
Characterization of a Disease-Suppressive Isolate of Lysobacter enzymogenes with Broad Antagonistic Activity against Bacterial, Oomycetal and Fungal Pathogens in Different Crops
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Screening of Different Lysobacter sp. Isolates
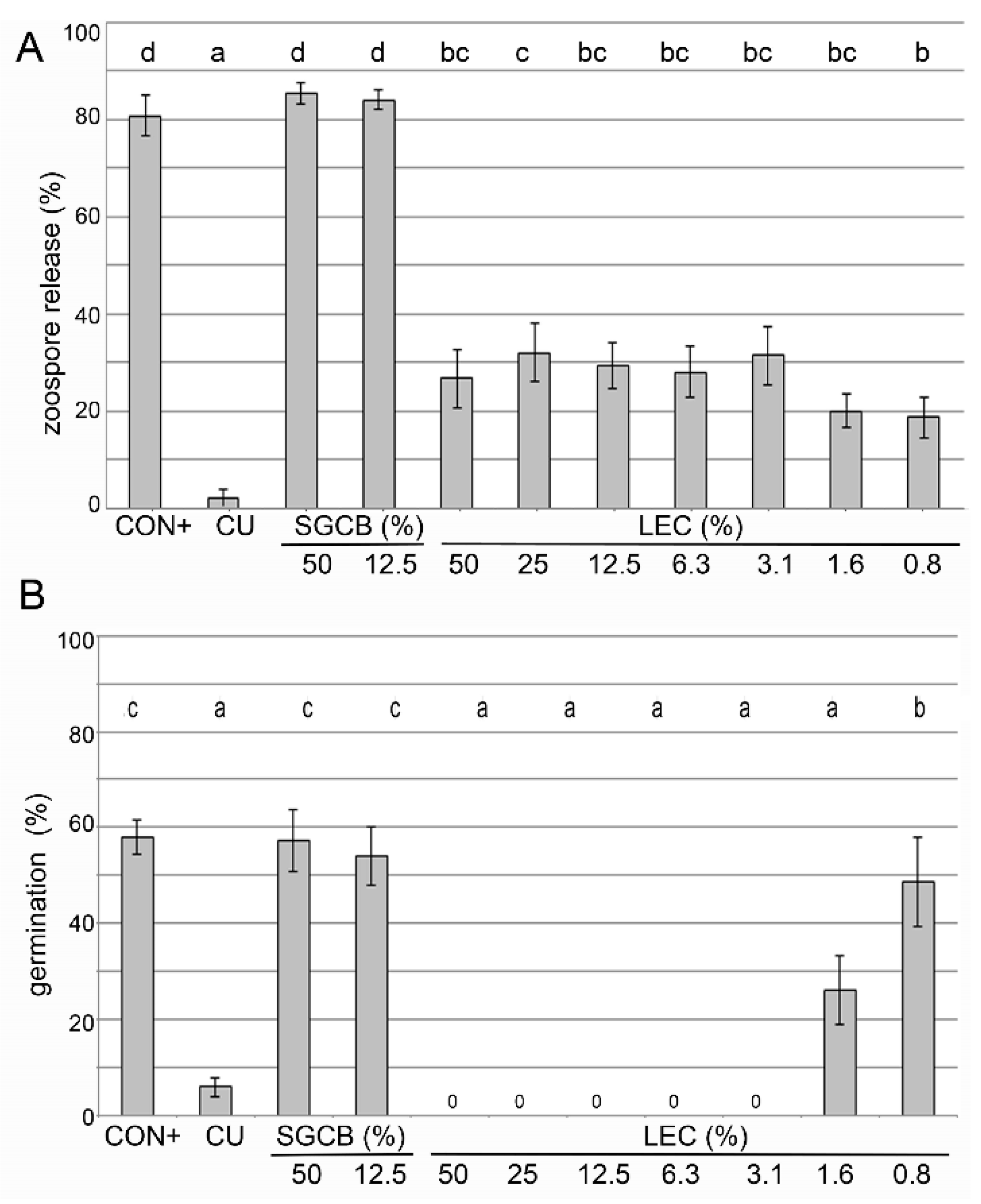
2.2. In Vitro Activity of Lysobacter enzymogenes Isolate LEC
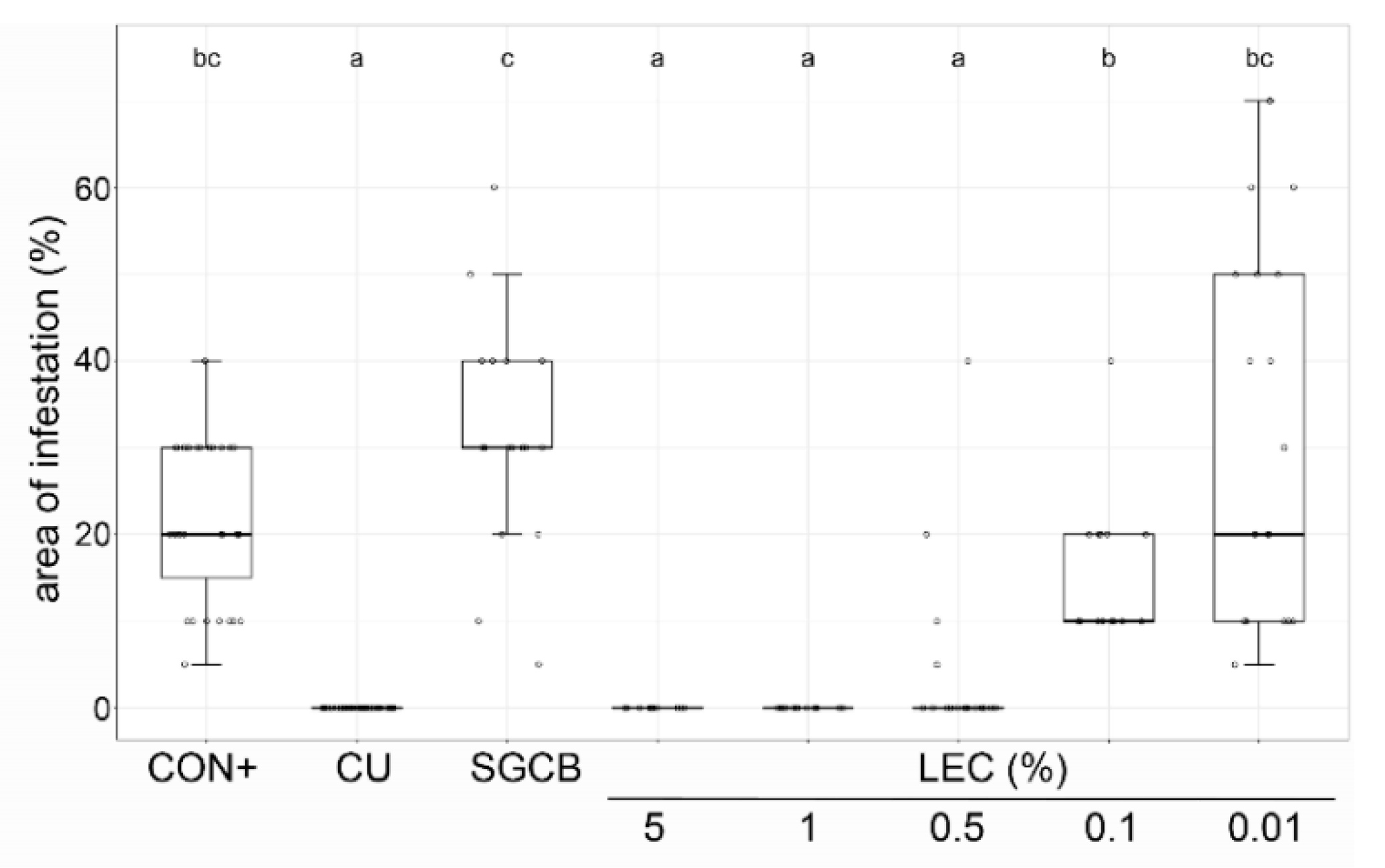
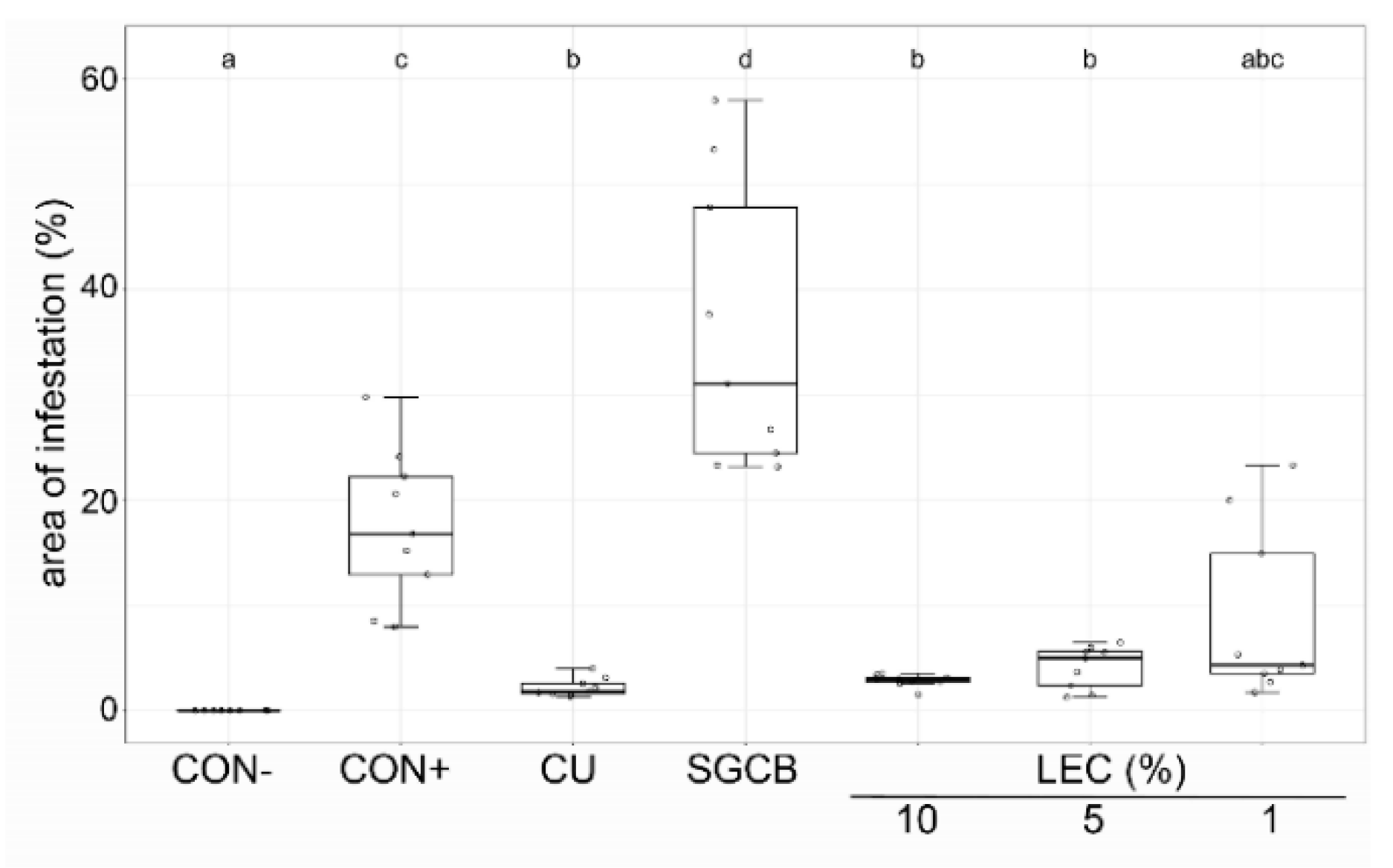
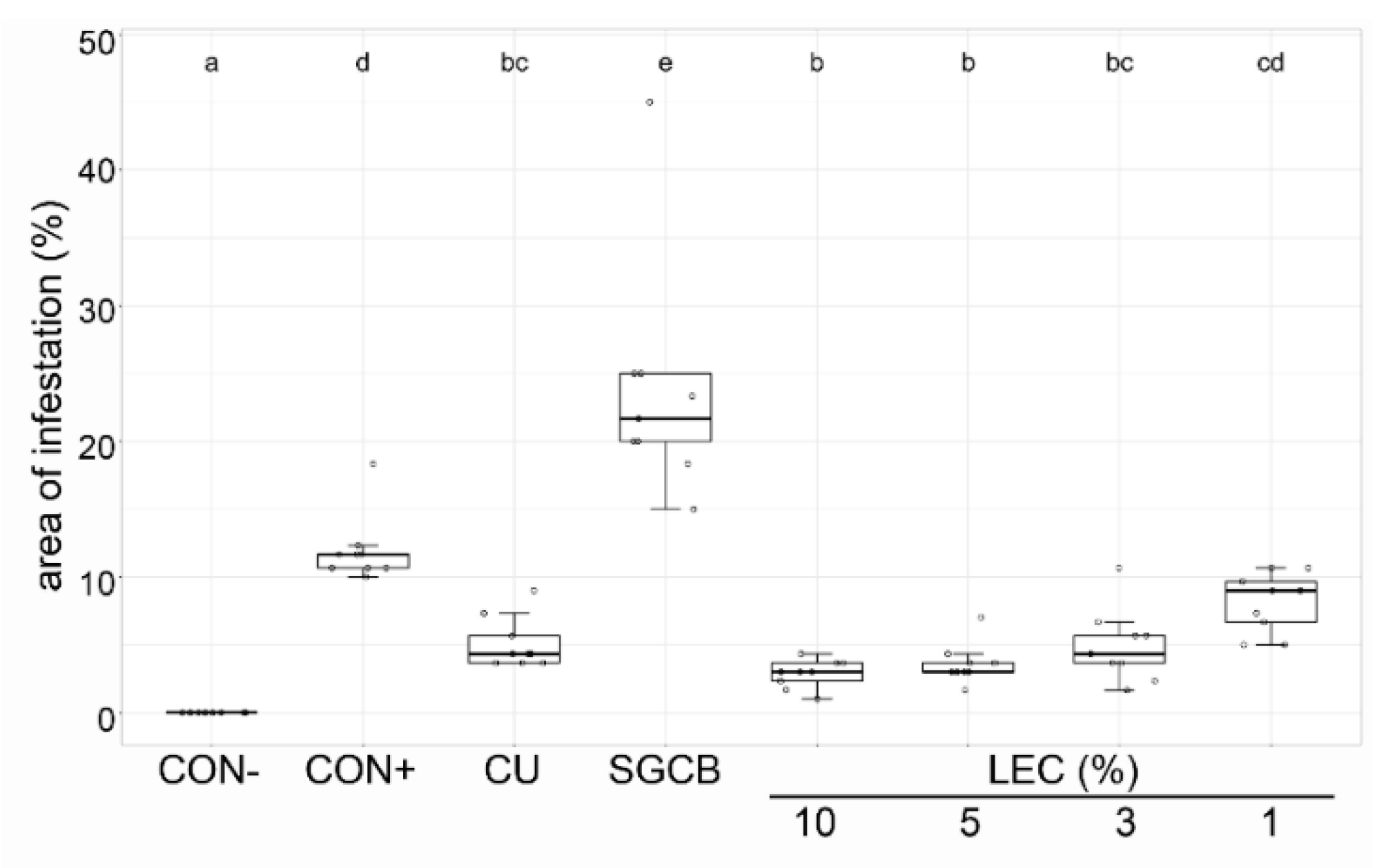
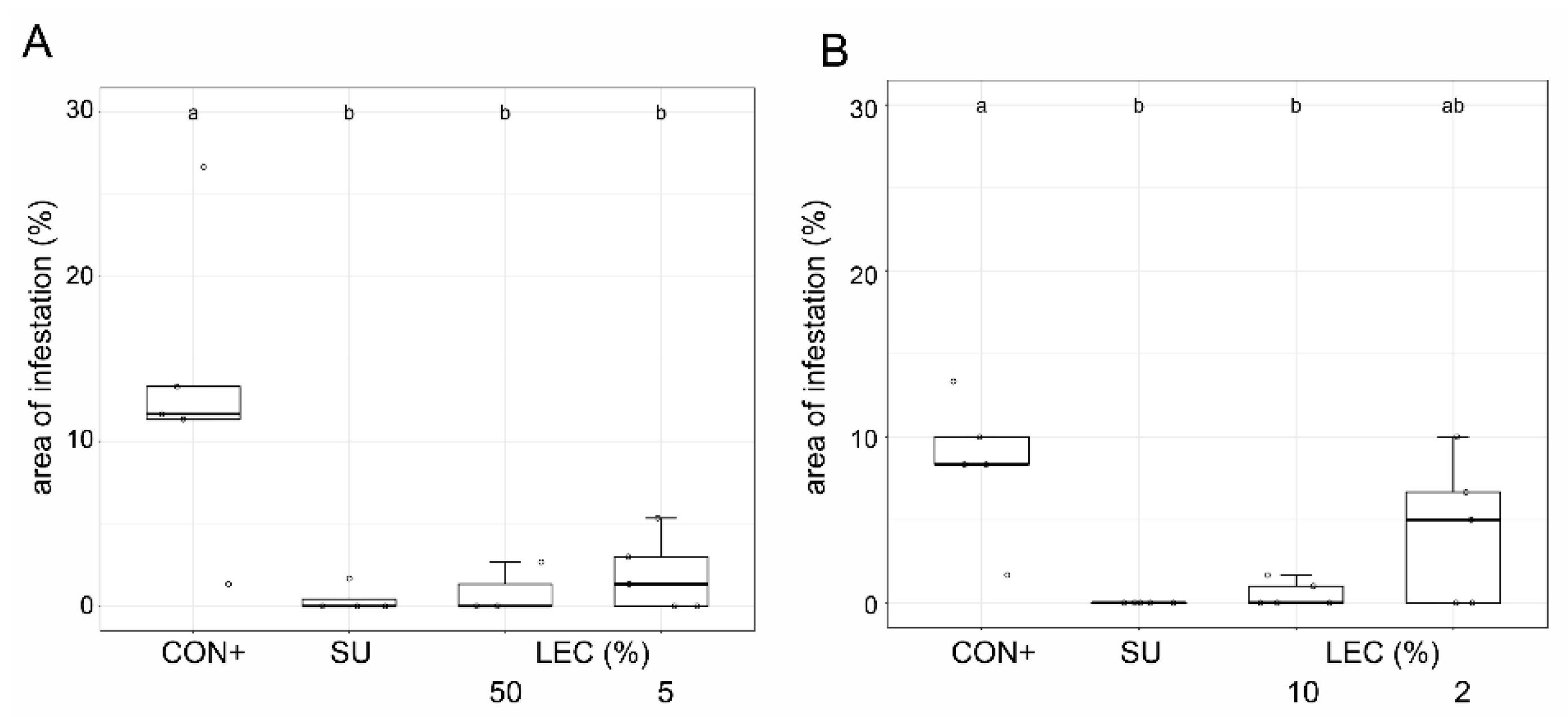
2.3. Ad Planta Activity of Lysobacter enzymogenes Isolate LEC
3. Discussion
4. Materials and Methods
4.1. Cultivation of Bacterial Isolates
4.1.1. Lysobacter spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lysobacter Species | Isolate | Source |
|---|---|---|
| Lysobacter sp. | BI-6067 | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6432/1 Kg | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6432/2 Kc (LEC) | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6434 | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6445 | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6447 | JKI-BI, cabbage roots, greenhouse |
| L. enzymogenes | BI-6457 | Cabbage roots, field, Darmstadt |
| Lysobacter sp. | Nr. 31, Wolf | Georg-August-University of Göttingen, Plant Pathology and Crop Protection Section |
4.1.2. Phytopathogenic Bacteria
4.2. Cultivation of Fungal and Oomycetal Isolates
4.3. Cultivation of Plants
4.4. Screening of Different Lysobacter sp. Isolates
4.4.1. Genotyping of Isolates
4.4.2. Analysis of Enzyme Activity
4.4.3. Dual Cultures of Lysobacter spp. with Fungal and Oomycetal Pathogens
4.4.4. Dual Cultures of Lysobacter sp. with Bacterial Pathogens
4.5. In Vitro Activity of Lysobacter enzymogenes Isolate LEC
4.5.1. Zoospore Release of Pseudoperonospora cubensis
4.5.2. Sporangia Germination Test with Phytophthora infestans
4.5.3. Spore Germination Test of Venturia inaequalis
4.5.4. Botrytis cinerea on Strawberry Leaves
4.5.5. Plasmopara viticola on Grapevine Leaf Discs
