
Understanding the Active Mechanisms of Plant (Sesuvium portulacastrum L.) against Heavy Metal Toxicity
 , , ,
, , ,
Abstract
:1. Introduction
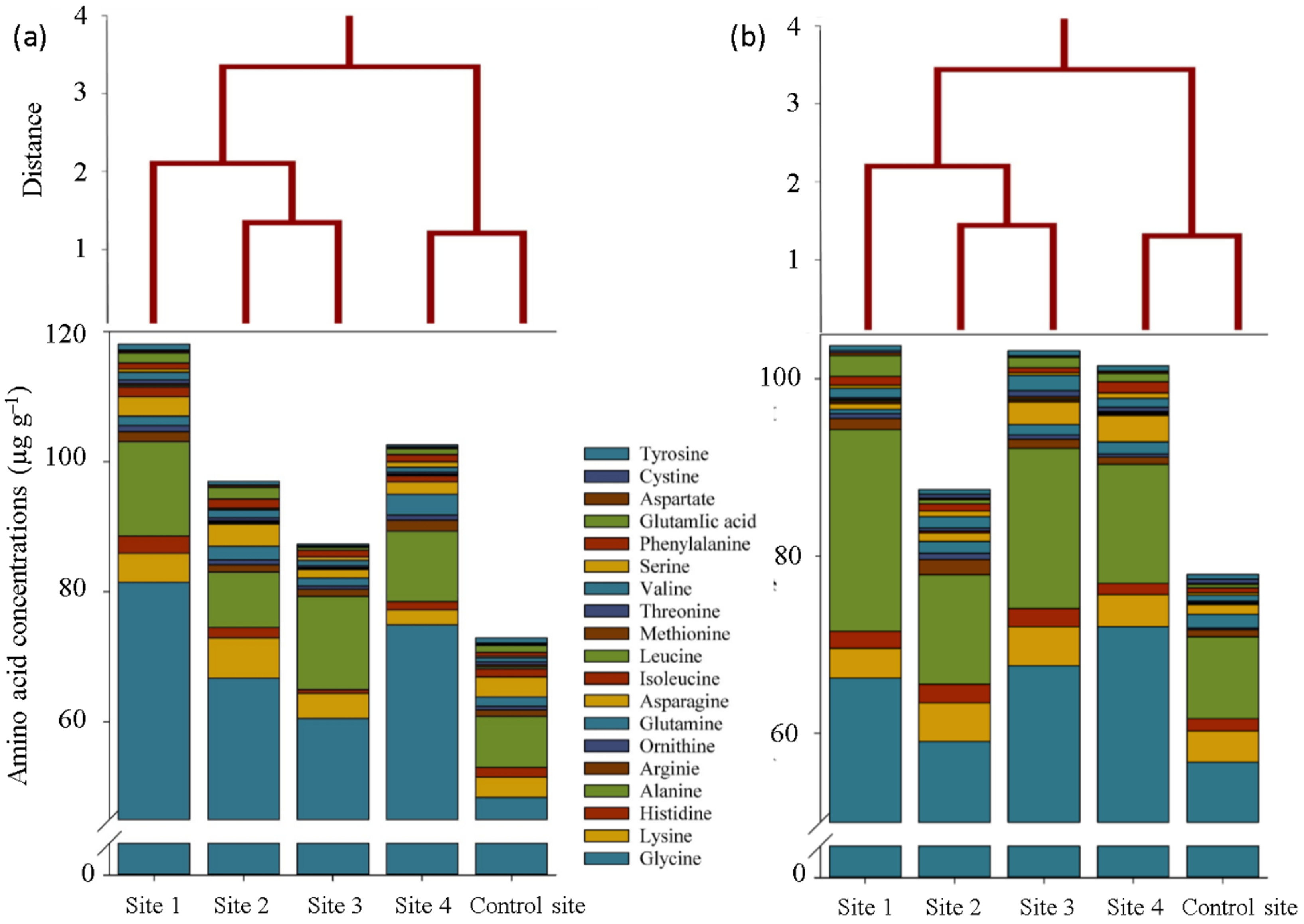
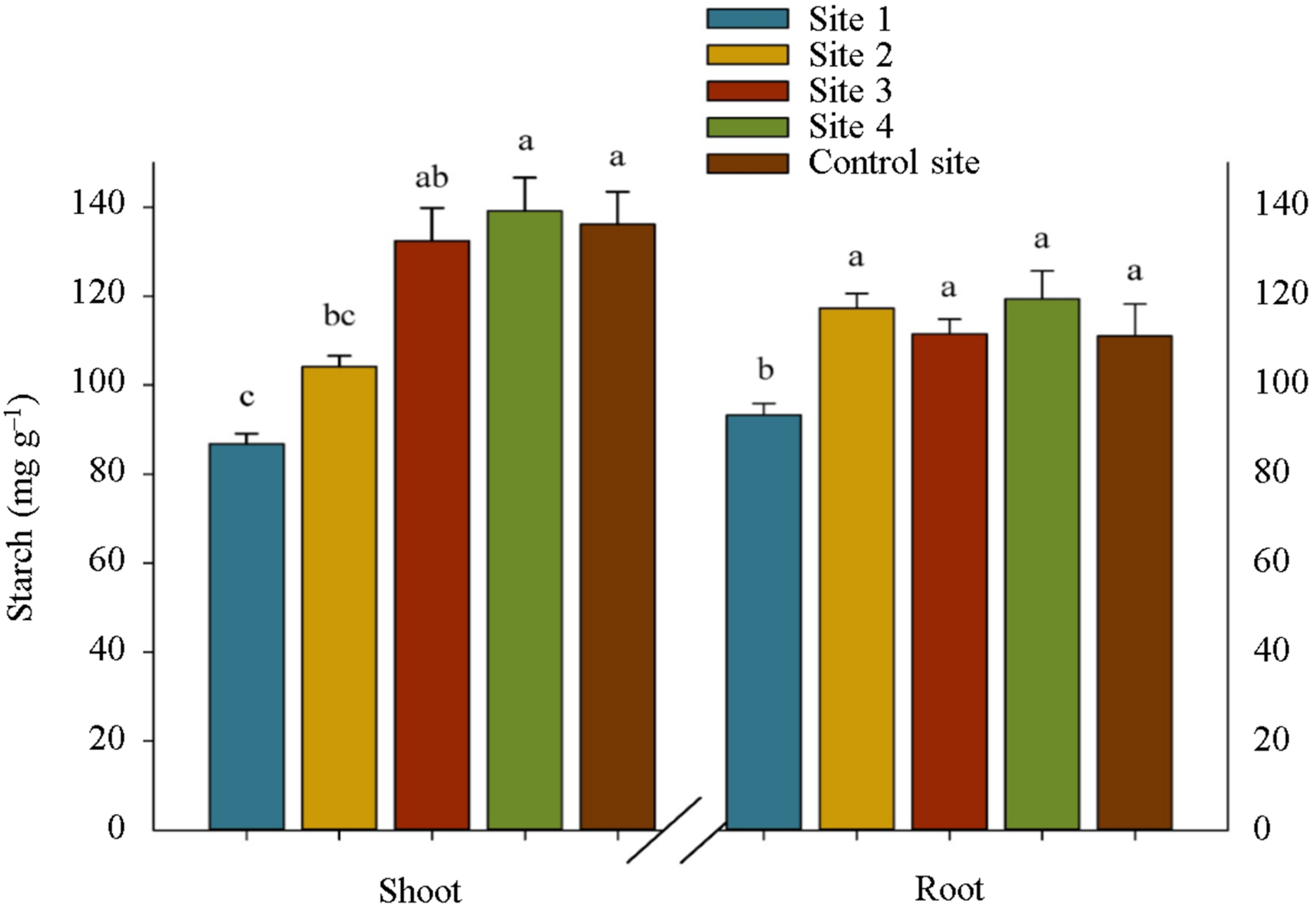
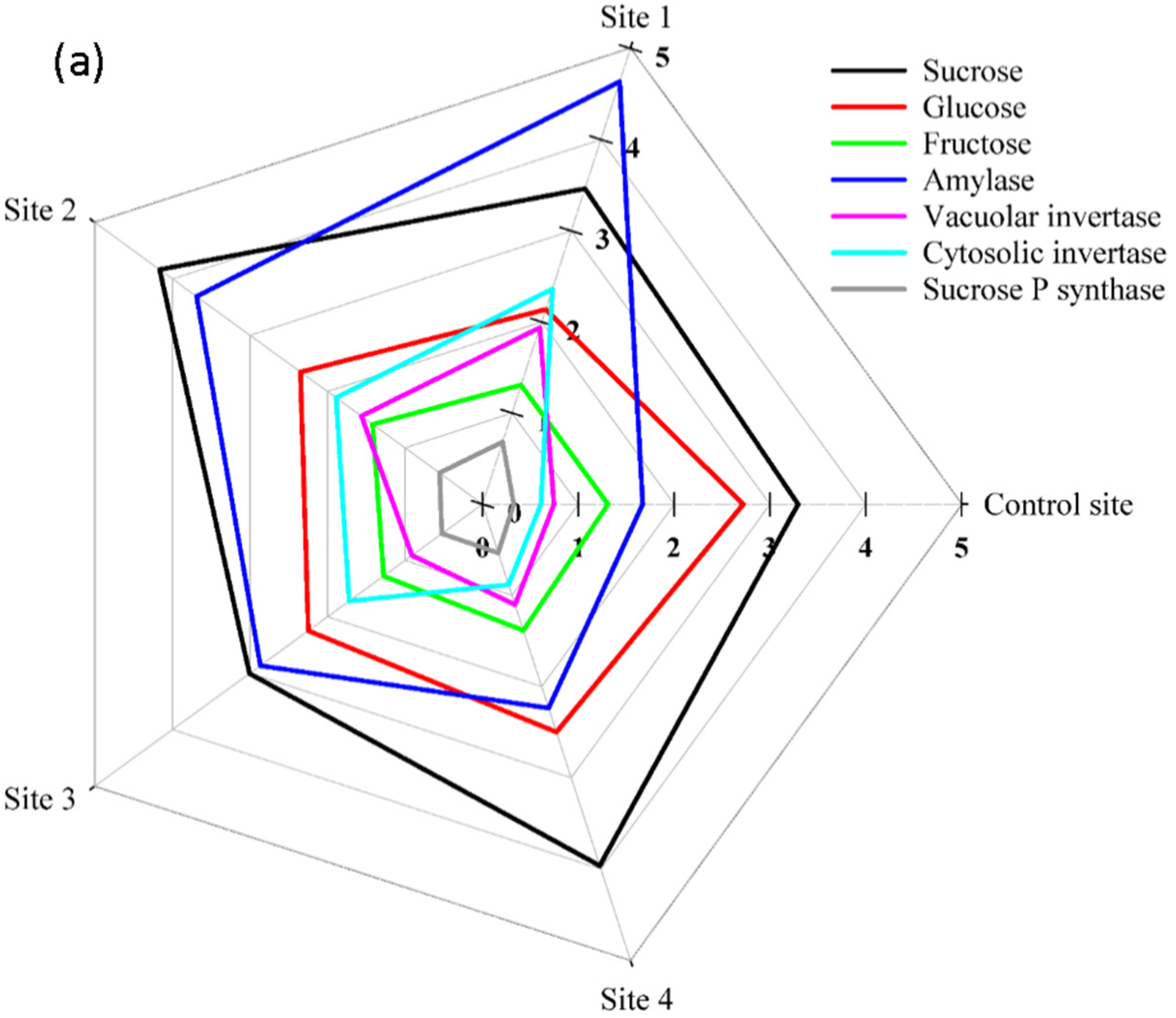
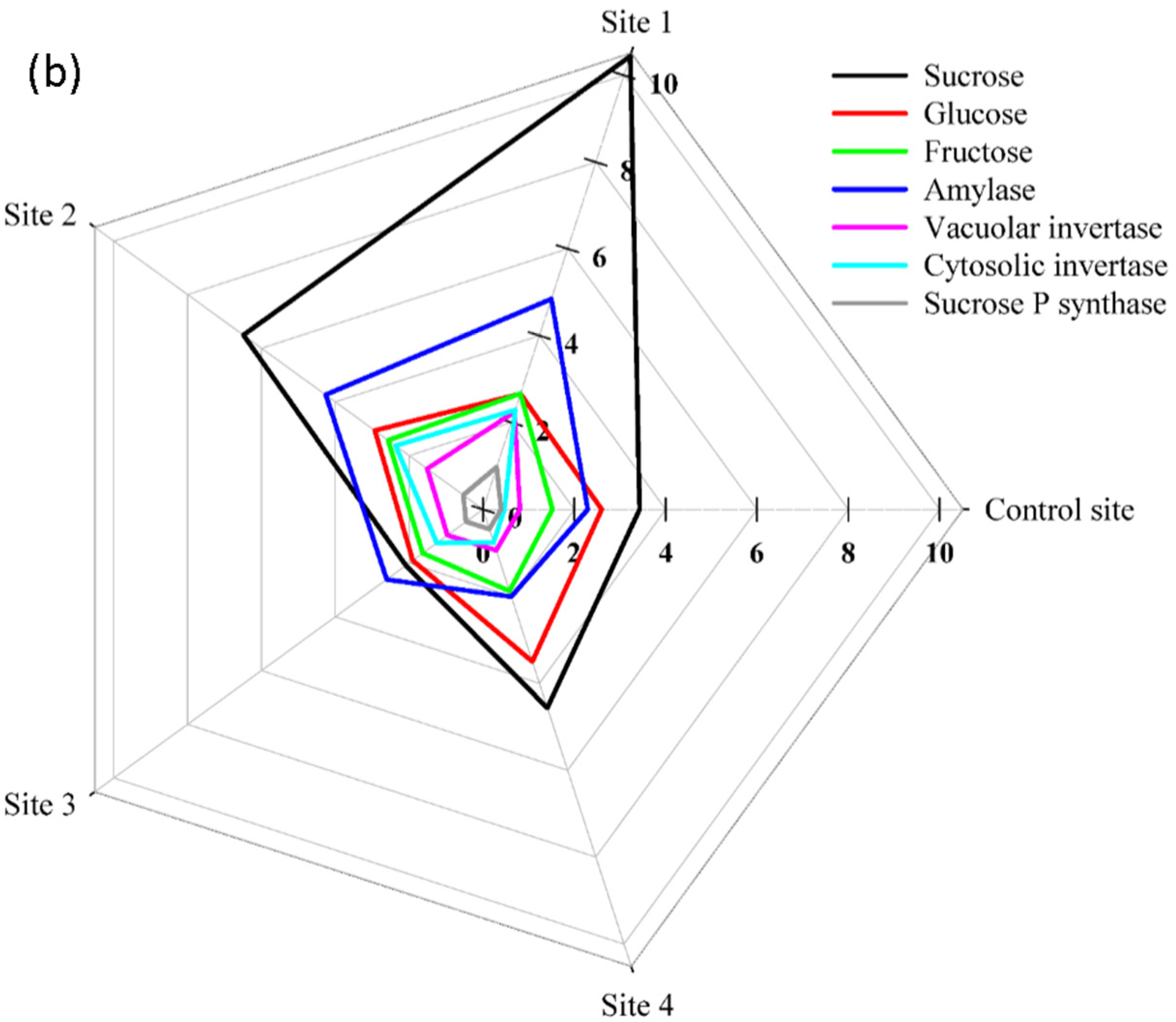
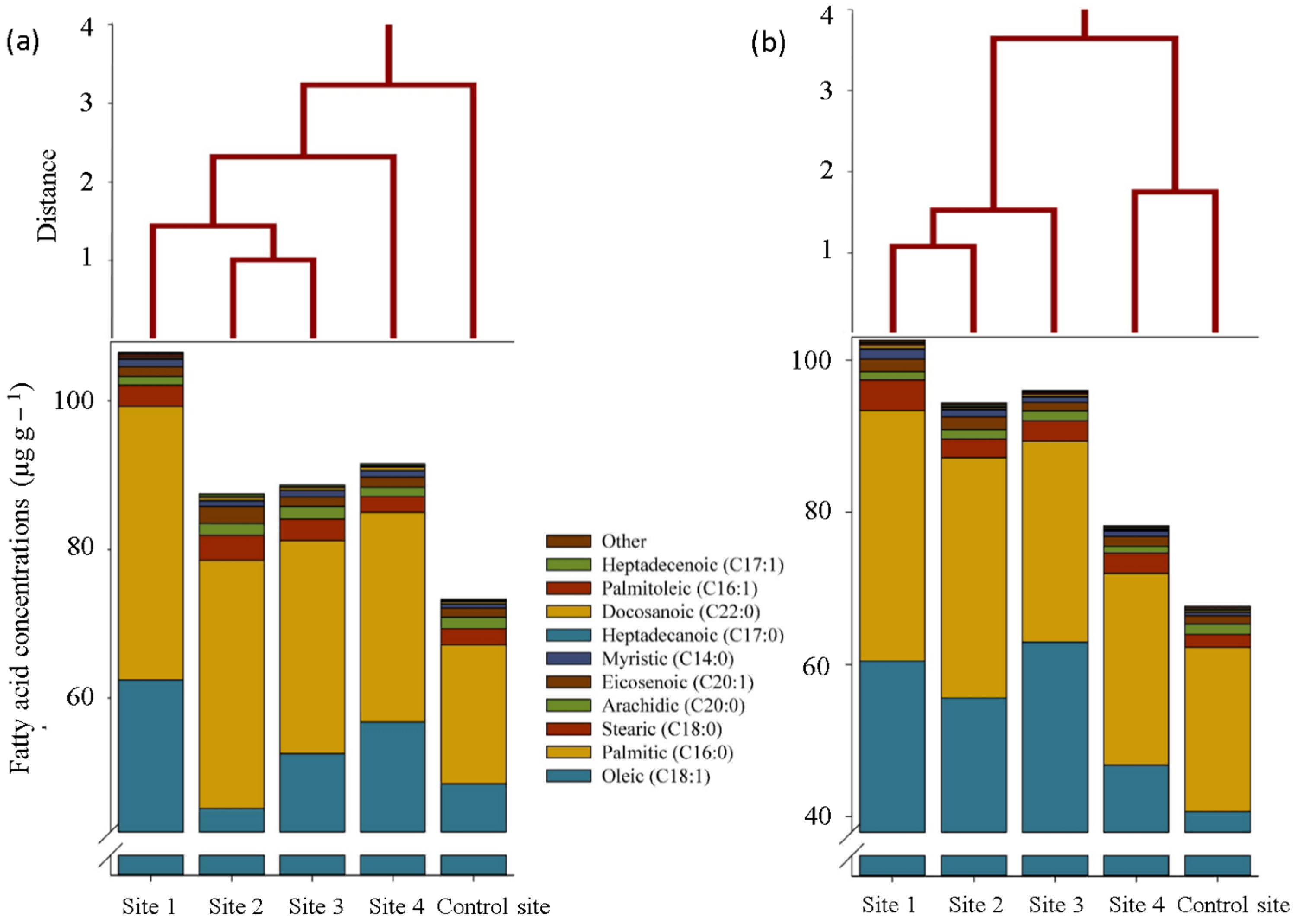
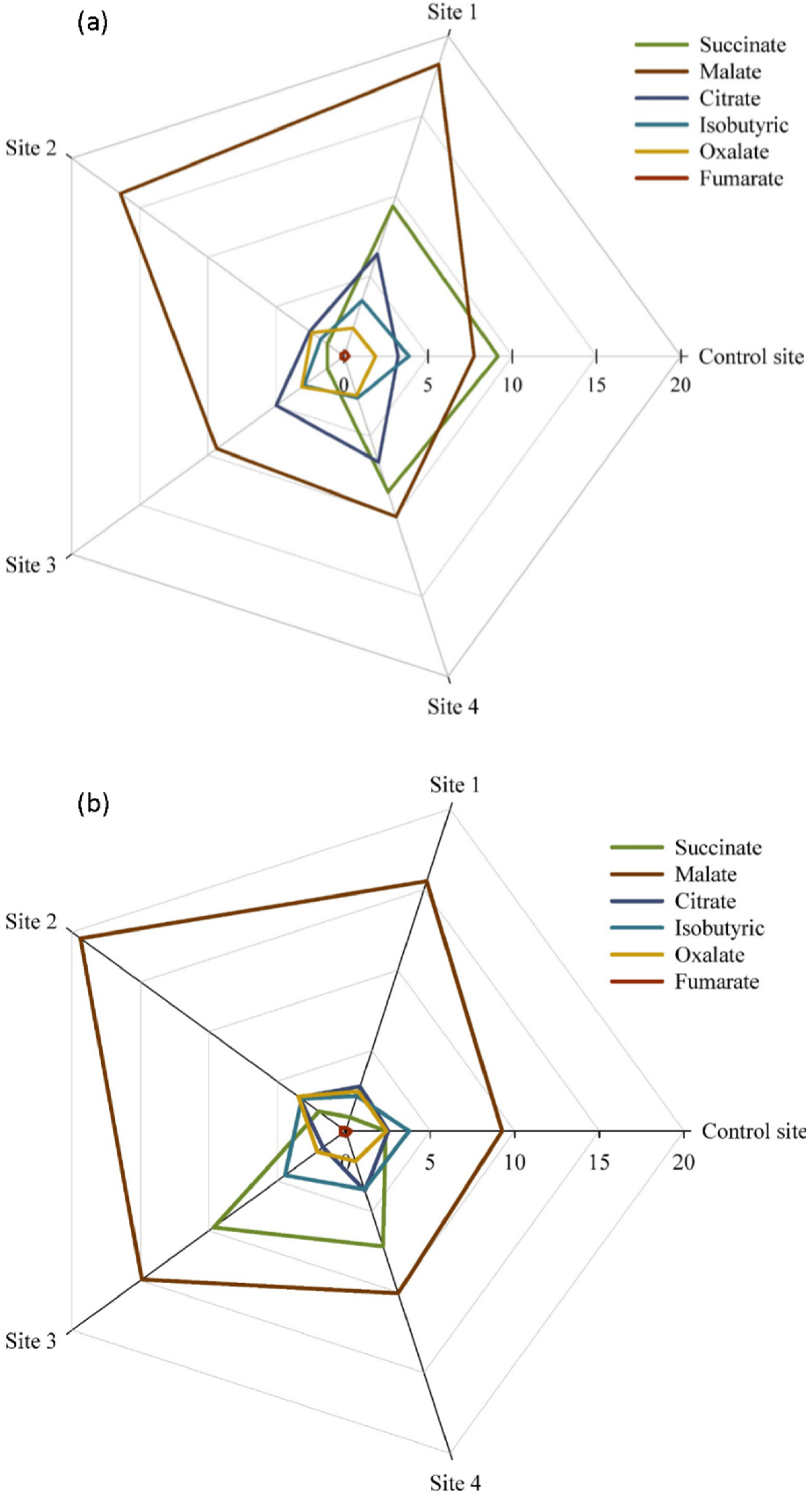
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Set up
4.2. Measurement of Amino Acid Profile
4.3. Carbohydrate Extraction and Estimation
4.4. Organic Acid Analysis
4.5. Assessment of Fatty Acid Profile
4.6. Determination of Tocopherol Content
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, S.U.; Nawaz, M.F.; Gul, S.; Yasin, G.; Hussain, B.; Li, Y.; Cheng, H. State-of-the-art OMICS strategies against toxic effects of heavy metals in plants: A review. Ecotoxicol. Environ. Saf. 2022, 242, 113952. [Google Scholar] [CrossRef] [PubMed]
