
A Proximal Sensor-Based Approach for Clean, Fast, and Accurate Assessment of the Eucalyptus spp. Nutritional Status and Differentiation of Clones
 , ,
, ,
Abstract
:1. Introduction
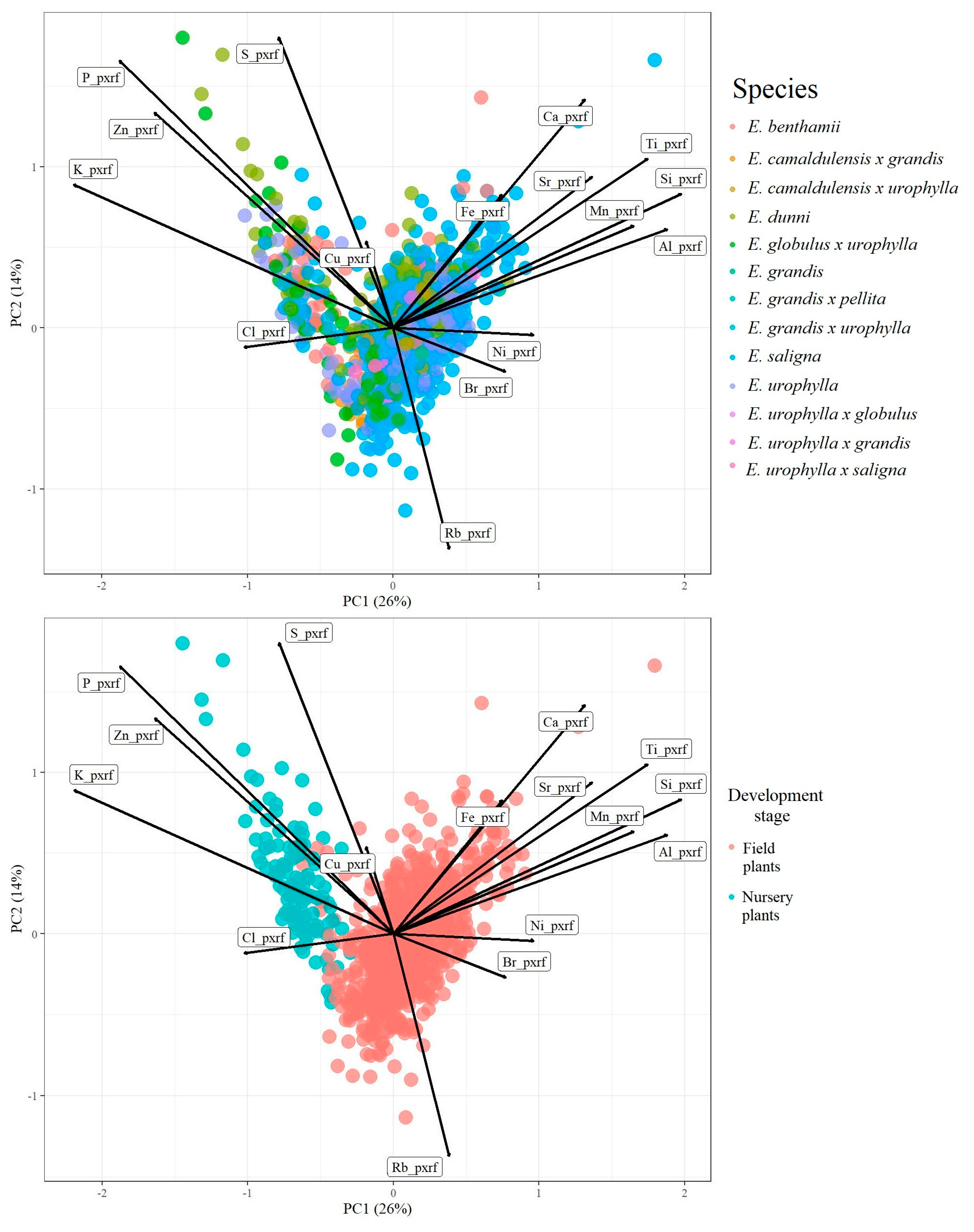
2. Results and Discussion
2.1. Elemental Chemical Characterization of Eucalyptus Leaves Using pXRF Spectrometry
2.2. Validation Performance of Leaf Nutrient Predictions
2.3. Validation of Leaf Nutrient Categorical Predictions
2.4. Variables Importance
2.5. Data Variability versus Prediction Model Accuracy
2.6. Future Applications
3. Materials and Methods
3.1. Eucalyptus Leaves Sampling
3.2. Conventional Analysis of Eucalyptus Leaves
3.3. pXRF Spectrometry Analysis
3.4. Statistical Analyses and Modeling
3.5. Evaluation of Models’ Performance
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IBÁ Brazilian Tree Industry Annual Report. 2021. Available online: https://iba.org/eng/datafiles/publicacoes/relatorios/relatorioiba2021.pdf (accessed on 21 September 2022).
