A. Aspartic Proteinases Are Expressed in Pitchers of the Carnivorous Plant Nepenthes Alata Blanco. Planta 2002, 214, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, H.; Pnini-Cohen, S.; Schuster, S.; Movtchan, A.; Zilberstein, A. Isolation and Characterization of Chitinase Genes from Pitchers of the Carnivorous Plant Nepenthes Khasiana. J. Exp. Bot. 2006, 57, 2775–2784. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J.; Adamec, L.; Lichtscheidl, I.K.; Peroutka, M.; Adlassnig, W.; Vrba, J. Fluorescence Labelling of Phosphatase Activity in Digestive Glands of Carnivorous Plants. Plant Biol. 2006, 8, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, P.; Hogan, J. Cloning and Characterization of a Ribonuclease, a Cysteine Proteinase, and an Aspartic Proteinase from Pitchers of the Carnivorous Plant Nepenthes Ventricosa Blanco. Int. J. Plant Sci. 2006, 167, 239–248. [Google Scholar] [CrossRef]
- Athauda, S.B.P.; Matsumoto, K.; Rajapakshe, S.; Kuribayashi, M.; Kojima, M.; Kubomura-Yoshida, N.; Iwamatsu, A.; Shibata, C.; Inoue, H.; Takahashi, K. Enzymic and Structural Characterization of Nepenthesin, a Unique Member of a Novel Subfamily of Aspartic Proteinases. Biochem. J. 2004, 381, 295–306. [Google Scholar] [CrossRef]
- Hatano, N.; Hamada, T. Proteome Analysis of Pitcher Fluid of the Carnivorous Plant Nepenthes alata. J. Proteome Res. 2008, 7, 809–816. [Google Scholar] [CrossRef]
- Mithöfer, A. Carnivorous Pitcher Plants: Insights in an Old Topic. Phytochemistry 2011, 72, 1678–1682. [Google Scholar] [CrossRef]
- Eilenberg, H.; Pnini-Cohen, S.; Rahamim, Y.; Sionov, E.; Segal, E.; Carmeli, S.; Zilberstein, A. Induced Production of Antifungal Naphthoquinones in the Pitchers of the Carnivorous Plant Nepenthes khasiana. J. Exp. Bot. 2010, 61, 911–922. [Google Scholar] [CrossRef]
- Buch, F.; Rott, M.; Rottloff, S.; Paetz, C.; Hilke, I.; Raessler, M.; Mithöfer, A. Secreted Pitfall-Trap Fluid of Carnivorous Nepenthes Plants Is Unsuitable for Microbial Growth. Ann. Bot. 2013, 111, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Likhitwitayawuid, K.; Kaewamatawong, R.; Ruangrungsi, N.; Krungkrai, J. Antimalarial Naphthoquinones from Nepenthes thorelii. Planta Med. 1998, 64, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Sandur, S.K.; Ichikawa, H.; Sethi, G.; Ahn, K.S.; Aggarwal, B.B. Plumbagin (5-Hydroxy-2-Methyl-1,4-Naphthoquinone) Suppresses NF-ΚB Activation and NF-ΚB-Regulated Gene Products Through Modulation of P65 and IκBα Kinase Activation, Leading to Potentiation of Apoptosis Induced by Cytokine and Chemotherapeutic Agents. J. Biol. Chem. 2006, 281, 17023–17033. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.-S.; Lee, S.-K.; Cha, B.-J. Antifungal Activity of Plumbagin Purified from Leaves of Nepenthes Ventricosa x Maxima against Phytopathogenic Fungi. Plant Pathol. J. 2007, 23, 113–115. [Google Scholar] [CrossRef]
- Wal, A.; Staszek, P.; Pakula, B.; Paradowska, M.; Krasuska, U. ROS and RNS Alterations in the Digestive Fluid of Nepenthes × Ventrata Trap at Different Developmental Stages. Plants 2022, 11, 3304. [Google Scholar] [CrossRef] [PubMed]
- Gaume, L.; Forterre, Y. A Viscoelastic Deadly Fluid in Carnivorous Pitcher Plants. PLoS ONE 2007, 2, 1185. [Google Scholar] [CrossRef]
- Macías, F.A.; Molinillo, J.M.; Varela, R.M.; Galindo, J.C. Allelopathy—A Natural Alternative for Weed Control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A. Sitophilus Granarius Responses to Blends of Five Groups of Cereal Kernels and One Group of Plant Volatiles. J. Stored Prod. Res. 2015, 63, 63–66. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A.; Krasińska, A.; Wrzesińska, D.; Delaney, K.J. Volatile Organic Compounds Released by Rumex Confertus Following Hypera Rumicis Herbivory and Weevil Responses to Volatiles. J. Appl. Entomol. 2016, 140, 308–316. [Google Scholar] [CrossRef]