- Terzano, R.; Rascio, I.; Allegretta, I.; Porfido, C.; Spagnuolo, M.; Khanghahi, M.Y.; Crecchio, C.; Sakellariadou, F.; Gattullo, C.E. Fire effects on the distribution and bioavailability of potentially toxic elements (PTEs) in agricultural soils. Chemosphere 2021, 281, 130752. [Google Scholar] [CrossRef] [PubMed]
- Rascio, I.; Curci, M.; Gattullo, C.; Lavecchia, A.; Khanghahi, M.Y.; Terzano, R.; Crecchio, C. Combined Effect of Laboratory-Simulated Fire and Chromium Pollution on Microbial Communities in an Agricultural Soil. Biology 2021, 10, 587. [Google Scholar] [CrossRef] [PubMed]
- Tytła, M.; Widziewicz, K.; Zielewicz, E. Heavy metals and its chemical speciation in sewage sludge at different stages of processing. Environ. Technol. 2016, 37, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Alsherif, E.A.; Al-Shaikh, T.M.; Almaghrabi, O.; AbdElgawad, H. High Redox Status as the Basis for Heavy Metal Tolerance of Sesuvium portulacastrum L. Inhabiting Contaminated Soil in Jeddah, Saudi Arabia. Antioxidants 2022, 11, 19. [Google Scholar] [CrossRef]