4.6. Ad Planta Activity of Lysobacter enzymogenes Isolate LEC
4.6.1. Pseudoperonospora cubensis on Cucumber Plants
4.6.2. Phytophthora infestans on Tomato Plants
4.6.3. Alternaria solani on Tomato Plants
4.6.4. Venturia inaequalis on Apple Plants
4.7. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfray, C.; Beddington, J.; Crute, I.; Haddad, L.; Lawrence, D.; Muir, J.; Pretty, J.; Robinson, S.; Thomas, S.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Köhler, H.-R.; Triebskorn, R. Wildlife ecotoxicology of pesticides: Can we track effects to the population level and beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Alavanja, M.C.R.; Hoppin, J.A.; Kamel, F. Health effects of chronic pesticide exposure: Cancer and neurotoxicity. Annu. Rev. Public Health 2004, 25, 155–197. [Google Scholar] [CrossRef] [PubMed]
- Massi, F.; Torriani, S.F.F.; Borghi, L.; Toffolatti, S.L. Fungicide resistance evolution and detection in plant pathogens: Plasmopara viticola as a case study. Microorganisms 2021, 9, 119. [Google Scholar] [CrossRef]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Schwab, S.; Rupp, C.; Bieler, E.; Dürrenberger, M.; Bleyer, G.; Schumacher, S.; Kassemeyer, H.-H.; Fuchs, R.; Schlücker, E. Microencapsulation—An innovative technique to improve the fungicide efficacy of copper against grapevine downy mildew. Crop Prot. 2021, 139, 105382. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Ortiz, A.; Sansinenea, E. Recent advancements for microorganisms and their natural compounds useful in agriculture. Appl. Microbiol. Biotechnol. 2021, 105, 891–897. [Google Scholar] [CrossRef]
- Feldmann, F.; Jehle, J.; Bradáčová, K.; Weinmann, M. Biostimulants, soil improvers, bioprotectants: Promoters of bio-intensification in plant production. J. Plant Dis. Prot. 2022, 129, 707–713. [Google Scholar] [CrossRef]
- Andrić, S.; Meyer, T.; Ongena, M. Bacillus responses to plant-associated fungal and bacterial communities. Front. Microbiol. 2020, 11, 1350. [Google Scholar] [CrossRef]