- Staszek, P.; Krasuska, U.; Wal, A.; Zak, J.; Gniazdowska, A. NO and Metabolic Reprogramming under Phytotoxicity Stress. In Nitric Oxide in Plant Biology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 297–318. [Google Scholar]
- Chan, X.-Y.; Hong, K.-W.; Yin, W.-F.; Chan, K.-G. Microbiome and Biocatalytic Bacteria in Monkey Cup (Nepenthes Pitcher) Digestive Fluid. Sci. Rep. 2016, 6, 20016. [Google Scholar] [CrossRef]
- Bittleston, L.S.; Wolock, C.J.; Yahya, B.E.; Chan, X.Y.; Chan, K.G.; Pierce, N.E.; Pringle, A. Convergence between the Microcosms of Southeast Asian and North American Pitcher Plants. Elife 2018, 7, e36741. [Google Scholar] [CrossRef]
- Hossain, M.S.; Rahman, G.K.M.M.; Solaiman, A.R.M.; Alam, M.S.; Rahman, M.M.; Mia, M.A.B. Estimating Electrical Conductivity for Soil Salinity Monitoring Using Various Soil-Water Ratios Depending on Soil Texture. Commun. Soil Sci. Plant Anal. 2020, 51, 635–644. [Google Scholar] [CrossRef]
- Huliusz, P. Quantitative and Qualitative Diferentiation of Soil Salinity in Poland. Ber. Dtsch. Bodenkd. Ges. 2008, 1, 1–4. [Google Scholar]
- Gonçalves, S.; Ferraz, M.; Romano, A. Phytotoxic Properties of Drosophyllum Lusitanicum Leaf Extracts and Its Main Compound Plumbagin. Sci. Hortic. 2009, 122, 96–101. [Google Scholar] [CrossRef]
- Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Castellano, D.; Molinillo, J.M.G. Search for a Standard Phytotoxic Bioassay for Allelochemicals. Selection of Standard Target Species. J. Agric. Food Chem. 2000, 48, 2512–2521. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Bogatek, R. Allelopathic Interactions between Plants. Multi Site Action of Allelochemicals. Acta Physiol. Plant 2005, 27, 395–407. [Google Scholar] [CrossRef]
- Staszek, P.; Krasuska, U.; Ciacka, K.; Gniazdowska, A. ROS Metabolism Perturbation as an Element of Mode of Action of Allelochemicals. Antioxidants 2021, 10, 1648. [Google Scholar] [CrossRef]
- Soltys, D.; Rudzińska-Langwald, A.; Gniazdowska, A.; Wiśniewska, A.; Bogatek, R. Inhibition of Tomato (Solanum Lycopersicum L.) Root Growth by Cyanamide Is Due to Altered Cell Division, Phytohormone Balance and Expansin Gene Expression. Planta 2012, 236, 1629–1638. [Google Scholar] [CrossRef]
- Graña, E.; Sotelo, T.; Díaz-Tielas, C.; Araniti, F.; Krasuska, U.; Bogatek, R.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Citral Induces Auxin and Ethylene-Mediated Malformations and Arrests Cell Division in Arabidopsis Thaliana Roots. J. Chem. Ecol. 2013, 39, 271–282. [Google Scholar] [CrossRef]
- Haugland, E.; Brandsaeter, L.O. Experiments on Bioassay Sensitivity in the Study of Allelopathy. J. Chem. Ecol. 1996, 22, 1845–1859. [Google Scholar] [CrossRef]
- Shekari, F.; Shekari, F.; Najafi, J.; Abassi, A.; Radmanesh, Z.; Bones, A.M. Phytotoxic Effects of Catnip (Nepeta Meyeri Benth.) on Early Growth Stages Development and Infection Potential of Field Dodder (Cuscuta Campestris Yunck). Plants 2022, 11, 2629. [Google Scholar] [CrossRef]
- Oracz, K.; Voegele, A.; Tarkowská, D.; Jacquemoud, D.; Turečková, V.; Urbanová, T.; Strnad, M.; Sliwinska, E.; Leubner-Metzger, G. Myrigalone A Inhibits Lepidium Sativum Seed Germination by Interference with Gibberellin Metabolism and Apoplastic Superoxide Production Required for Embryo Extension Growth and Endosperm Rupture. Plant Cell Physiol. 2012, 53, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Bouteau, H.E.-M.; Farrant, J.M.; Cooper, K.; Belghazi, M.; Job, C.; Job, D.; Corbineau, F.; Bailly, C. ROS Production and Protein Oxidation as a Novel Mechanism for Seed Dormancy Alleviation. Plant J. 2007, 50, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Inderjit; Weston, L.A.; Duke, S.O. Challenges, Achievements and Opportunities in Allelopathy Research. J. Plant Interact. 2005, 1, 69–81. [Google Scholar] [CrossRef]