- Anjitha, K.; Sameena, P.; Puthur, J.T. Functional aspects of plant secondary metabolites in metal stress tolerance and their importance in pharmacology. Plant Stress 2021, 2, 100038. [Google Scholar] [CrossRef]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Saeed, D.A.; Rizwan, M.; Nawaz, M.A.; Mehmood, S.; et al. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manag. 2016, 183, 521–529. [Google Scholar] [CrossRef]
- Feng, Z.; Ji, S.; Ping, J.; Cui, D. Recent advances in metabolomics for studying heavy metal stress in plants. TrAC Trends Anal. Chem. 2021, 143, 116402. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef]
- Wang, S.; He, T.; Xu, F.; Li, X.; Yuan, L.; Wang, Q.; Liu, H. Analysis of physiological and metabolite response of Celosia argentea to copper stress. Plant Biol. 2020, 23, 391–399. [Google Scholar] [CrossRef]
- Clemens, S.; Palmgren, M.; Kraemer, U. A long way ahead: Understanding and engineering plant metal accumulation. Trends Plant Sci. 2002, 7, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Chen, W.; Lai, X.; Dai, H.; Sun, H.; Zhou, X.; Chen, T. Metabolic responses and their correlations with phytochelatins in Amaranthus hypochondriacus under cadmium stress. Environ. Pollut. 2019, 252, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, D.-Y.; Zhu, Q.-H.; Li, B.-Z.; Xu, C.; Zhu, H.-H.; Zhang, Q. Agronomic traits and ionomics influence on Cd accumulation in various sorghum (Sorghum bicolor (L.) Moench) genotypes. Ecotoxicol. Environ. Saf. 2021, 214, 112019. [Google Scholar] [CrossRef] [PubMed]
- Pidatala, V.R.; Li, K.; Sarkar, D.; Wusirika, R.; Datta, R. Comparative metabolic profiling of vetiver (Chrysopogon zizanioides) and maize (Zea mays) under lead stress. Chemosphere 2018, 193, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Wang, S.; Sun, L.