- Anderson, A.J.; Kim, Y.C. Insights into plant-beneficial traits of probiotic Pseudomonas chlororaphis isolates. J. Med. Microbiol. 2020, 69, 361–371. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Parte, A.C.; Sarda Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Goker, M. List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.M.; Yang, H.J.; Huang, J.G.; Yuan, L. Lysobacter enzymogenes LE16 autolysates have potential as biocontrol agents-Lysobacter sp. autolysates as biofungicide. J. Appl. Microbiol. 2020, 129, 1684–1692. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic micro-organisms. J. Appl. Microbiol. 2018, 124, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, H. The Genus Lysobacter. In The Prokaryotes: Volume 6: Proteobacteria: Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 939–957. [Google Scholar]
- Lin, L.; Xu, K.; Shen, D.; Chou, S.H.; Gomelsky, M.; Qian, G. Antifungal weapons of Lysobacter, a mighty biocontrol agent. Environ. Microbiol. 2021, 23, 5704–5715. [Google Scholar] [CrossRef]
- Hayward, A.C.; Fegan, N.; Fegan, M.; Stirling, G.R. Stenotrophomonas and Lysobacter: Ubiquitous plant-associated gamma-proteobacteria of developing significance in applied microbiology. J. Appl. Microbiol. 2010, 108, 756–770. [Google Scholar] [CrossRef]
- Nakayama, T.; Homma, Y.; Hashidoko, Y.; Mizutani, J.; Tahara, S. Possible role of xanthobaccins produced by Stenotrophomonas sp. strain SB-K88 in suppression of sugar beet damping-off disease. Appl. Environ. Microbiol. 1999, 65, 4334–4339. [Google Scholar] [CrossRef]
- Puopolo, G. Identification and characterization of Lysobacter capsici strain PG4: A new plant health-promoting rhizobacterium. J. Plant Pathol. 2010, 92, 157–164. Available online: https://www.jstor.org/stable/41998779 (accessed on 29 January 2023).
- Puopolo, G.; Giovannini, O.; Pertot, I. Lysobacter capsici AZ78 can be combined with copper to effectively control Plasmopara viticola on grapevine. Microbiol. Res. 2014, 169, 633–642. [Google Scholar] [CrossRef]
