
Phytoremediation of Potentially Toxic Elements from Contaminated Saline Soils Using Salvadora persica L.: Seasonal Evaluation
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
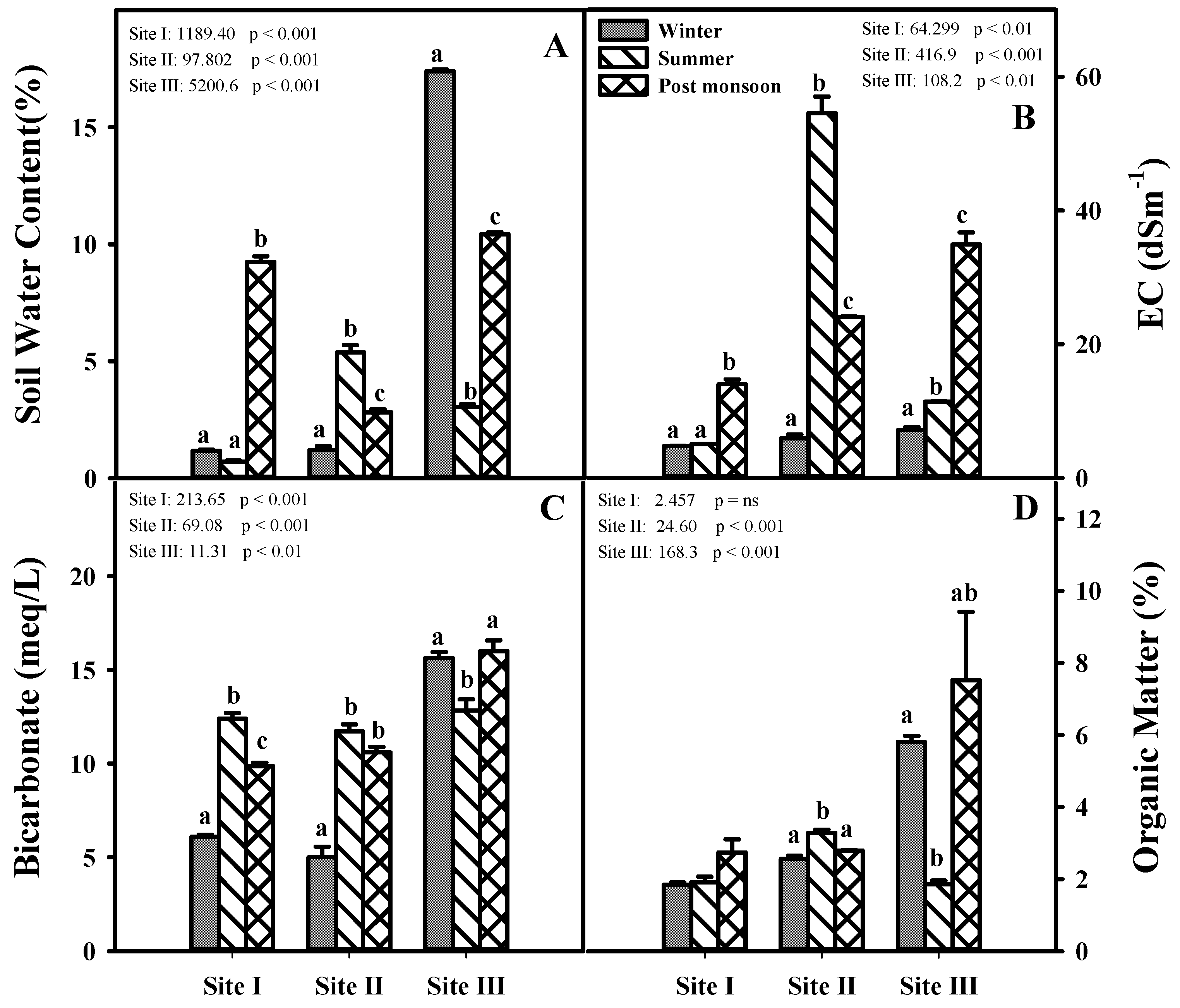
2.1. Seasonal Changes in Soil Characteristics at Different Study Sites
2.2. Seasonal and Spatial Variation in Soil Sediment Metal Ions
2.3. Seasonal Flux in Extractable Metal Ions in Plant Tissues
2.4. Seasonal Fluctuations in Leaf Water Relation Parameters at the Study Sites
3. Materials and Methods
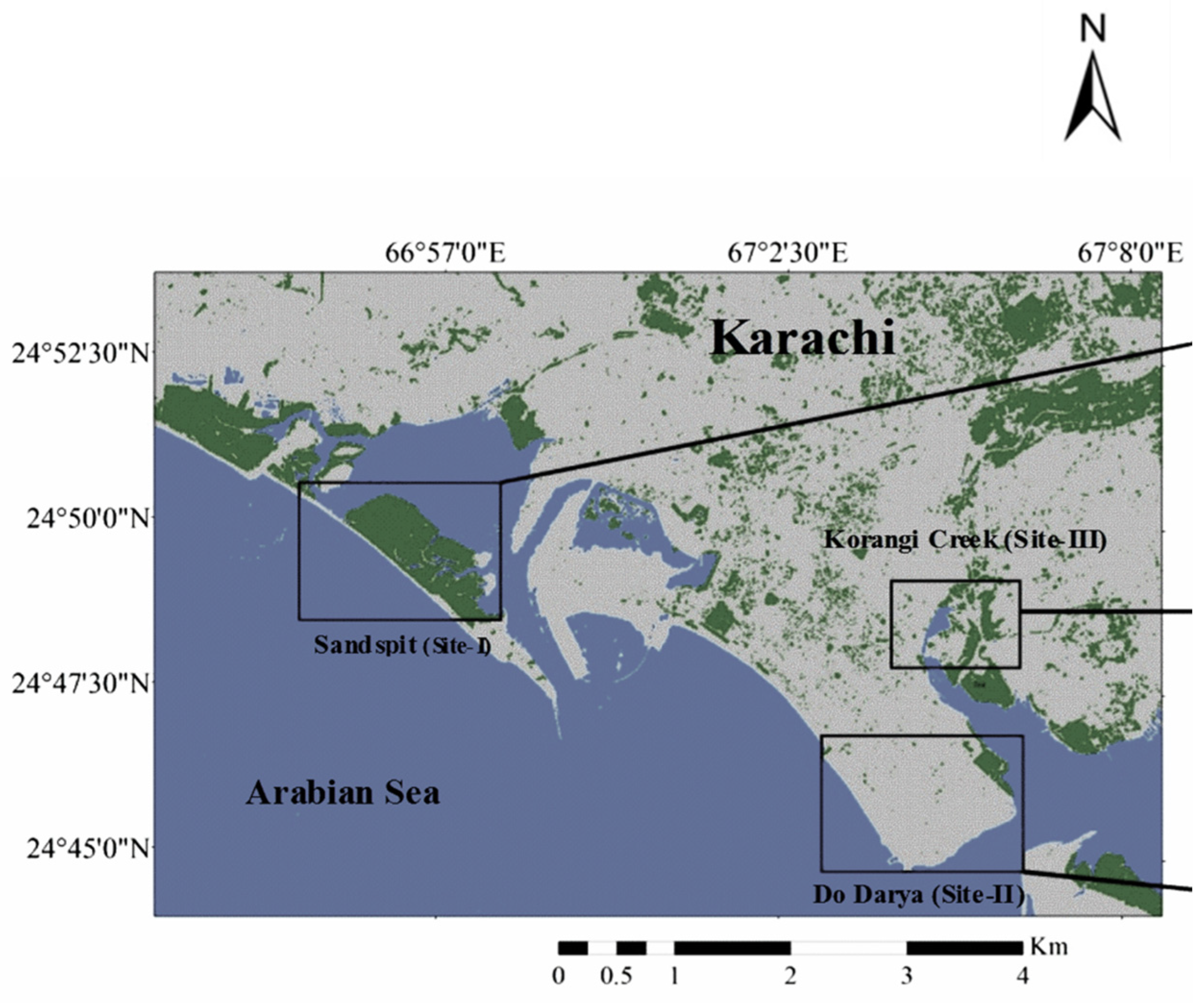
3.1. Description of Study Sites
3.2. Sample Collection
3.3. Plant Analyses
3.4. Soil Analyses
3.5. Heavy Metals (Fe, Mn, Pb, Cu, Zn, and Cr) and Ion (Na+ and K+) Analyses
3.6. Bio-Concentration Factor (BCF) and Translocation Factor (TF)
3.7. Water Relation (PV-Curve Derived) Parameters
3.8. Statistical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J.; Rafique, M.; Rizwan, M. Heavy metal remediation and resistance mechanism of Aeromonas, Bacillus, and Pseudomonas: A review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Ashfaq, A.; Khan, Z.I.; Ahmad, K.; Ashraf, M.A.; Hussain, M.I.; Elghareeb, E.M. Hazard of selenium metal contamination in vegetables grown in municipal solid waste amended soil: Assessment of the potential sources and systemic health effects. Agric. Water Manag. 2022, 271, 107768. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, F.; Zafar, A.; Khan, Z.I.; Ahmad, K.; Ch, S.A.; Batool, A.I.; Nadeem, M. Assessment of potential toxicological risk for public health of heavy metal iron in diverse wheat varieties irrigated with various types of waste water in South Asian country. Agric. Water Manag. 2023, 276, 108044. [Google Scholar] [CrossRef]
- Dahrazma, B.; Mulligan, C.N. Investigation of the removal of heavy metals from sediments using rhamnolipid in a continuous flow configuration. Chemosphere 2007, 69, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S. Plant Stress Physiology; Cabi: Cambridge, MA, USA, 2012. [Google Scholar]
- Singh, V.K.; Singh, R.; Rajput, V.D.; Singh, V.K. Halophytes for the sustainable remediation of heavy metal-contaminated sites: Recent developments and future perspectives. Chemosphere 2023, 313, 137524. [Google Scholar] [CrossRef]
- Jahromi, F.A.; Moore, F.; Keshavarzi, B.; Mohebbi-Nozar, S.L.; Mohammadi, Z.; Sorooshian, A.; Abbasi, S. Bisphenol A (BPA) and polycyclic aromatic hydrocarbons (PAHs) in the surface sediment and bivalves from Hormozgan Province coast-line in the Northern Persian Gulf: A focus on source apportionment. Mar. Pollut. Bull. 2020, 152, 110941. [Google Scholar] [CrossRef]
- Centofanti, T.; Banuelos, G. Evaluation of the halophyte Salsola soda as an alternative crop for saline soils high in selenium and boron. J. Environ. Manag. 2015, 157, 96–102. [Google Scholar] [CrossRef]
- Hossain, M.B.; Rakib, R.J.; Jolly, Y.; Rahman, M. Metals uptake and translocation in salt marsh macrophytes, Porteresia sp. from Bangladesh coastal area. Sci. Total Environ. 2020, 764, 144637. [Google Scholar] [CrossRef]