- Williams, R.; Hoagland, R. Bioassays- Useful Tools for the Study of Allelopathy. In Allelopathy; CRC Press: Boca Raton, FL, USA, 2003; pp. 315–351. [Google Scholar]
- Gniazdowska, A.; Krasuska, U.; Andrzejczak, O.; Soltys, D. Allelopathic Compounds as Oxidative Stress Agents: Yes or NO. In Reactive Oxygen and Nitrogen Species Signaling and Communication in Plants, Signaling and Communication in Plants; Springer: Cham, Switzerland, 2015; Volume 23, pp. 155–176. [Google Scholar]
- Krasuska, U.; Andrzejczak, O.; Staszek, P.; Borucki, W.; Gniazdowska, A. Toxicity of Canavanine in Tomato (Solanum Lycopersicum L.) Roots Is Due to Alterations in RNS, ROS and Auxin Levels. Plant Physiol. Biochem. 2016, 103, 84–95. [Google Scholar] [CrossRef]
- Baker, C.; Mock, M. An Improved Method for Monitoring Cell Death in Cell Suspension and Leaf Disc Assays Using Evans Blue. Plant Cell Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Ciacka, K.; Tyminski, M.; Gniazdowska, A.; Krasuska, U. Cold Stratification-Induced Dormancy Removal in Apple (Malus Domestica Borkh.) Seeds Is Accompanied by an Increased Glutathione Pool in Embryonic Axes. J. Plant Physiol. 2022, 274, 153736. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Electrical Conductivity [mS cm−1] | ||
|---|---|---|---|
| Control | 6.29 ± 0.07 c | 0.021 ± 0.017 A | |
| Crude fluid | |||
| Unfed trap | 50% fluid | 5.96 ± 0.17 bc | 0.786 ± 0.078 B |
| 100% fluid | 5.82 ± 0.03 b | 1.333 ± 0.012 C | |
| Fed trap | 50% fluid | 3.78 ± 0.19 a | 0.704 ± 0.042 B |
| 100% fluid | 3.38 ± 0.08 a | 1.247 ± 0.017 C | |
| Heat-treated fluid | |||
| Unfed trap | 50% fluid | 6.09 ± 0.29 bc | 0.771 ± 0.026 B |
| 100% fluid | 5.54 ± 0.10 b | 1.364 ± 0.052 C | |
| Fed trap | 50% fluid | 3.83 ± 0.31 a | 0.630 ± 0.040 B |
| 100% fluid | 3.36 ± 0.16 a | 1.127 ± 0.039 C | |
| Evans Blue Uptake to the Roots [mg dye g−1 FW] | Cell Damage [%] | ||
|---|---|---|---|
| Positive control (heat-treated roots) | 0.476 ± 0.011 c | 100 E | |
| Control (seedlings grown in water) | 0.086 ± 0.021 a | 18.1 A | |
| Crude fluid | |||
| Unfed trap | 50% fluid | 0.122 ± 0.064 b | 25.7 BC |
| 100% fluid | 0.151 ± 0.053 b | 31.8 D | |
| Fed trap | 50% fluid | 0.100 ± 0.048 b | 20.9 B |
| 100% fluid | 0.140 ± 0.079 b | 29.3 BC | |
| Heat-treated fluid | |||
| Unfed trap | 50% fluid | 0.109 ± 0.056 b | 22.9 BC |
| 100% fluid | 0.113 ± 0.055 b | 23.7 D | |
| Fed trap | 50% fluid | 0.135 ± 0.008 b | 28.4 B |
| 100% fluid | 0.149 ± 0.084 b | 31.3 BC | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staszek, P.; Piekarniak, M.; Wal, A.; Krasuska, U.; Gniazdowska, A. Is the Phytotoxic Effect of Digestive Fluid of Nepenthes x ventrata on Tomato Related to Reactive Oxygen Species? Plants 2023, 12, 755. https://doi.org/10.3390/plants12040755
Staszek P, Piekarniak M, Wal A, Krasuska U, Gniazdowska A. Is the Phytotoxic Effect of Digestive Fluid of Nepenthes x ventrata on Tomato Related to Reactive Oxygen Species? Plants. 2023; 12(4):755. https://doi.org/10.3390/plants12040755
Chicago/Turabian StyleStaszek, Pawel, Maciej Piekarniak, Agnieszka Wal, Urszula Krasuska, and Agnieszka Gniazdowska. 2023. "Is the Phytotoxic Effect of Digestive Fluid of Nepenthes x ventrata on Tomato Related to Reactive Oxygen Species?" Plants 12, no. 4: 755. https://doi.org/10.3390/plants12040755