
Transcriptome Sequencing of Broussonetia papyrifera Leaves Reveals Key Genes Involved in Flavonoids Biosynthesis
Abstract
:1. Introduction
2. Results
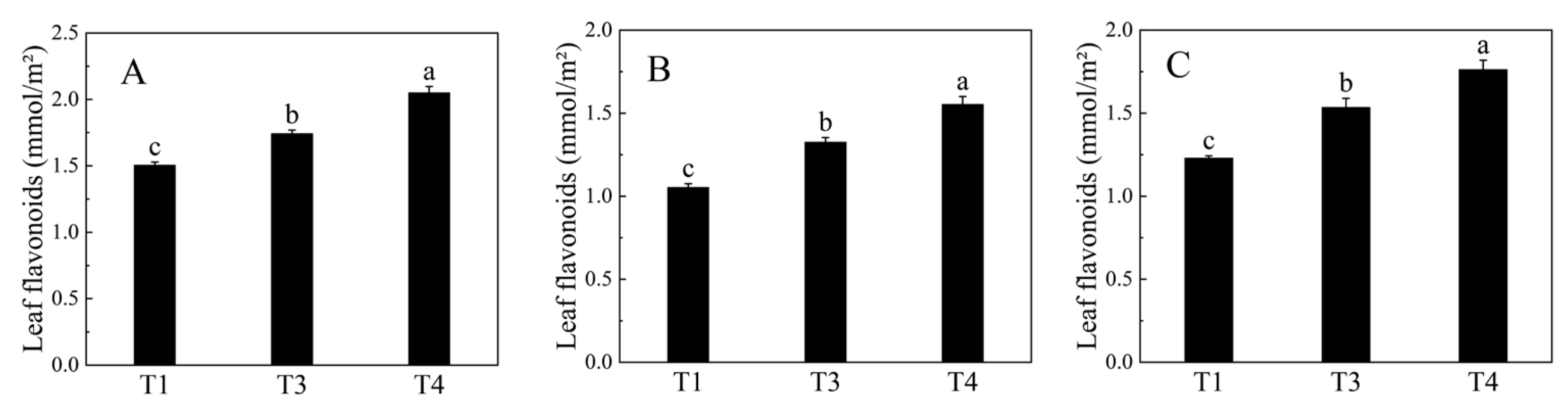
2.1. Changes in Total Flavonoid Content during B. papyrifera Leaves Development
2.2. RNA-Seq, De Novo Assembly of the Transcripts and Annotation
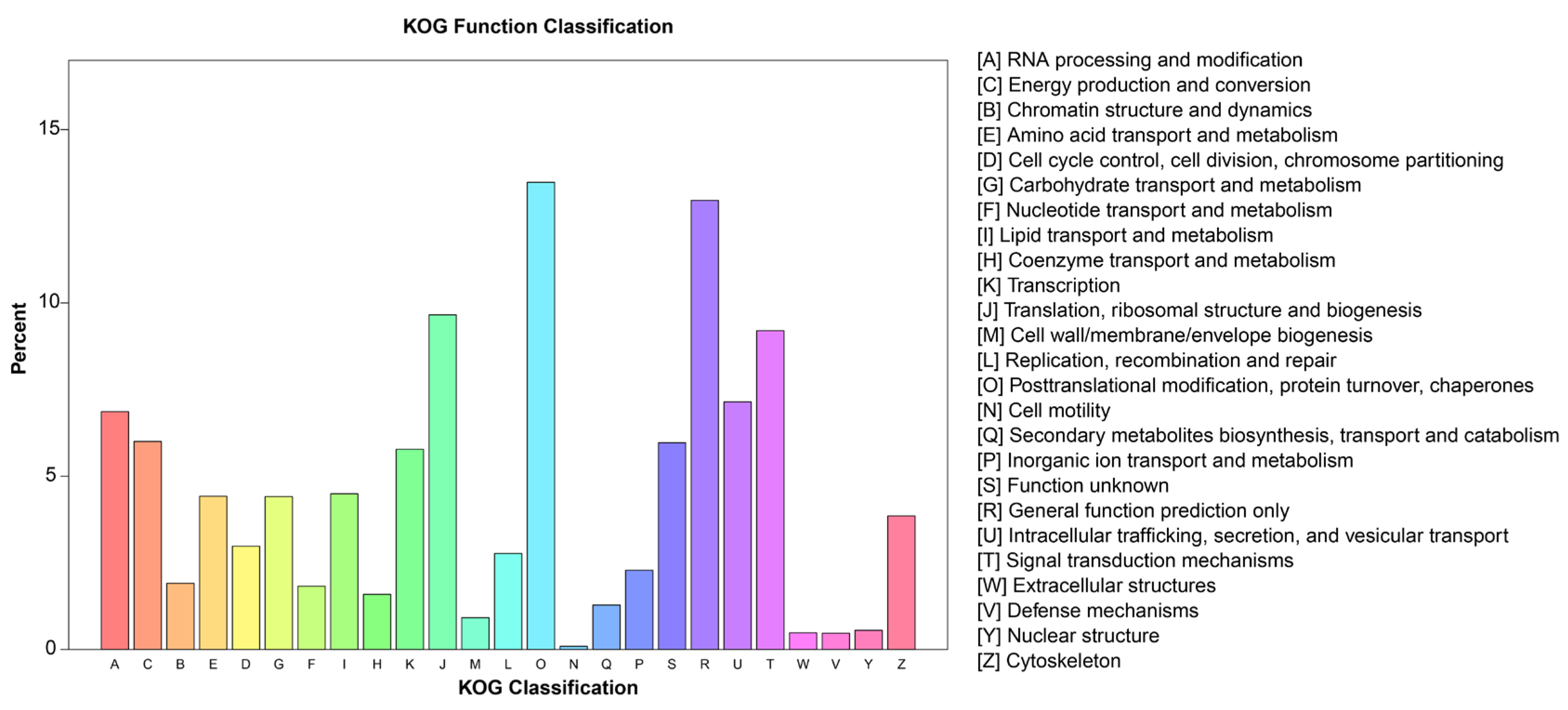
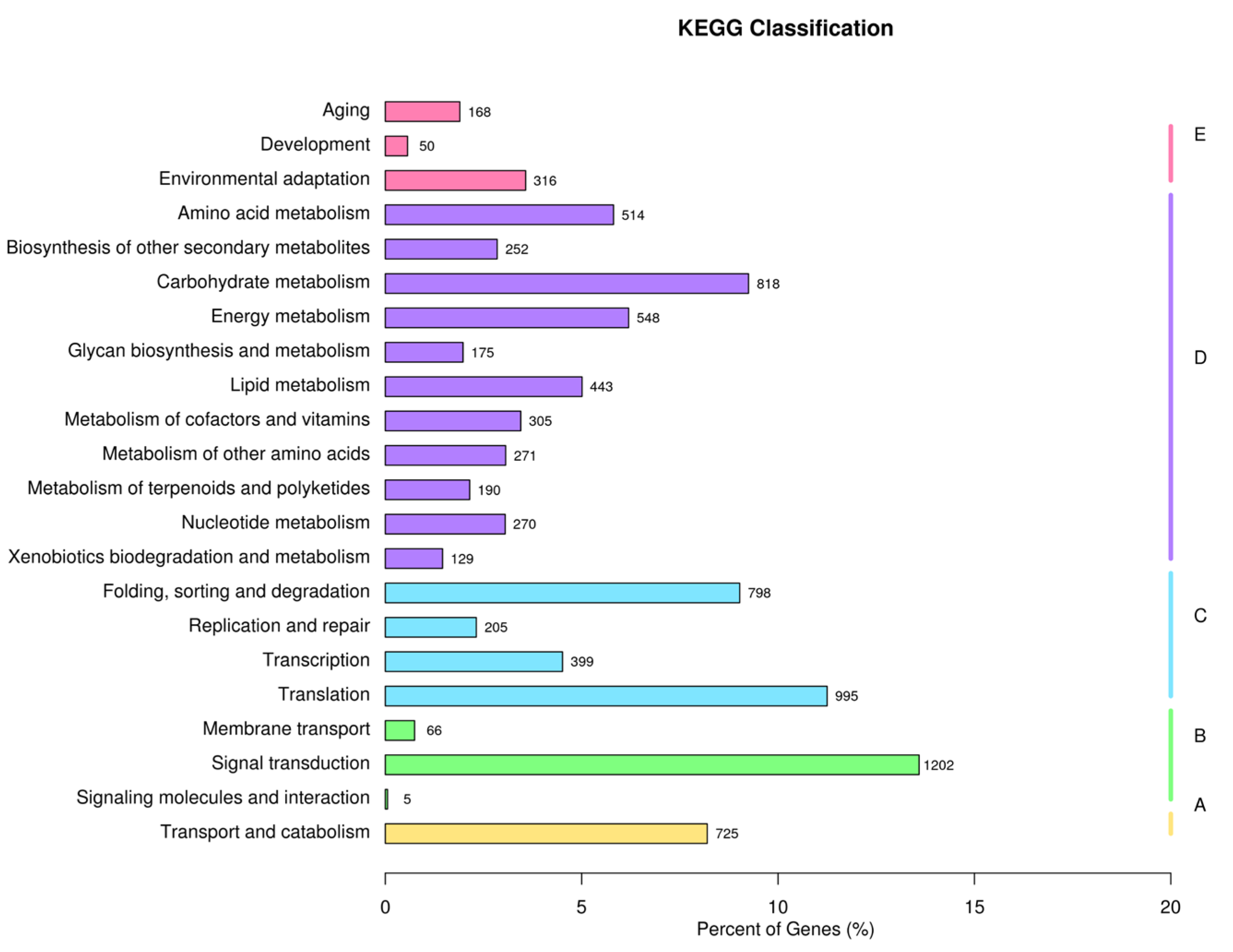
2.3. Functional Annotation and Classification
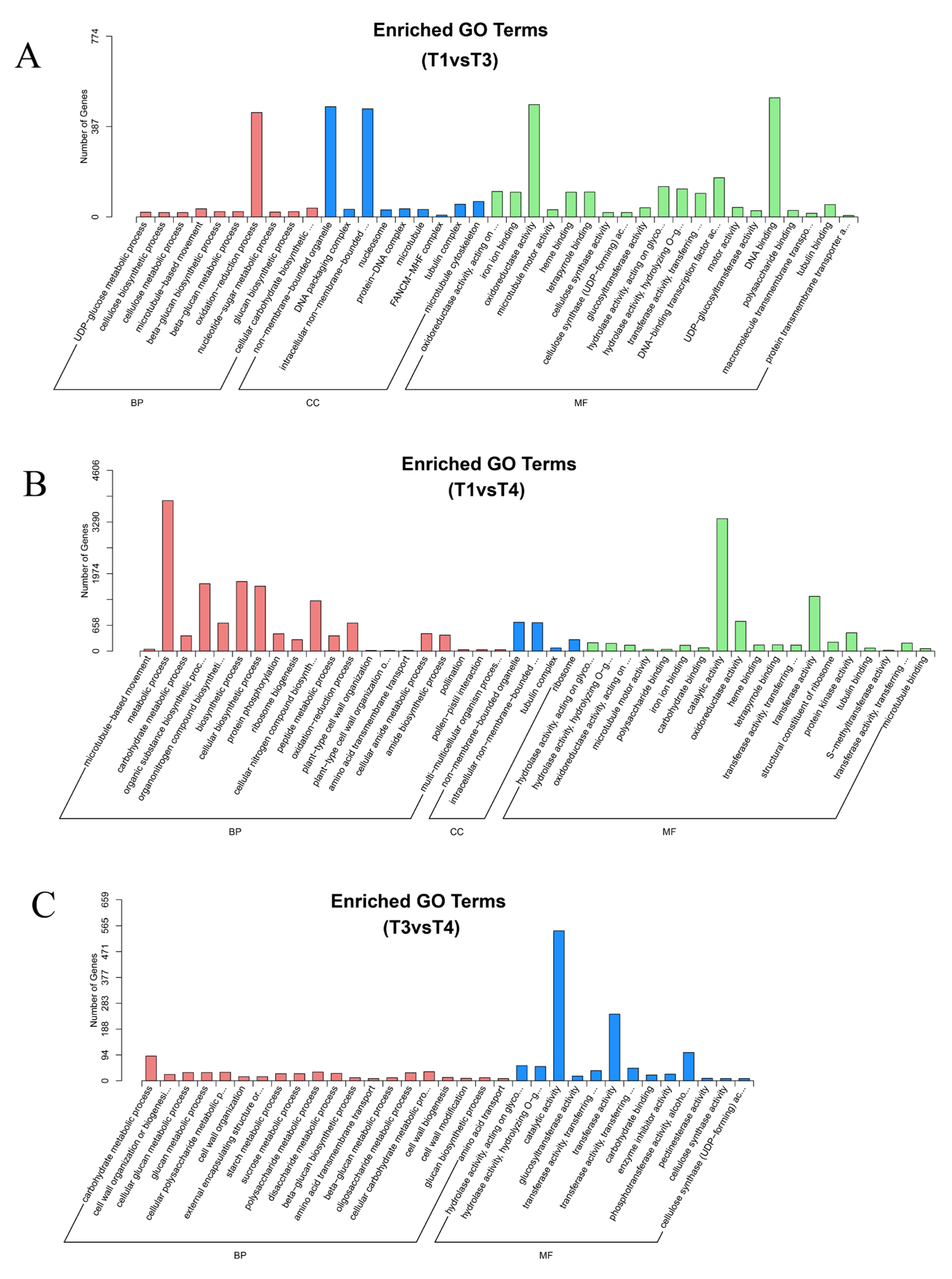
2.4. DEG Identification
2.5. Validation of qRT-PCR
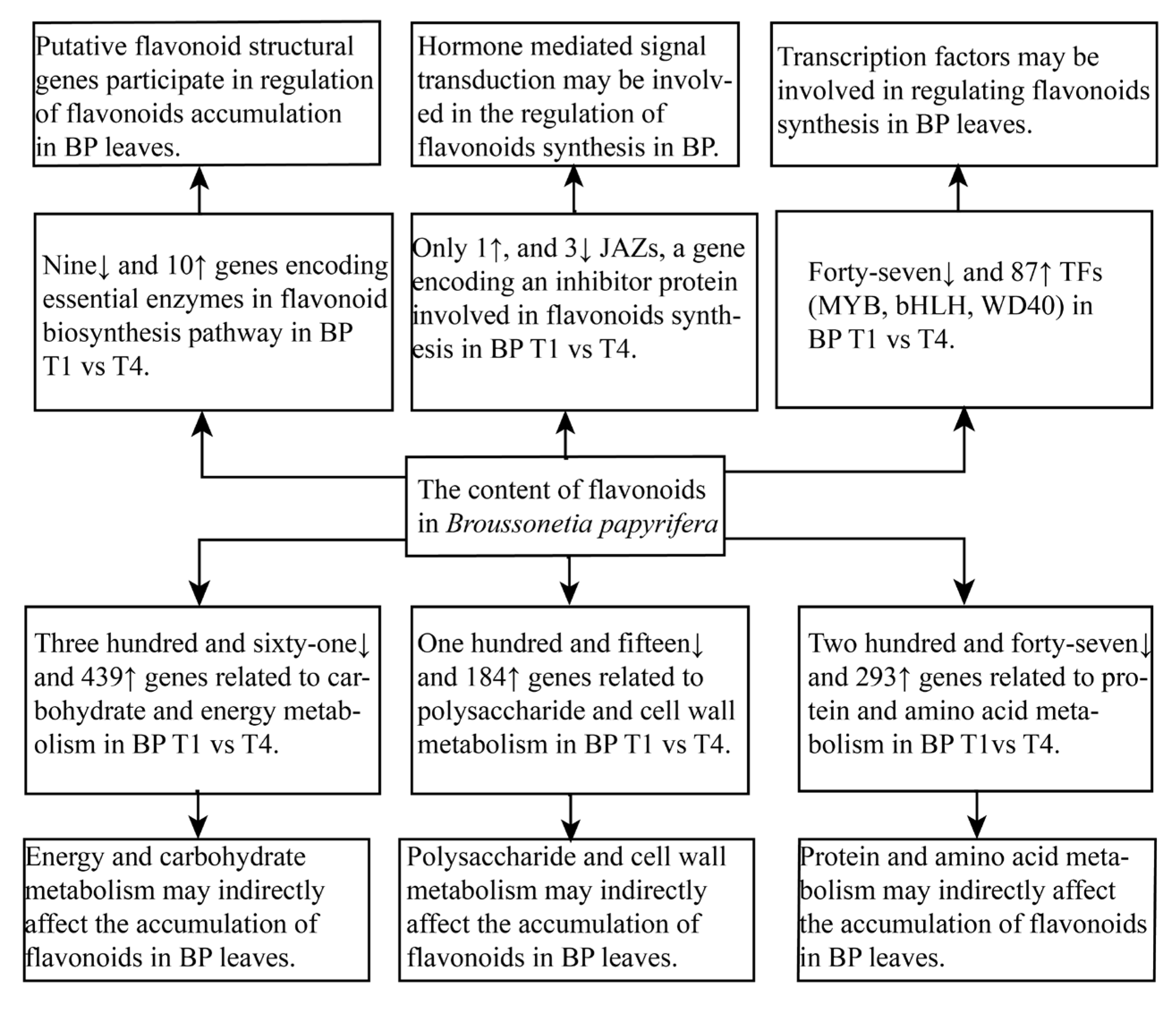
3. Discussion
3.1. Putative Genes Related to Flavonoid Biosynthesis and Transport
3.2. Genes Related to Signal Transduction
3.3. Transcription Factors
3.4. Other Genes That May Affect the Synthesis of Flavonoids in Broussonetia papyrifera
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of Total Flavonoids
4.3. RNA Extraction, cDNA Preparation, and RNA-Seq
4.4. Transcript Assembly and Gene Expression Analysis
4.5. Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, W.; Li, H.; Zheng, X. Researches of constituents of Broussonetia papyrifera. Chin. J. New Drugs 2008, 17, 272. [Google Scholar]
- Gao, Y.; Qiu, Y.; Gao, L.; Qu, J. Depressive effect of alcohol extract and flavonoids of Broussonetia papyrifera leaves on isolated auricles. Acta Acad. Med. Taishan 1988, 125–129. [Google Scholar]
- Yang, X.; He, R.; Cao, Y. Study on Protective Effects of TFBP on Human Immortalized Keratinocytes line HaCaT. J. Environ. Health 2004, 134–136. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Bavaresco, L.; Falginella, L.; Gonçalves, M.I.V.Z.; Di Gaspero, G. BerryPhenolics in Grape Berry and Key Antioxidants. In The Biochemistry of the Grape; University of Minho: Braga, Portugal, 2012; p. 303. [Google Scholar]