- Mujeeb, A.; Aziz, I.; Ahmed, M.Z.; Alvi, S.K.; Shafiq, S. Comparative assessment of heavy metal accumulation and bio-indication in coastal dune halophytes. Ecotoxicol. Environ. Saf. 2020, 195, 110486. [Google Scholar] [CrossRef]
- Bonanno, G.; Cirelli, G.L. Comparative analysis of element concentrations and translocation in three wetland congener plants: Typha domingensis, Typha latifolia and Typha angustifolia. Ecotoxicol. Environ. Saf. 2017, 143, 92–101. [Google Scholar] [CrossRef]
- Manousaki, E.; Kalogerakis, N. Phytoextraction of Pb and Cd by the Mediterranean saltbush (Atriplex halimus L.): Metal uptake in relation to salinity. Environ. Sci. Pollut. Res. 2009, 16, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Mariem, W.; Kilani, B.R.; Benet, G.; Abdelbasset, L.; Stanley, L.; Charlotte, P.; Chedly, A.; Tahar, G. How does NaCl improve tolerance to cadmium in the halophyte Sesuvium portulacastrum? Chemosphere 2014, 117, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Manousaki, E.; Galanaki, K.; Papadimitriou, L.; Kalogerakis, N. Metal Phytoremediation by the Halophyte Limoniastrum monopetalum (L.) Boiss: Two Contrasting Ecotypes. Int. J. Phytoremediat. 2014, 16, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Bankaji, I.; Caçador, I.; Sleimi, N. Physiological and biochemical responses of Suaeda fruticosa to cadmium and copper stresses: Growth, nutrient uptake, antioxidant enzymes, phytochelatin, and glutathione levels. Environ. Sci. Pollut. Res. 2015, 22, 13058–13069. [Google Scholar] [CrossRef] [PubMed]
- Vromman, D.; Flores-Bavestrello, A.; Šlejkovec, Z.; Lapaille, S.; Teixeira-Cardoso, C.; Briceño, M.; Kumar, M.; Martinez, J.P.; Lutts, S. Arsenic accumulation and distribution in relation to young seedling growth in Atriplex atacamensis Phil. Sci. Total Environ. 2011, 412–413, 286–295. [Google Scholar] [CrossRef]
- Panda, A.; Rangani, J.; Kumari, A.; Parida, A.K. Efficient regulation of arsenic translocation to shoot tissue and modulation of phytochelatin levels and antioxidative defense system confers salinity and arsenic tolerance in the Halophyte Suaeda maritima. Environ. Exp. Bot. 2017, 143, 149–171. [Google Scholar] [CrossRef]
- Sghaier, D.B.; Duarte, B.; Bankaji, I.; Caçador, I.; Sleimi, N. Growth, chlorophyll fluorescence and mineral nutrition in the halophyte Tamarix gallica cultivated in combined stress conditions: Arsenic and NaCl. J. Photochem. Photobiol. B Biol. 2015, 149, 204–214. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of native plants with phytoremediation potential for highly contaminated Mediterranean soil restoration: Tools for a non-destructive and integrative approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2012, 362, 319–334. [Google Scholar] [CrossRef]
- Caparrós, P.G.; Ozturk, M.; Gul, A.; Batool, T.S.; Pirasteh-Anosheh, H.; Unal, B.T.; Altay, V.; Toderich, K.N. Halophytes have potential as heavy metal phytoremediators: A comprehensive review. Environ. Exp. Bot. 2022, 193, 104666. [Google Scholar] [CrossRef]