-N.; Wang, H. Metabolic profiling of root exudates from two ecotypes of Sedum alfredii treated with Pb based on GC-MS. Sci. Rep. 2017, 7, 39878. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Cao, X.; Tan, C.; Deng, Y.; Cai, R.; Peng, X.; Bai, J. Analysis of the effect of cadmium stress on root exudates of Sedum plumbizincicola based on metabolomics. Ecotoxicol. Environ. Saf. 2020, 205, 111152. [Google Scholar] [CrossRef]
- Nigam, R.; Srivastava, S.; Prakash, S.; Srivastava, M.M. Cadmium mobilisation and plant availability–The impact of organic acids commonly exuded from roots. Plant Soil 2001, 230, 107–113. [Google Scholar] [CrossRef]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Dubey, S.; Misra, P.; Dwivedi, S.; Chatterjee, S.; Bag, S.K.; Mantri, S.; Asif, M.H.; Rai, A.; Kumar, S.; Shri, M.; et al. Transcriptomic and metabolomic shifts in rice roots in response to Cr (VI) stress. BMC Genom. 2010, 11, 648. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Mishra, B.; Chand, S.; Sangwan, N.S. ROS management is mediated by ascorbate-glutathione-α-tocopherol triad in co-ordination with secondary metabolic pathway under cadmium stress in Withania somnifera. Plant Physiol. Biochem. 2019, 139, 620–629. [Google Scholar] [CrossRef] [PubMed]
- De Agostini, A.; Cogoni, A.; Cortis, P.; Vacca, A.; Becerril, J.M.; Hernández, A.; Esteban, R. Heavy metal tolerance strategies in metallicolous and non-metallicolous populations of mosses: Insights of γ + β-tocopherol regulatory role. Environ. Exp. Bot. 2022, 194, 104738. [Google Scholar] [CrossRef]
- Sadiq, M.; Akram, N.A.; Ashraf, M.; Al-Qurainy, F.; Ahmad, P. Alpha-tocopherol-induced regulation of growth and metabolism in plants under non-stress and stress conditions. J. Plant Growth Regul. 2019, 38, 1325–1340. [Google Scholar] [CrossRef]
- Ali, E.; Hassan, Z.; Irfan, M.; Hussain, S.; Rehman, H.-U.; Shah, J.M.; Shahzad, A.N.; Ali, M.; Alkahtani, S.; Abdel-Daim, M.M.; et al. Indigenous Tocopherol Improves Tolerance of Oilseed Rape to Cadmium Stress. Front. Plant Sci. 2020, 11, 547133. [Google Scholar] [CrossRef] [PubMed]
- Hejna, M.; Moscatelli, A.; Stroppa, N.; Onelli, E.; Pilu, S.; Baldi, A.; Rossi, L. Bioaccumulation of heavy metals from wastewater through a Typha latifolia and Thelypteris palustris phytoremediation system. Chemosphere 2020, 241, 125018. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, T.; Niu, K.; Ma, H. Comparison and Characterization of Oxidation Resistance and Carbohydrate Content in Cd-Tolerant and -Sensitive Kentucky Bluegrass under Cd Stress. Agronomy 2021, 11, 2358. [Google Scholar] [CrossRef]
- Akinfemiwa, O.; Zubair, M.; Muniraj, T. Amylase. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK557738/ (accessed on 10 November 2022).