- Markellou, E.; Kapaxidi, E.; Karamaouna, F.; Samara, M.; Kyriakopoulou, K.; Anastasiadou, P.; Vavoulidou, E.; Meidanis, M.; Machera, K.; Mandoulaki, A.; et al. Evaluation of plant protection efficacy in field conditions and side effects of Lysobacter capsici AZ78, a biocontrol agent of Plasmopara viticola. Biocontrol Sci. Technol. 2022, 32, 930–951. [Google Scholar] [CrossRef]
- Ji, G.-H.; Wei, L.-F.; He, Y.-Q.; Wu, Y.-P.; Bai, X.-H. Biological control of rice bacterial blight by Lysobacter antibioticus strain 13-1. Biol. Control 2008, 45, 288–296. [Google Scholar] [CrossRef]
- Folman, L.B.; De Klein, M.J.E.M.; Postma, J.; van Veen, J.A. Production of antifungal compounds by Lysobacter enzymogenes isolate 3.1T8 under different conditions in relation to its efficacy as a biocontrol agent of Pythium aphanidermatum in cucumber. Biol. Control 2004, 31, 145–154. [Google Scholar] [CrossRef]
- Jochum, C.C.; Osborne, L.E.; Yuen, G.Y. Fusarium head blight biological control with Lysobacter enzymogenes strain C3. Biol. Control 2006, 39, 336–344. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 1–19. [Google Scholar] [CrossRef]
- Vasilyeva, N.V.; Tsfasman, I.M.; Suzina, N.E.; Stepnaya, O.A.; Kulaev, I.S. Secretion of bacteriolytic endopeptidaseL5 of Lysobacter sp. XL1 into the medium by means of outer membrane vesicles. FEBS J. 2008, 275, 3827–3835. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuen, G.Y. Effects of culture fluids and preinduction of chitinase production on biocontrol of Bipolaris leaf spot by Stenotrophomonas maltophilia C3. Biol. Control 2000, 18, 277–286. [Google Scholar] [CrossRef]
- Christensen, P.; Cook, F.D. Lysobacter, a new genus of nonfruiting, gliding bacteria with a high base ratio. Int. J. Syst. Evol. Microbiol. 1978, 28, 367–393. [Google Scholar] [CrossRef]
- Palumbo, J.D.; Yuen, G.Y.; Jochum, C.C.; Tatum, K.; Kobayashi, D.Y. Mutagenesis of β-1,3-glucanase genes in Lysobacter enzymogenes strain C3 results in reduced biological control activity toward Bipolaris leaf spot of tall fescue and Pythium damping-off of sugar beet. Phytopathology 2005, 95, 701–707. [Google Scholar] [CrossRef]