- Caçador, I.; Vale, C.; Catarino, F. Seasonal variation of Zn, Pb, Cu and Cd concentrations in the root–sediment system of Spartina maritima and Halimione portulacoides from Tagus estuary salt marshes. Mar. Environ. Res. 2000, 49, 279–290. [Google Scholar] [CrossRef]
- Phulwaria, M.; Ram, K.; Gahlot, P.; Shekhawat, N.S. Micropropagation of Salvadora persica-a tree of arid horticulture and forestry. New For. 2011, 42, 317–327. [Google Scholar] [CrossRef]
- Reddy, M.P.; Shah, M.T.; Patolia, J.S. Salvadora persica, a potential species for industrial oil production in semiarid saline and alkali soils. Ind. Crop. Prod. 2008, 28, 273–278. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. BioMed Res. Int. 2014, 2014, 589341. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Qaiser, M. Halophytes of Pakistan: Characteristics, distribution and potential economic usages. In Sabkha Ecosystems, Tasks for Vegetation Science; Khan, M.A., Benno, B., Kust, G.S., Barth, H.J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 129–153. [Google Scholar] [CrossRef]
- Patel, H.M.; Patel, N.K. Sacred and medicinal plant diversity of Patan sacred grove of Patan district (N.G.). Life Sci. Leafl. 2017, 143, 50–60. [Google Scholar]
- Khatak, M.; Khatak, S.; Siddqui, A.; Vasudeva, N.; Aggarwal, A. Salvadora persica (Short Review). Pharmacogn. Rev. 2010, 4, 209–215. [Google Scholar] [CrossRef]
- Rao, G.G.; Nayak, A.K.; Chinchmalatpure, A.R.; Nath, A.; Babu, V.R. Growth and Yield of Salvadora persica, A Facultative Halophyte Grown on Saline Black Soil (Vertic Haplustept). Arid. Land Res. Manag. 2004, 18, 51–61. [Google Scholar] [CrossRef]
- Dagar, J.C.; Tomar, O.S.; Kumar, Y.; Bhagwan, H.; Yadav, R.K.; Tyagi, N.K. Performance of some under-explored crops under saline irrigation in a semiarid climate in Northwest India. Land Degrad. Dev. 2006, 17, 285–299. [Google Scholar] [CrossRef]
- Aljlil, A.A. Miswak (Salvadora persica roots): Discovery of a new biomaterial for removing heavy metals from water in Saudi Arabia. Mat. Technol. 2017, 51, 35–39. [Google Scholar] [CrossRef]
- Ileri, O.; Cay, S.; Uyanik, A.; Erduran, N. Removal of Common Heavy Metals from Aqueous Solutions by Waste Salvadora persica L. Branches (Miswak). Int. J. Environ. Res. 2014, 8, 987–996. [Google Scholar] [CrossRef]
- Patel, M.; Parida, A.K. Salinity alleviates the arsenic toxicity in the facultative halophyte Salvadora persica L. by the modulations of physiological, biochemical, and ROS scavenging attributes. J. Hazard. Mater. 2020, 401, 123368. [Google Scholar] [CrossRef] [PubMed]
- Chaudhri, I.I. The Vegetation of Karachi. Vegetatio 1961, 10, 229–246. [Google Scholar] [CrossRef]