- He, F.; Mu, L.; Yan, G.-L.; Liang, N.-N.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Biosynthesis of Anthocyanins and Their Regulation in Colored Grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, J.-L.; Austin, M.B.; Stewart, C.; Noel, J.P. Structure and Function of Enzymes Involved in the Biosynthesis of Phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, J.-L.; Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure of Chalcone Synthase and the Molecular Basis of Plant Polyketide Biosynthesis. Nat. Struct. Biol. 1999, 6, 10. [Google Scholar]
- Jez, J.M.; Noel, J.P. Mechanism of Chalcone Synthase. J. Biol. Chem. 2000, 275, 39640–39646. [Google Scholar] [CrossRef] [Green Version]
- Jez, J.M.; Bowman, M.E.; Noel, J.P. Structure and Mechanism of the Evolutionarily Unique Plant Enzyme Chalcone Isomerase. Nat. Struct. Biol. 2000, 7, 6. [Google Scholar]
- Martens, S.; Mithöfer, A. Flavones and Flavone Synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef]
- Martens, S.; Forkmann, G.; Matern, U.; Lukačin, R. Cloning of Parsley Flavone Synthase I. Phytochemistry 2001, 58, 43–46. [Google Scholar] [CrossRef]
- Gebhardt, Y.H.; Witte, S.; Steuber, H.; Matern, U.; Martens, S. Evolution of Flavone Synthase I from Parsley Flavanone 3 β -Hydroxylase by Site-Directed Mutagenesis. Plant Physiol. 2007, 144, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, Y.; Witte, S.; Forkmann, G.; Lukačin, R.; Matern, U.; Martens, S. Molecular Evolution of Flavonoid Dioxygenases in the Family Apiaceae. Phytochemistry 2005, 66, 1273–1284. [Google Scholar] [CrossRef]