- Brescia, F.; Vlassi, A.; Bejarano, A.; Seidl, B.; Marchetti-Deschmann, M.; Schuhmacher, R.; Puopolo, G. Characterisation of the antibiotic profile of Lysobacter capsici AZ78, an effective biological control agent of plant pathogenic microorganisms. Microorganisms 2021, 9, 1320. [Google Scholar] [CrossRef]
- Hou, J.; Robbel, L.; Marahiel, M.A. Identification and characterization of the Lysobactin biosynthetic gene cluster reveals mechanistic insights into an unusual termination module architecture. Chem. Biol. 2011, 18, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Jochum, C.C.; Yu, F.; Zaleta-Rivera, K.; Du, L.; Harris, S.D.; Yuen, G.Y. An antibiotic complex from Lysobacter enzymogenes Strain C3: Antimicrobial activity and role in plant disease control. Phytopathology 2008, 98, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zaleta-Rivera, K.; Zhu, X.; Huffman, J.; Millet, J.C.; Harris, S.D.; Yuen, G.; Li, X.-C.; Du, L. Structure and biosynthesis of heat-stable antifungal factor (HSAF), a broad-spectrum antimycotic with a novel mode of action. Antimicrob. Agents Chemother. 2007, 51, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cheng, C.; Jiang, T.; Xu, H.; Chen, Y.U.N.; Ma, Z.; Qian, G.; Liu, F. Control of wheat Fusarium head blight by heat-stable antifungal factor (HSAF) from Lysobacter enzymogenes. Plant Dis. 2019, 103, 1286–1292. [Google Scholar] [CrossRef]
- Folman, L.B.; Postma, J.; van Veen, J.A. Characterisation of Lysobacter enzymogenes (Christensen and Cook 1978) strain 3.1T8, a powerful antagonist of fungal diseases of cucumber. Microbiol. Res. 2003, 158, 107–115. [Google Scholar] [CrossRef]
- Islam, M.T.; Hashidoko, Y.; Deora, A.; Ito, T.; Tahara, S. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne peronosporomycetes. Appl. Environ. Microbiol. 2005, 71, 3786–3796. [Google Scholar] [CrossRef]