- Siddique, A.; Mumtaz, M.; Zaigham, N.A.; Mallick, K.A.; Saied, S.; Zahir, E.; Khwaja, H.A. Heavy metal toxicity levels in the coastal sediments of the Arabian Sea along the urban Karachi (Pakistan) region. Mar. Pollut. Bull. 2009, 58, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Khan, M.U.; Khoso, S.A. Identification of biological threats zones along the coastline of Karachi. Int. J. Environ. Sci. Technol. 2018, 16, 3557–3564. [Google Scholar] [CrossRef]
- Alamgir, A.; Fatima, N.; Khan, M.A.; Rehman, M.; Shaukat, S.S. A preliminary pollution appraisal of western backwater at Karachi Coastal area. Appl. Water Sci. 2019, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Elahi, N.; Ahmed, Q.; Bat, L.; Yousuf, F. Physicochemical parameters and seasonal variation of coastal water from Balochistan coast, Pakistan. J. Coast. Life Med. 2015, 3, 199–203. [Google Scholar] [CrossRef]
- Jilani, S. Present pollution profile of Karachi coastal waters. J. Coast. Conserv. 2017, 22, 325–332. [Google Scholar] [CrossRef]
- Richards, L. Diagnosis and improvement of saline and alkali soils. Soil Sci. 1954, 78, 154. [Google Scholar] [CrossRef]
- Otte, M.; Haarsma, M.; Broekman, R.; Rozema, J. Relation between heavy metal concentrations in salt marsh plants and soil. Environ. Pollut. 1993, 82, 13–22. [Google Scholar] [CrossRef]
- Chen, F.; Ma, J.; Akhtar, S.; Khan, Z.I.; Ahmad, K.; Ashfaq, A.; Nawaz, H.; Nadeem, M. Assessment of chromium toxicity and potential health implications of agriculturally diversely irrigated food crops in the semi-arid regions of South Asia. Agric. Water Manag. 2022, 272, 107833. [Google Scholar] [CrossRef]
- Shoukat, E.; Aziz, I.; Ahmed, M.Z.; Abideen, Z.; Khan, M.A. Growth patterns of Phragmites karka under saline conditions depend on the bulk elastic modulus. Crop. Pasture Sci. 2018, 69, 535. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Hadi, S.M.S.; Ahmed, M.Z.; Hameed, A.; Khan, M.A.; Gul, B. Seed germination and seedling growth responses of toothbrush tree (Salvadora persica Linn.) to different interacting abiotic stresses. Flora 2018, 243, 45–52. [Google Scholar] [CrossRef]
- Duarte, B.; Reboreda, R.; Caçador, I. Seasonal variation of extracellular enzymatic activity (EEA) and its influence on metal speciation in a polluted salt marsh. Chemosphere 2008, 73, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Rozema, J.; Otte, M.L.; Broekman, R.; Kamber, G.; Punte, H. The response of Spartina anglica to heavy metal pollution. In Spartina Anglica—A Research Review; HMSO: London, UK, 1990; pp. 63–68. Available online: https://core.ac.uk/download/pdf/61341.pdf#page=69 (accessed on 27 January 2023).
- Abu-Dieyeh, M.H.; Usman, K.; Alduroobi, H.; Al-Ghouti, M. Mercury toxicity: The importance of microbial diversity for improved environmental remediation. In Heavy Metals in the Environment; CRC Press: Boca Raton, FL, USA, 2018; pp. 248–267. [Google Scholar] [CrossRef]
- Petrović, J.V.; Alagić, S.; Milić, S.M.; Tošić, S.B.; Bugarin, M.M. Chemometric characterization of heavy metals in soils and shoots of the two pioneer species sampled near the polluted water bodies in the close vicinity of the copper mining and metallurgical complex in Bor (Serbia): Phytoextraction and biomonitoring contexts. Chemosphere 2021, 262, 127808. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef]