- Rosado, A.; Hicks, G.R.; Norambuena, L.; Rogachev, I.; Meir, S.; Pourcel, L.; Zouhar, J.; Brown, M.Q.; Boirsdore, M.P.; Puckrin, R.S.; et al. Sortin1-Hypersensitive Mutants Link Vacuolar-Trafficking Defects and Flavonoid Metabolism in Arabidopsis Vegetative Tissues. Chem. Biol. 2011, 18, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zang, C.; Ge, H.; Zhang, J.; Grierson, D.; Yin, X.; Chen, K. Involvement of PAL, C4H, and 4CL in Chilling Injury-Induced Flesh Lignification of Loquat Fruit. Horts 2017, 52, 127–131. [Google Scholar] [CrossRef]
- Wei, Y.; Zhou, D.; Peng, J.; Pan, L.; Tu, K. Hot Air Treatment Induces Disease Resistance through Activating the Phenylpropanoid Metabolism in Cherry Tomato Fruit. J. Agric. Food Chem. 2017, 65, 8003–8010. [Google Scholar] [CrossRef]
- Li, Y.; Sun, L.; Wang, H.; Gao, R.; Zhang, J.; Hu, B.; Chang, Y. Cloning and Expression Analysis of Phenylalanine Ammonia-Lyase (PAL) Gene Family and Cinnamate 4-Hydroxylase (C4H) from Dryopteris Fragrans. Biologia 2015, 70, 606–614. [Google Scholar] [CrossRef]
- Abdollahi Mandoulakani, B.; Eyvazpour, E.; Ghadimzadeh, M. The Effect of Drought Stress on the Expression of Key Genes Involved in the Biosynthesis of Phenylpropanoids and Essential Oil Components in Basil (Ocimum basilicum L.). Phytochemistry 2017, 139, 1–7. [Google Scholar] [CrossRef]
- Liu, L. The Effects of UV Radiation on Flavonoid Accumulation and Gene Expression in Sauvignon Blanc Grape Berries. Ph.D. Thesis, Lincoln University, Lincoln, UK, 2014. [Google Scholar]