- Vilasboa, J.; Da Costa, C.T.; Fett-Neto, A.G. Environmental Modulation of Mini-Clonal Gardens for Cutting Production and Propagation of Hard- and Easy-to-Root Eucalyptus Spp. Plants 2022, 11, 3281. [Google Scholar] [CrossRef] [PubMed]
- Cagáň, Ľ.; Apacsová Fusková, M.; Hlávková, D.; Skoková Habuštová, O. Essential Oils: Useful Tools in Storage-Pest Management. Plants 2022, 11, 3077. [Google Scholar] [CrossRef] [PubMed]
- Polito, F.; Kouki, H.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic and Antibiofilm Activity of the Essential Oils of Eucalyptus Bicostata, E. Gigantea, E. Intertexta, E. Obliqua, E. Pauciflora and E. Tereticornis. Plants 2022, 11, 3017. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.; Soares, C.; Martins, M.; Sousa, B.; Valente, I.; Pereira, R.; Fidalgo, F. Herbicidal Effects and Cellular Targets of Aqueous Extracts from Young Eucalyptus Globulus Labill. Leaves. Plants 2021, 10, 1159. [Google Scholar] [CrossRef]
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the Bioherbicide Potential of Eucalyptus Globulus Labill: Biochemistry and Effects of Its Aqueous Extract. PLoS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [Green Version]
- Brisola, S.H.; Demarco, D. Stem Anatomical Analysis of Eucalyptus Grandis, E. Urophylla and E. Grandis x Urophylla: Wood Development and Its Industrial Importance. Sci. Florest. 2011, 39, 317–330. [Google Scholar]
- Zhan, N.; Shang, X.; Wang, Z.; Xie, Y.; Liu, G.; Wu, Z. Screening Cellulose Synthesis Related Genes of EgrEXP and EgrHEX in Eucalyptus Grandis. Gene 2022, 824, 146396. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.; Guimarães, P.T.G.; Venegaz, V.H.A. 5a Aproximação—Recomendações Para o Uso de Corretivos e Fertilizantes Em Minas Gerais, 1st ed.; SBCS: Viçosa, Brazil, 1999. [Google Scholar]
- de Oliveira, C.T.; Rozane, D.E.; de Amorim, D.A.; de Souza, H.A.; Fernandes, B.S.; Natale, W. Diagnosis of the Nutritional Status of ‘Paluma’ Guava Trees Using Leaf and Flower Analysis. Rev. Bras. Frutic. 2020, 42, e-132. [Google Scholar] [CrossRef]
- Hanlon, E.A. Elemental Determination by Atomic Absorption Spectrophotometry. In Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Silva, S.H.G.; Ribeiro, B.T.; Guerra, M.B.B.; Carvalho, H.W.P.; Lopes, G.; Carvalho, G.S.; Guilherme, L.R.G.; Resende, M.; Mancini, M.; Curi, N.; et al. pXRF in tropical soils: Methodology, applications, achievements and challenges. Adv. Agron. 2021, 167, 1–62. [Google Scholar] [CrossRef]
- Weindorf, D.C.; Chakraborty, S. Portable X-ray Fluorescence Spectrometry Analysis of Soils. Soil Sci. Soc. Am. J. 2020, 84, 1384–1392. [Google Scholar] [CrossRef]
- Zhu, Y.; Weindorf, D.C. Determination of Soil Calcium Using Field Portable X-Ray Fluorescence. Soil Sci. 2009, 174, 151–155. [Google Scholar] [CrossRef]
- Andrade, R.; Silva, S.H.G.; Weindorf, D.C.; Chakraborty, S.; Faria, W.M.; Guilherme, L.R.G.; Curi, N. Tropical Soil Order and Suborder Prediction Combining Optical and X-Ray Approaches. Geoderma Reg. 2020, 23, e00331. [Google Scholar] [CrossRef]