- Briat, J.; Lobreaux, S. Iron transport and storage in plants. Trends Plant Sci. 1997, 2, 187–193. [Google Scholar] [CrossRef]
- Aziz, I.; Mujeeb, A. Halophytes for phytoremediation of hazardous metal(loid)s: A terse review on metal tolerance, bio-indication and hyperaccumulation. J. Hazard. Mater. 2021, 424, 127309. [Google Scholar] [CrossRef]
- Parida, A.K.; Veerabathini, S.K.; Kumari, A.; Agarwal, P.K. Physiological, Anatomical and Metabolic Implications of Salt Tolerance in the Halophyte Salvadora persica under Hydroponic Culture Condition. Front. Plant Sci. 2016, 7, 351. [Google Scholar] [CrossRef]
- Zhu, D.; Wei, Y.; Zhao, Y.; Wang, Q.; Han, J. Heavy Metal Pollution and Ecological Risk Assessment of the Agriculture Soil in Xunyang Mining Area, Shaanxi Province, Northwestern China. Bull. Environ. Contam. Toxicol. 2018, 101, 178–184. [Google Scholar] [CrossRef]
- Markert, B. Multi element analysis in plant material-analytical tools and biological questions. In Biochemistry of Trace Elements; Adriano, D.C., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 401–428. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010; p. 01375. [Google Scholar]
- Lewicki, S.; Zdanowski, R.; Krzyżowska, M.; Lewicka, A.; Dębski, B.; Niemcewicz, M.; Goniewicz, M. The role of Chromium III in the organism and its possible use in diabetes and obesity treatment. Ann. Agric. Environ. Med. 2014, 21, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Himelblau, E.; Amasino, R.M. Nutrients mobilized from leaves of Arabidopsis thaliana during leaf senescence. J. Plant Physiol. 2001, 158, 1317–1323. [Google Scholar] [CrossRef]
- Soldatova, N.A.; Khryanin, V.N. Influence of lead salts on growing processes in plants Cannabis sativa L. J. News PGPU Sect. Young Sci. 2008, 6, 215–218. [Google Scholar]
- Mamatha, P.; Sugali, S.; Swamy, A.V.N.; Boyina, R.P.R. Quantitative and risk analysis of heavy metals in selected leafy vegetables. Der. Pharma Chem. 2014, 6, 179–185. [Google Scholar]
- Lasat, M.M. Phytoextraction of Toxic Metals: A review of biological mechanisms. J. Environ. Qual. 2002, 31, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Butcher, D.J. Phytoremediation of Lead in Soil: Recent Applications and Future Prospects. Appl. Spectrosc. Rev. 2009, 44, 123–139. [Google Scholar] [CrossRef]
- Påhlsson, A.-M.B. Toxicity of heavy metals (Zn, Cu, Cd, Pb) to vascular plants. Water Air Soil Pollut. 1989, 47, 287–319. [Google Scholar] [CrossRef]
- Kučera, T.; Horáková, H.; Šonská, A. Toxic metal ions in photoautotrophic organisms. Photosynthetica 2008, 46, 481–489. [Google Scholar] [CrossRef]
- Ismail, S.; Saifullah, S.M.; Khan, S.H. Bio-geochemical studies of Indus delta mangrove ecosystem through heavy metal as-sessment. Pak. J. Bot. 2014, 46, 1277–1285. Available online: https://www.pakbs.org/pjbot/PDFs/46(4)/17.pdf (accessed on 27 January 2023).