- Ramsay, N.A.; Walker, A.R.; Mooney, M.; Gray, J.C. Two Basic-Helix-Loop-Helix Genes (MYC-146 and GL3) from Arabidopsis Can Activate Anthocyanin Biosynthesis in a White-FLowered Matthiola Incana Mutant. Plant Mol. Biol. 2003, 52, 679–688. [Google Scholar] [CrossRef]
- Takos, A.M.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-Induced Expression of a MYB Gene Regulates Anthocyanin Biosynthesis in Red Apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [Green Version]
- Hichri, I.; Heppel, S.C.; Pillet, J.; Léon, C.; Czemmel, S.; Delrot, S.; Lauvergeat, V.; Bogs, J. The Basic Helix-Loop-Helix Transcription Factor MYC1 Is Involved in the Regulation of the Flavonoid Biosynthesis Pathway in Grapevine. Mol. Plant 2010, 3, 509–523. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A Colorful Model for the Regulation and Evolution of Biochemical Pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, C.; Zhang, M.; Yin, X.; Xu, C.; Chen, K. The MrWD40-1 Gene of Chinese Bayberry (Myrica Rubra) Interacts with MYB and BHLH to Enhance Anthocyanin Accumulation. Plant Mol. Biol. Rep. 2013, 31, 1474–1484. [Google Scholar] [CrossRef]
- Ben-Simhon, Z.; Judeinstein, S.; Nadler-Hassar, T.; Trainin, T.; Bar-Ya’akov, I.; Borochov-Neori, H.; Holland, D. A Pomegranate (Punica granatum L.) WD40-Repeat Gene Is a Functional Homologue of Arabidopsis TTG1 and Is Involved in the Regulation of Anthocyanin Biosynthesis during Pomegranate Fruit Development. Planta 2011, 234, 865–881. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The Flavonoid Biosynthetic Pathway in Arabidopsis: Structural and Genetic Diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhu, J.; Zhao, S.; Hou, Y.; Li, F.; Tai, Y.; Wan, X.; Wei, C. Transcriptome Profiling Using Single-Molecule Direct RNA Sequencing Approach for In-Depth Understanding of Genes in Secondary Metabolism Pathways of Camellia Sinensis. Front. Plant Sci. 2017, 8, 1205. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J. Flavonoid Transport Mechanisms: How to Go, and with Whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, Perception, Signal Transduction and Action in Plant Stress Response, Growth and Development. An Update to the 2007 Review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, C.-T.; Wang, C.Y.; Chen, P. Resveratrol Content in Strawberry Fruit Is Affected by Preharvest Conditions. J. Agric. Food Chem. 2007, 55, 8269–8274. [Google Scholar] [CrossRef]
- Wang, K.; Jin, P.; Cao, S.; Shang, H.; Yang, Z.; Zheng, Y. Methyl Jasmonate Reduces Decay and Enhances Antioxidant Capacity in Chinese Bayberries. J. Agric. Food Chem. 2009, 57, 5809–5815. [Google Scholar] [CrossRef]
- Flores, G.; Blanch, G.P.; del Castillo, M.L.R. Effect of Postharvest Methyl Jasmonate Treatment on Fatty Acid Composition and Phenolic Acid Content in Olive Fruits during Storage: MJ-Induced Bioactive Compounds in Olives. J. Sci. Food Agric. 2017, 97, 2767–2772. [Google Scholar] [CrossRef]
- Luo, X.; Zhu, D.; Huang, Y.; Li, M.; Yao, P.; Gao, F.; Li, C.; Zhao, H. Effects of Methyl Jasmonate Accumulation of Flavonoids and Related Gene Expression of Buckwheat Sprouts. Genom. Appl. Biol. 2015, 34, 1040–1046. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb -Related Genes of the Kyoho Grape (Vitis Labruscana) Regulate Anthocyanin Biosynthesis. Planta 2002, 215, 924–933. [Google Scholar] [CrossRef]