- Lazazzara, V.; Perazzolli, M.; Pertot, I.; Biasioli, F.; Puopolo, G.; Cappellin, L. Growth media affect the volatilome and antimicrobial activity against Phytophthora infestans in four Lysobacter type strains. Microbiol. Res. 2017, 201, 52–62. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuen, G.Y. Biological control of Bipolaris sorokiniana on tall fescue by Stenotrophomonas maltophilia strain C3. Phytopathology 1999, 89, 817–822. [Google Scholar] [CrossRef]
- Knudsen, I.M.B.; Hockenhull, J.; Jensen, D.F.; Gerhardson, B.; Hökeberg, M.; Tahvonen, R.; Teperi, E.; Sundheim, L.; Henriksen, B. Selection of biological control agents for controlling soil and seed-borne diseases in the field. Eur. J. Plant Pathol. 1997, 103, 775–784. [Google Scholar] [CrossRef]
- Pfeiffer, T.; von Galen, A.; Zink, P.; Hübner, S.; Linkies, A.; Felgentreu, D.; Drechsel, J.; Birr, T.; Röder, O.; Kotte, M.; et al. Selection of bacteria and fungi for control of soilborne seedling diseases of maize. J. Plant Dis. Prot. 2021, 128, 1227–1241. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jung, H.; Kim, K.Y.; Park, S.K. An effective biocontrol bioformulation against Phytophthora blight of pepper using growth mixtures of combined chitinolytic bacteria under different field conditions. Eur. J. Plant Pathol. 2008, 120, 373–382. [Google Scholar] [CrossRef]
- Koch, E. Screening of rhizobacteria for antagonistic activity against Pythium ultimum on cucumber and kale. J. Plant Dis. Prot. 1997, 104, 353–361. Available online: https://www.jstor.org/stable/43215169 (accessed on 29 January 2023).
- Puopolo, G.; Cimmino, A.; Palmieri, M.C.; Giovannini, O.; Evidente, A.; Pertot, I. Lysobacter capsici AZ78 produces cyclo(l-Pro-l-Tyr), a 2,5-diketopiperazine with toxic activity against sporangia of Phytophthora infestans and Plasmopara viticola. J. Appl. Microbiol. 2014, 117, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Koch, E.; Kempf, H.J.; Hessenmüller, A. Characterization of the biocontrol activity and evaluation of potential plant growth-promoting properties of selected rhizobacteria. J. Plant Dis. Prot. 1998, 105, 567–580. Available online: https://www.jstor.org/stable/43386979 (accessed on 29 January 2023).