- MacFarlane, G.; Pulkownik, A.; Burchett, M. Accumulation and distribution of heavy metals in the grey mangrove, Avicennia marina (Forsk.)Vierh.: Biological indication potential. Environ. Pollut. 2003, 123, 139–151. [Google Scholar] [CrossRef]
- Kaewtubtim, P.; Meeinkuirt, W.; Seepom, S.; Pichtel, J. Heavy metal phytoremediation potential of plant species in a man-grove ecosystem in Pattani Bay, Thailand. Appl. Ecol. Environ. Res. 2016, 14, 367–382. Available online: http://www.aloki.hu/pdf/1401_367382.pdf (accessed on 27 January 2023). [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Negi, S. Bioaccumulation of heavy metals in Salvadora persica Linn. Pharma Innov. J. 2018, 7, 933–936. [Google Scholar]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.; Foster, K.J.; et al. Energy costs of salt tolerance in crop plants. New Phytol. 2019, 225, 1072–1090. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Gorai, M.; Ennajeh, M.; Khemira, H.; Neffati, M. Combined effect of NaCl-salinity and hypoxia on growth, photosynthesis, water relations and solute accumulation in Phragmites australis plants. Flora 2010, 205, 462–470. [Google Scholar] [CrossRef]
- Shabala, S.; Mackay, A. Ion transport in halophytes. In Advances in Botanical Research; Kader, J.C., Delseny, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 57, pp. 151–199. [Google Scholar] [CrossRef]
- Weis, J.S.; Weis, P. Metal uptake, transport and release by wetland plants: Implications for phytoremediation and restoration. Environ. Int. 2004, 30, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Manousaki, E.; Kalogerakis, N. Halophytes—An Emerging Trend in Phytoremediation. Int. J. Phytoremediat. 2011, 13, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Lokhande, V.H.; Suprasanna, P. Prospects of halophytes in understanding and managing abiotic stress tolerance. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: London, UK, 2012; pp. 29–56. [Google Scholar] [CrossRef]
- Rucińska-Sobkowiak, R. Water relations in plants subjected to heavy metal stresses. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil | Root | Leaves | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sites | Metal | Winter | Summer | Post Monsoon | Winter | Summer | Post Monsoon | Winter | Summer | Post Monsoon |
| Site I | Cr | 19.6 ± 0.6 a | 9.93 ± 0.2 b | 3.70 ± 0.1 c | 1.25 ± 0.03 a | 3.09 * ± 0.0 b | 1.25 ± 0.1 a | 3.37 * ± 1.8 a | 0.01 ± 0.0 c | 0.65 ± 0.1 b |
| Site II | 23.2 ± 1.5 a | 9.88 ± 0.2 b | 5.84 ± 0.2 b | 1.25 ± 0.0 a | 2.04 * ± 0.03 b | 0.90 ± 0.2 a | 0.50 * ± 0.1 a | 0.45 * ± 0.1 a | 0.01 ± 0.0 b | |
| Site III | 35.9 * ± 0.3 a | 8.8 ± 0.04 b | 19.9 ± 0.3 c | 2.64 ± 0.1 a | 1.09 ± 0.2 b | 3.03 * ± 0.02 a | 2.22 * ± 0.4 b | 2.15 ± 0.2 b | 1.35 ± 0.3 a | |