- Lin-Wang, K.; Bolitho, K.; Grafton, K.; Kortstee, A.; Karunairetnam, S.; McGhie, T.K.; Espley, R.V.; Hellens, R.P.; Allan, A.C. An R2R3 MYB Transcription Factor Associated with Regulation of the Anthocyanin Biosynthetic Pathway in Rosaceae. BMC Plant Biol. 2010, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Feller, A.; Hernandez, J.M.; Grotewold, E. An ACT-like Domain Participates in the Dimerization of Several Plant Basic-Helix-Loop-Helix Transcription Factors. J. Biol. Chem. 2006, 281, 28964–28974. [Google Scholar] [CrossRef]
- Li, S. Transcriptional Control of Flavonoid Biosynthesis: Fine-Tuning of the MYB-BHLH-WD40 (MBW) Complex. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef]
- Zhou, F. Amino acid analysis of leaves, flowers and fruits of Broussonetia papyrifera. J. Pharm. Pract. 2005, 23, 154–156. [Google Scholar]
- Liu, J.; Li, Y.; Zhou, M.; Wu, P.; Ma, X.; Li, M. Advances in the Studies of Plant Protein Post-translational Modification. Biotechnol. Bull. 2021, 37, 67–76. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell Wall Hemicellulose Contributes Significantly to Aluminum Adsorption and Root Growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef] [Green Version]
- Chandran, D.; Sharopova, N.; Ivashuta, S.; Gantt, J.S.; VandenBosch, K.A.; Samac, D.A. Transcriptome Profiling Identified Novel Genes Associated with Aluminum Toxicity, Resistance and Tolerance in Medicago Truncatula. Planta 2008, 228, 151–166. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant Expansins: Diversity and Interactions with Plant Cell Walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Pignocchi, C.; Fletcher, J.M.; Wilkinson, J.E.; Barnes, J.D.; Foyer, C.H. The Function of Ascorbate Oxidase in Tobacco. Plant Physiol. 2003, 132, 1631–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Man, Y.; Po, Y.; Sun, N.; Li, R.; JIn, Y. Role of cytoskeleton in plant cell wall formation. Sci. China Life Sci. 2020, 50, 176–186. [Google Scholar]
- Krizek, D.T.; Britz, S.J.; Mirecki, R.M. Inhibitory Effects of Ambient Levels of Solar UV-A and UV-B Radiation on Growth of Cv. New Red Fire Lettuce. Physiol. Plant. 1998, 103, 1–7. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Raw Reads | Clean Reads | Clean Bases | Error (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| T1-1 | 21,325,528 | 20,725,009 | 6.22G | 0.03 | 97.84 | 93.76 | 46.63 |
| T1-2 | 22,271,891 | 21,756,316 | 6.53G | 0.03 | 97.75 | 93.61 | 46.76 |
| T1-3 | 21,793,119 | 21,357,102 | 6.41G | 0.03 | 97.82 | 93.76 | 46.23 |
| T3-1 | 22,174,936 | 21,643,438 | 6.49G | 0.03 | 97.60 | 93.30 | 46.16 |
| T3-2 | 22,126,417 | 21,170,390 | 6.35G | 0.03 | 97.87 | 94.00 | 46.63 |
| T3-3 | 21,308,885 | 20,786,945 | 6.24G | 0.03 | 97.97 | 94.11 | 46.71 |
| T4-1 | 23,421,640 | 22,539,743 | 6.76G | 0.03 | 97.89 | 93.95 | 46.15 |
| T4-2 | 22,434,529 | 21,668,818 | 6.50G | 0.03 | 97.88 | 94.03 | 46.10 |
| T4-3 | 22,656,272 | 22,197,669 | 6.66G | 0.03 | 97.48 | 92.79 | 45.86 |
| 300–500 bp | 500 bp–1 kbp | 1 kbp–2 kbp | >2 kbp | Total | |
|---|---|---|---|---|---|
| Number of Unigenes | 11,342 | 10,629 | 7915 | 11,651 | 41,537 |
| Number of transcripts | 17,930 | 20,939 | 32,898 | 52,898 | 124,575 |