- Dreyer, F.; Koch, E. Characterization of bacterial populations from the rhizoplane of cabbage seedlings grown in different substrates. IOBC/WPRS Bull. 1996, 19, 36–41. [Google Scholar]
- Tang, B.; Zhao, Y.C.; Shi, X.M.; Xu, H.Y.; Zhao, Y.Y.; Dai, C.C.; Liu, F.Q. Enhanced heat stable antifungal factor production by Lysobacter enzymogenes OH11 with cheap feedstocks: Medium optimization and quantitative determination. Lett. Appl. Microbiol. 2018, 66, 439–446. [Google Scholar] [CrossRef]
- Miller, P.M. V-8 juice agar as a general-purpose medium for fungi and bacteria. Phytopathology 1955, 45, 461–462. [Google Scholar]
- Caten, C.E.; Jinks, J.L. Spontaneous variability of single isolates of Phytophthora infestans. I. Cultural variation. Can. J. Bot. 1968, 46, 329–348. [Google Scholar] [CrossRef]
- Kunz, S.; Mögel, G.; Hinze, M.; Volk, F. Control of apple scab by curative applications of biocontrol agents. In Proceedings of the Ecofruit-13th International Conference on Cultivation Technique and Phytopathological Problems in Organic Fruit-Growing, Weinsberg, Germany, 18–20 February 2008; Fördergemeinschaft Ökologischer Obstbau e.V.: Weinsberg, Germany, 2008; pp. 62–67. [Google Scholar]
- Schuster, C.; Konstantinidou-Doltsinis, S.; Schmitt, A. Glycyrrhiza glabra extract protects plants against important phytopathogenic fungi. Commun. Agric. Appl. Biol. Sci. 2010, 75, 531–540. [Google Scholar]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana activates expression of defence genes in grapevine and prevents infections by grapevine downy mildew Plasmopara viticola. Plant Pathol. 2019, 68, 1719–1731. [Google Scholar] [CrossRef]
- Hamp, T.J.; Jones, W.J.; Fodor, A.A. Effects of experimental choices and analysis noise on surveys of the “Rare Biosphere”. Appl. Environ. Microbiol. 2009, 75, 3263–3270. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Cowan, I.; Steel, S.T. Cowan and Steel’s Manual for the Identification of Medical Bacteria, 3 ed.; Barrow, G.I., Feltham, R.K.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; ISBN 9780521543286. [Google Scholar] [CrossRef]
- Atlas, R.M. Handbook of Microbiological Media; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9780429130496. [Google Scholar] [CrossRef]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Skujins, J.J.; Potgieter, H.J.; Alexander, M. Dissolution of fungal cell walls by a streptomycete chitinase and β-(1→3) glucanase. Arch. Biochem. Biophys. 1965, 111, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.P.; Burbano-Rosero, E.M.; Almeida, B.C.; Martins, G.G.; Albertini, L.S.; Rivera, I.N.G. Culture medium for isolating chitinolytic bacteria from seawater and plankton. World J. Microbiol. Biotechnol. 2009, 25, 2079–2082. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Ogunsanya, A.; Odeniyi, O.A. Optimization and characterization of tyrosinases from multi-enzyme producing Fusarium solani and Fumago sp. Period. Polytech. Chem. Eng. 2019, 63, 582–590. [Google Scholar] [CrossRef]
- Marth, E.H. Standard Methods for the Examination of Dairy Products, 14th ed.; FAO of the UN: Rome, Italy, 1978. [Google Scholar]
- Shahin, E.A.; Shepard, J.F. An Efficient Technique for Inducing Profuse Sporulation of Alternaria Species. Phytopathology 1979, 69, 618–620. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, v4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 3 December 2020).
- Pinheiro, J.; Bates, D.; Deb, R.S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed Effects Models_. R Package Version 3.1-149, 3.1-149; 2020. Available online: https://CRAN.R-project.org/package=nlme (accessed on 5 October 2020).
- Aitken, A.C. IV.—On least squares and linear combination of observations. Proc. R. Soc. Edinb. 1936, 55, 42–48. [Google Scholar] [CrossRef]
- Hansen, C.B. Generalized least squares inference in panel and multilevel models with serial correlation and fixed effects. J. Econom. 2007, 140, 670–694. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Unconstrained parametrizations for variance-covariance matrices. Stat. Comput. 1996, 6, 289–296. [Google Scholar] [CrossRef]
- Western, D.; Mose, V.N.; Worden, J.; Maitumo, D. Predicting extreme droughts in Savannah Africa: A comparison of proxy and direct measures in detecting biomass fluctuations, trends and their cause. PLoS ONE 2015, 10, e0136516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitarrini, G.; Soini, E.; Riccadonna, S.; Franceschi, P.; Zulini, L.; Masuero, D.; Vecchione, A.; Stefanini, M.; Di Gaspero, G.; Mattivi, F.; et al. Identification of biomarkers for defense response to Plasmopara viticola in a resistant grape variety. Front. Plant Sci. 2017, 8, 1524. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.4; 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 25 February 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, v.3.3.5; Springer: New York, NY, USA, 2016; Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 17 November 2021).