| Site I | Pb | 4.27 ± 0.3 a | 12.3 ± 0.9 b | 7.94 ± 0.2 c | 1.69 ± 0.2 a | 1.54 ± 0.03 a | 13.7 *± 0.5 b | 2.72 ± 0.1 a | 4.66 * ± 0.2 b | 2.05 ± 0.3 a |
| Site II | 16.4 ± 1.4 a | 5.29 ± 0.1 b | 9.89 ± 0.1 c | 0.85 ± 0.03 a | 0.01 ± 0.0 a | 3.09 * ± 0.3 b | 2.7 * ± 0.02 a | 1.10 ± 0.3 b | 1.55 ± 0.2 b | |
| Site III | 32.5 * ± 0.2 a | 2.74 ± 0.4 b | 64.7 ± 0.2 c | 2.04 ± 0.1 a | 3.02 ± 0.4 a | 8.19 * ± 0.7 b | 1.45 ± 0.3 a | 1.37 ± 03 a | 2.80 * ± 0.3 b | |
| Site I | Cu | 2.70 ± 0.4 a | 4.04 ± 0.1 b | 2.47 ± 0.04 a | 4.53 ± 0.1 a | 2.54 ± 0.03 b | 7.63 * ± 0.03 c | 5.82 * ± 0.1 a | 1.69 ± 0.1 b | 2.48 ± 0.2 c |
| Site II | 10.9 ± 0.6 a | 5.64 ± 0.1 b | 3.81 ± 0.04 c | 13.5 * ± 1.5 a | 2.99 ± 0.1 b | 6.23 ± 0.03 b | 4.27 * ± 1.1 a | 2.8 ± 0.01 ab | 1.16 ± 0.1 b | |
| Site III | 23.3 ± 0.4 a | 4.89 ± 0.1 b | 23.4 ± 0.2 a | 3.19 ± 0.0 a | 2.33 ± 0.1 b | 3.81 * ± 0.1 c | 12.7 * ± 4.7 b | 3.52 ± 0.2 a | 3.56 ± 0.3 a | |
| Site I | Mn | 11.6 ± 0.5 a | 228.5 * ± 0.9 b | 109 ± 1.4 c | 3.64 ± 0.03 a | 12.5 * ± 0.9 b | 6.48 ± 0.3 c | 9.77 ± 2.8 a | 28.7 * ± 4.1 b | 20.1 ± 1.6 ab |
| Site II | 216 ± 0.7 a | 230.5 * ± 0.3 b | 204.5 ± 1.1 c | 4.52 ± 0.04 a | 15.02 * ± 0.1 b | 6.48 ± 0.3 c | 9.62 ± 2.1 a | 25.4 ± 0.6 ab | 54.4 * ± 14.4 b | |
| Site III | 268 * ± 1.1 a | 161.2 ± 0.4 b | 201.9 ± 0.1 c | 5.85 ± 0.14 | 6.44 * ± 0.3 | 5.87 ± 0.08 | 9.36 ± 1.5 a | 30.7 * ± 3.9 b | 25.0 ± 2.3 b | |
| Site I | Zn | 13.7 ± 1.9 a | 33.4 ± 3.7 b | 47.4 * ± 0.8 c | 24.41 ± 0.3 a | 13.95 ± 1.1 b | 33.4 * ± 0.3 c | 23.2 ± 1.6 ab | 26.4 * ± 3.1 a | 13.8 ± 3.0 b |
| Site II | 65.6 ± 3.3 a | 11.5 ± 2.0 b | 5.78 ± 0.5 b | 50.6 * ±11 a | 15.44 ± 2.0 b | 30.4 ± 0.3 ab | 30.7 ± 0.4 a | 40.2 * ± 1.0 a | 17.6 ± 4.2 b | |
| Site III | 81.2 ± 0.7 a | 10.5 ± 0.9 b | 105.4 * ± 0.6 c | 26.4 * ± 0.9 a | 13.37 ± 0.9 b | 19.1 ± 1.1 c | 27.5 ± 0.9 a | 23.0 ± 1.7 a | 39.8 * ± 0.4 b | |
| Site I | Fe | 5516.3 ± 144 a | 15,016 * ± 39 b | 5178.6 ± 115 a | 919 * ± 13 a | 162.0 ± 4.4 b | 115.9 ± 0.7 c | 40.2 ± 3.9 a | 217 * ± 16 b | 168.4 ± 16.2 b |
| Site II | 14,492.8 ± 692 | 14,793.5 * ± 321 | 13,697.7 ± 10.9 | 55.2 ± 13 a | 113.3 ± 13.7 b | 205.6 * ± 0.6 c | 120.9 ± 21 a | 152 * ± 2.2 a | 147.7 ± 47.0 a | |
| Site III | 17,389.5 * ± 171 a | 9354.7 ± 82 b | 14,201.3 ± 257 c | 228.9 ± 1.7 a | 199.2 ± 2.6 b | 243.4 * ± 9.8 a | 148.8 ± 32 b | 159.7 * ± 24 b | 126.1 ± 3.0 a | |
| Soil | Root | Leaves | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sites | Metal | Winter | Summer | Post Monsoon | Winter | Summer | Post Monsoon | Winter | Summer | Post Monsoon |
| Site I | Na+ | 2.96 ± 0.1 a | 2.82 ± 0.1 a | 9.32 ± 0.2 b | 44.7 ± 0.1 a | 9.59 ± 0.45 b | 111.6 ± 7.2 c | 11.9 ± 4.7 a | 24.4 ± 1.5 a | 90.5 ± 3.5 b |