- Li, C.; Cao, Y.; Li, T.; Guo, M.; Ma, X.; Zhu, Y.; Fan, J. Changes in antioxidant system and sucrose metabolism in maize varieties exposed to Cd. Environ. Sci. Pollut. Res. 2022, 29, 64999–65011. [Google Scholar] [CrossRef]
- Alharthi, S.T.; El-Sheikh, M.A.; Alfarhan, A.A. Biological change of western Saudi Arabia: Alien plants diversity and their relationship with edaphic variables. J. King Saud Univ.-Sci. 2023, 35, 102496. [Google Scholar] [CrossRef]
- Ding, G.; Yang, Q.; Ruan, X.; Si, T.; Yuan, B.; Zheng, W.; Xie, Q.; Souleymane, O.A.; Wang, X. Proteomics analysis of the effects for different salt ions in leaves of true halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2022, 170, 234–248. [Google Scholar] [CrossRef]
- Zinta, G.; AbdElgawad, H.; Peshev, D.; Weedon, J.T.; van den Ende, W.; Nijs, I.; Janssens, I.A.; Beemster, G.T.S.; Asard, H. Dynamics of metabolic responses to periods of combined heat and drought in Arabidopsis thaliana under ambient and elevated atmospheric CO2. J. Exp. Bot. 2018, 69, 2159–2170. [Google Scholar] [CrossRef] [Green Version]
- Galtier, N.; Foyer, C.H.; Murchie, E.; Aired, R.; Quick, P.; Voelker, T.A.; Thépenier, C.; Lascève, G.; Betsche, T. Effects of light and atmospheric carbon dioxide enrichment on photosynthesis and carbon partitioning in the leaves of tomato (Lycopersicon esculentum L.) plants over-expressing sucrose phosphate synthase. J. Exp. Bot. 1995, 46, 1335–1344. [Google Scholar] [CrossRef]
- de Sousa, A.; AbdElgawad, H.; Asard, H.; Pinto, A.; Soares, C.; Branco-Neves, S.; Braga, T.; Azenha, M.; Selim, S.; Al Jaouni, S.; et al. Metalaxyl Effects on Antioxidant Defenses in Leaves and Roots of Solanum nigrum L. Front. Plant Sci. 2017, 8, 1967. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Peshev, D.; Zinta, G.; van den Ende, W.; Janssens, I.A.; Asard, H. Climate Extreme Effects on the Chemical Composition of Temperate Grassland Species under Ambient and Elevated CO2: A Comparison of Fructan and Non-Fructan Accumulators. PLoS ONE 2014, 9, e92044. [Google Scholar] [CrossRef] [PubMed]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC–MS combined with k-means cluster analysis. Ind. Crops Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- AbdElgawad, H.; De Vos, D.; Zinta, G.; Domagalska, M.A.; Beemster, G.T.S.; Asard, H. Grassland species differentially regulate proline concentrations under future climate conditions: An integrated biochemical and modelling approach. New Phytol. 2015, 208, 354–369. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Site | S4 | S3 | S2 | S1 | |
|---|---|---|---|---|---|
| Cd | 0.43 (µg g−1) | +239% | +608% | +1054% | +1107% |
| Ni | 0.441 (µg g−1) | −9% | +82% | +153% | +186% |
| As | 0.461 (µg g−1) | −27% | −50% | −24% | −2% |
| Cu | 0.863 (µg g−1) | +56% | +29% | +56% | +37% |
| Pb | 0.072 (µg g−1) | +1503% | +6475% | +10,189% | +14,761% |
| Co | 0.108 (µg g−1) | +595% | +3527% | +5240% | +7928% |
| Hg | 0.088 (µg g−1) | +309% | +3036% | +6483% | +6320% |
| Al | 0.046 (µg g−1) | +761% | +602% | +1267% | +1465% |
| V | 0.046 (µg g−1) | +585% | +922% | +1383% | +1683% |
| Cr | 0.06 (µg g−1) | +335% | +928% | +1125% | +1600% |
| Zn | 0.046 (µg g−1) | +109% | +1191% | +1587% | +1835% |
| Total bacteria | 232 | −11% | −18% | −21% | −37% |
| Spore forming bacteria | 47.2 | −5% | −8% | −15% | −27% |
| Actinobacteria | 33.5 | −17% | −23% | -37% | −35% |
| Total Fungi | 45.8 | −21% | −28% | −36% | −45% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsherif, E.A.; Yaghoubi Khanghahi, M.; Crecchio, C.; Korany, S.M.; Sobrinho, R.L.; AbdElgawad, H. Understanding the Active Mechanisms of Plant (Sesuvium portulacastrum L.) against Heavy Metal Toxicity. Plants 2023, 12, 676. https://doi.org/10.3390/plants12030676
Alsherif EA, Yaghoubi Khanghahi M, Crecchio C, Korany SM, Sobrinho RL, AbdElgawad H. Understanding the Active Mechanisms of Plant (Sesuvium portulacastrum L.) against Heavy Metal Toxicity. Plants. 2023; 12(3):676. https://doi.org/10.3390/plants12030676
Chicago/Turabian StyleAlsherif, Emad A., Mohammad Yaghoubi Khanghahi, Carmine Crecchio, Shereen Magdy Korany, Renato Lustosa Sobrinho, and Hamada AbdElgawad. 2023. "Understanding the Active Mechanisms of Plant (Sesuvium portulacastrum L.) against Heavy Metal Toxicity" Plants 12, no. 3: 676. https://doi.org/10.3390/plants12030676