| Fungal/Oomycetal Pathogen | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ascochyta fabae | Alternaria radicina | Alternaria solani | Botrytis cinerea | Bipolaris sorokiniana | Fusarium culmorum | Phoma lingam | Rhizoctonia solani | Phytophthora infestans | Pythium ultimum | ||
| Lysobacter Species | Isolate | Diameter of Inhibition Zone (mm) | |||||||||
| Lysobacter sp. | BI-6067 | 28 ± 3 | 0 ± 0 | 21 ± 1 | 17 ± 2 | 16 ± 2 | 24 ± 0 | 27 ± 1 | 20 ± 0 | 31 ± 5 | 10 ± 0 |
| L. enzymogenes | BI-6432/1 Kg | 11 ± 0 | 0 ± 0 | 13 ± 2 | 17 ± 5 | 11 ± 1 | 10 ± 0 | 20 ± 4 | 15 ± 0 | 36 ± 3 | 10 ± 1 |
| L. enzymogenes (LEC) | BI-6432/2 Kc | 43 ± 1 | 0 ± 0 | 31 ± 7 | 42 ± 1 | 16 ± 1 | 32 ± 2 | 40 ± 3 | 37 ± 2 | 47 ± 5 | 12 ± 1 |
| L. enzymogenes | BI-6434 | 39 ± 5 | 0 ± 0 | 30 ± 10 | 45 ± 2 | 32 ± 2 | 30 ± 7 | 31 ± 1 | 22 ± 3 | 37 ± 1 | 10 ± 2 |
| L. enzymogenes | BI-6445 | 29 ± 9 | 0 ± 0 | 0 ± 0 | 28 ± 2 | 0 ± 0 | 6 ± 8 | 0 ± 0 | 11 ± 15 | 29 ± 2 | 0 ± 0 |
| L. enzymogenes | BI-6447 | 35 ± 6 | 9 ± 13 | 40 ± 7 | 37 ± 6 | 14 ± 0 | 47 ± 7 | 43 ± 2 | 20 ± 4 | 34 ± 2 | 10 ± 0 |
| L. enzymogenes | BI-6457 | 42 ± 4 | 14 ± 2 | 35 ± 6 | 37 ± 8 | 22 ± 6 | 34 ± 1 | 47 ± 2 | 21 ± 1 | 46 ± 4 | 12 ± 0 |
| Lysobacter sp. 1 | Nr. 31, Wolf | 32 ± 0 | 0 ± 0 | 0 ± 0 | 36 ± 3 | 20 ± 4 | 24 ± 2 | 30 ± 7 | 0 ± 0 | 30 ± 1 | 0 ± 0 |
| Bacterial Pathogen | ||||||
|---|---|---|---|---|---|---|
| Clavibacter michiganensis | Erwinia amylovora | Pseudomonas syringae | Pseudomonas tabaci | Xanthomonas campestris | ||
| Lysobacter Species | Isolate | Inhibition (+: Yes; −: No) | ||||
| Lysobacter sp. | BI-6067 | + | + | − | − | + |
| L. enzymogenes | BI-6432/1 Kg | + | + | − | − | + |
| L. enzymogenes (LEC) | BI-6432/2 Kc | + | + | − | − | + |
| L. enzymogenes | BI-6434 | + | + | − | − | + |
| L. enzymogenes | BI-6445 | + | + | − | − | + |
| L.enzymogenes | BI-6447 | + | + | − | − | + |
| L.enzymogenes | BI-6457 | + | + | − | − | + |
| Lysobacter sp. | Nr. 31, Wolf | + | − | − | − | + |
| Species | Isolate | Source |
|---|---|---|
| Clavibacter michiganensis ssp. sepedonicus | GSPB2825 | GSPB |
| Erwinia amylovora | E.a.639 | LTZ |
| Pseudomonas syringae pv. phaseolicola | GSPB1715 | GSPB |
| Pseudomonas tabaci | GSPB117 | GSPB |
| Xanthomonas campestris | GSPB1386 | GSPB |
| Fungal Species | Isolate | Source |
| Alternaria radicina | A.rad.1 | JKI-BI, carrot Seeds |
| Alternaria solani | A.s.714_1 | TU München, Dr. Hausladen, potato “Amado“, Hamersdorf |
| Alternaria solani | A.s.752_3 | TU München, Dr. Hausladen, potato “Kuras“, 29,468 Bergen |
| Ascochyta fabae | AF-4Re | JKI-BI/Prof. G. Kahl, University Frankfurt, Vicia faba, Iran |
| Botrytis cinerea | B.c.1 | JKI-BI, unknown |
| Botrytis cinerea | 222 | Bio- Protect GmbH, strawberry fruit, Konstanz |
| Bipolaris sorokiniana | BI-7191 | JKI-BI, Barley, Ukraine |
| Fusarium culmorum | VIII 18 | Kiel University, wheat |
| Phoma lingam | T12aD34 | University of Göttingen, oilseed rape |
| Rhizoctonia solani | AG2-2IIIb | Kiel University, maize |
| Venturia inaequalis | Bio- Protect GmbH, apple leaves, Konstanz | |
| Oomycetal Species | Isolate | Source |
| Pythium ultimum | P.u.1 | JKI-BI, cress |
| Phytophthora infestans | Syngenta K5509 | Syngenta, potato |
| Pseudoperonospora cubensis | JKI-BI, cucumber leaves | |
| Plasmopara viticola | isolate mixture | Geisenheim University, mix of five isolates from different origins (Geisenheim, Saulheim, Osann-Monzel, Wackernheim (Germany) and Remich (Luxemburg)) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drenker, C.; El Mazouar, D.; Bücker, G.; Weißhaupt, S.; Wienke, E.; Koch, E.; Kunz, S.; Reineke, A.; Rondot, Y.; Linkies, A. Characterization of a Disease-Suppressive Isolate of Lysobacter enzymogenes with Broad Antagonistic Activity against Bacterial, Oomycetal and Fungal Pathogens in Different Crops. Plants 2023, 12, 682. https://doi.org/10.3390/plants12030682
Drenker C, El Mazouar D, Bücker G, Weißhaupt S, Wienke E, Koch E, Kunz S, Reineke A, Rondot Y, Linkies A. Characterization of a Disease-Suppressive Isolate of Lysobacter enzymogenes with Broad Antagonistic Activity against Bacterial, Oomycetal and Fungal Pathogens in Different Crops. Plants. 2023; 12(3):682. https://doi.org/10.3390/plants12030682
Chicago/Turabian StyleDrenker, Christian, Doris El Mazouar, Gerrit Bücker, Sonja Weißhaupt, Eveline Wienke, Eckhard Koch, Stefan Kunz, Annette Reineke, Yvonne Rondot, and Ada Linkies. 2023. "Characterization of a Disease-Suppressive Isolate of Lysobacter enzymogenes with Broad Antagonistic Activity against Bacterial, Oomycetal and Fungal Pathogens in Different Crops" Plants 12, no. 3: 682. https://doi.org/10.3390/plants12030682