| Site II | 1.19 ± 0.0 a | 13.5 ± 0.5 b | 8.16 ± 0.2 c | 11.0 ± 0.9 a | 11.5 ± 2.0 a | 30.6 ± 0.7 b | 10.6 ± 0.7 | 34.4 ± 4.3 | 53.6 ± 26.5 | |
| Site III | 2.01 ± 0.0 a | 44.4 ± 5.8 b | 13.8 ± 1.1 a | 11.0 ± 0.01 a | 5.68 ± 0.5 b | 8.61 ± 1.2 ab | 40.6 ± 7.1 | 38.2 ± 2.1 | 40.7 ± 0.7 | |
| Site I | K+ | 0.28 ± 0.01 a | 0.85 ± 0.0 b | 0.65 ± 0.0 c | 7.88 ± 0.2 ab | 5.64 ± 0.5 a | 17.8 ± 4.3 b | 4.19 ± 0.3 a | 11.3 ± 0.5 b | 10.32 ± 1.1 b |
| Site II | 2.15 ± 0.3 | 1.91 ± 0.5 | 2.75 ± 0.01 | 9.26 ± 0.3 a | 6.57 ± 0.0 a | 14.1 ± 1.1 b | 7.23 ± 0.3 a | 9.39 ± 0.6 b | 8.33 ± 0.1 ab | |
| Site III | 2.66 ± 0.0 a | 1.22 ± 0.1 a | 6.52 ± 1.1 b | 7.83 ± 0.1 a | 2.82 ± 0.01 b | 7.38 ± 0.6 a | 9.26 ± 0.9 | 9.39 ± 0.6 | 10.34 ± 1.1 | |
| Sites | Parameter | Winter | Summer | Post monsoon |
|---|---|---|---|---|
| Site I | ΨWo | 1.2 ± 0.1 a | 1.65 * ± 0.2 b | 1.1 ± 0.1 a |
| Site II | 1.8 * ± 0.2 b | 1.4 ± 0.2 a | 1.6 ± 0.2 b | |
| Site III | 1.5 ± 0.3 a | 2.0 * ± 0.04 b | 1.8 ± 0.1 ab | |
| Site I | ΨSo | 1.8 ± 0.3 a | 2.3 * ± 0.2 b | 1.75 ± 0.2 c |
| Site II | 2.3 ± 0.2 b | 1.9 ± 0.1 a | 2.4 * ± 0.3 b | |
| Site III | 2.5 * ± 0.2 b | 1.8 ± 0.4 a | 2.4 * ± 0.2 b | |
| Site I | ΨTLP | 2.7 ± 0.4 a | 3.2 * ± 0.1 b | 2.8 ± 0.04 a |
| Site II | 3.6 * ± 0.4 b | 2.8 ± 0.2 a | 3.4 ± 0.2 b | |
| Site III | 3.8 * ± 0.3 b | 2.8 ± 0.1 a | 3.6 ± 0.2 b | |
| Site I | ε | 6.4 ± 0.5 a | 7.0 ± 0.9 b | 6.6 ± 1.4 c |
| Site II | 6.2 ± 0.2 b | 5.0 ± 0.3 a | 4.8 ± 0.3 a | |
| Site III | 7.6 * ± 1.1 c | 6.1 ± 0.4 b | 5.5 ± 0.3 a | |
| Site I | SUC | 3.1 ± 0.5 b | 2.9 ± 0.2 a | 3.0 ± 0.4 b |
| Site II | 4.2 ± 0.2 a | 5.5 * ± 0.3 b | 4.9 ± 0.3 ab | |
| Site III | 3.6 ± 0.2 a | 4.7 * ± 0.3 c | 4.1 ± 0.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mujeeb, A.; Abideen, Z.; Aziz, I.; Sharif, N.; Hussain, M.I.; Qureshi, A.S.; Yang, H.-H. Phytoremediation of Potentially Toxic Elements from Contaminated Saline Soils Using Salvadora persica L.: Seasonal Evaluation. Plants 2023, 12, 598. https://doi.org/10.3390/plants12030598
Mujeeb A, Abideen Z, Aziz I, Sharif N, Hussain MI, Qureshi AS, Yang H-H. Phytoremediation of Potentially Toxic Elements from Contaminated Saline Soils Using Salvadora persica L.: Seasonal Evaluation. Plants. 2023; 12(3):598. https://doi.org/10.3390/plants12030598
Chicago/Turabian StyleMujeeb, Amtul, Zainul Abideen, Irfan Aziz, Nadia Sharif, Muhammad Iftikhar Hussain, Asad Sarwar Qureshi, and Hsi-Hsien Yang. 2023. "Phytoremediation of Potentially Toxic Elements from Contaminated Saline Soils Using Salvadora persica L.: Seasonal Evaluation" Plants 12, no. 3: 598. https://doi.org/10.3390/plants12030598