| Min Length | Mean Length | Median Length | Max Length | N50 | N90 | Total Nucleotides | |
|---|---|---|---|---|---|---|---|
| Transcripts | 301 | 2005 | 1705 | 17,100 | 2835 | 1078 | 249,817,198 |
| Genes | 301 | 1515 | 902 | 17,100 | 2568 | 592 | 62,948,261 |
| Datebase | Number of Unigenes | Percentage |
|---|---|---|
| NR | 25,804 | 62.12 |
| NT | 19,586 | 47.15 |
| KO | 10,361 | 24.94 |
| Swiss Prot | 22,354 | 53.81 |
| PFAM | 21,289 | 51.25 |
| GO | 21,287 | 51.24 |
| KOG | 8480 | 20.41 |
| Annotated in all Databases | 3358 | 8.08 |
| Annotated in at least one Database | 30,177 | 72.65 |
| Total Unigenes | 41,537 | 100 |
| Compare * | |log2FoldChange| < 2 | Percentage | |log2FoldChange| > 2 | Percentage |
|---|---|---|---|---|
| T1vsT3 | 3190 | 59.00% | 2217 | 41.00% |
| T3vsT4 | 874 | 67.80% | 415 | 32.20% |
| T1vsT4 | 4272 | 45.15% | 5189 | 54.85% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Huang, Z.; Li, X.; Zhao, W.; Wang, Y. Transcriptome Sequencing of Broussonetia papyrifera Leaves Reveals Key Genes Involved in Flavonoids Biosynthesis. Plants 2023, 12, 563. https://doi.org/10.3390/plants12030563
Guo P, Huang Z, Li X, Zhao W, Wang Y. Transcriptome Sequencing of Broussonetia papyrifera Leaves Reveals Key Genes Involved in Flavonoids Biosynthesis. Plants. 2023; 12(3):563. https://doi.org/10.3390/plants12030563
Chicago/Turabian StyleGuo, Peng, Ziqi Huang, Xinke Li, Wei Zhao, and Yihan Wang. 2023. "Transcriptome Sequencing of Broussonetia papyrifera Leaves Reveals Key Genes Involved in Flavonoids Biosynthesis" Plants 12, no. 3: 563. https://doi.org/10.3390/plants12030563