- Andrade, R.; Silva, S.H.G.; Weindorf, D.C.; Chakraborty, S.; Faria, W.M.; Guilherme, L.R.G.; Curi, N. Micronutrients Prediction via PXRF Spectrometry in Brazil: Influence of Weathering Degree. Geoderma Reg. 2021, 27, e00431. [Google Scholar] [CrossRef]
- Pearson, D.; Chakraborty, S.; Duda, B.; Li, B.; Weindorf, D.C.; Deb, S.; Brevik, E.; Ray, D.P. Water Analysis via Portable X-Ray Fluorescence Spectrometry. J. Hydrol. 2017, 544, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Andrade, R.; Mancini, M.; Teixeira, A.F.d.S.; Silva, S.H.G.; Weindorf, D.C.; Chakraborty, S.; Guilherme, L.R.G.; Curi, N. Proximal Sensor Data Fusion and Auxiliary Information for Tropical Soil Property Prediction: Soil Texture. Geoderma 2022, 422, 115936. [Google Scholar] [CrossRef]
- Mancini, M.; Andrade, R.; Teixeira, A.F.d.S.; Silva, S.H.G.; Weindorf, D.C.; Chakraborty, S.; Guilherme, L.R.G.; Curi, N. Proximal Sensor Data Fusion for Brazilian Soil Properties Prediction: Exchangeable/Available Macronutrients, Aluminum, and Potential Acidity. Geoderma Reg. 2022, 30, e00573. [Google Scholar] [CrossRef]
- Teixeira, A.F.S.; Andrade, R.; Mancini, M.; Silva, S.H.G.; Weindorf, D.C.; Chakraborty, S.; Guilherme, L.R.G.; Curi, N. Proximal Sensor Data Fusion for Tropical Soil Property Prediction: Soil Fertility Properties. J. S. Am. Earth Sci. 2022, 116, 103873. [Google Scholar] [CrossRef]
- Weindorf, D.C.; Bakr, N.; Zhu, Y. Advances in Portable X-Ray Fluorescence (PXRF) for Environmental, Pedological, and Agronomic Applications. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; Volume 128, pp. 1–45. ISBN 978-0-12-802139-2. [Google Scholar]
- Cohen, D.R.; Zissimos, A.M.; Schifano, J.A.; Rutherford, N.F. Biogeochemical Response of Pinus Brutia and Olea Europaea to Lithological Variations and Cu Mineralisation in Cyprus. Sci. Total Environ. 2021, 759, 143434. [Google Scholar] [CrossRef] [PubMed]
- McQueen, K.G.; Alorbi, A.; Schifano, J.A.; Cohen, D.R. Nickel Uptake by Cypress Pine (Callitris Glaucophylla) in the Miandetta Area, Australia: Implications for Use in Biogeochemical Exploration. Minerals 2021, 11, 808. [Google Scholar] [CrossRef]
- McGladdery, C.; Weindorf, D.C.; Chakraborty, S.; Li, B.; Paulette, L.; Podar, D.; Pearson, D.; Kusi, N.Y.O.; Duda, B. Elemental Assessment of Vegetation via Portable X-Ray Fluorescence (PXRF) Spectrometry. J. Environ. Manag. 2018, 210, 210–225. [Google Scholar] [CrossRef]
- Pereira Junior, S.M.; Maihara, V.A.; Moreira, E.G.; Salvador, V.L.R.; Sato, I.M. Determination of Cu, Cr, and As in Preserved Wood (Eucalyptus Sp.) Using x-Ray Fluorescent Spectrometry Techniques. J. Radioanal. Nucl. Chem. 2016, 308, 7–12. [Google Scholar] [CrossRef]
- Costa Junior, G.T.; Nunes, L.C.; Feresin Gomes, M.H.; Almeida, E.; Pereira Carvalho, H.W. Direct Determination of Mineral Nutrients in Soybean Leaves under Vivo Conditions by Portable X-ray Fluorescence Spectroscopy. X-Ray Spectrom. 2020, 49, 274–283. [Google Scholar] [CrossRef]
- Borges, C.S.; Weindorf, D.C.; Carvalho, G.S.; Guilherme, L.R.G.; Takayama, T.; Curi, N.; Lima, G.J.E.O.; Ribeiro, B.T. Foliar Elemental Analysis of Brazilian Crops via Portable X-Ray Fluorescence Spectrometry. Sensors 2020, 20, 2509. [Google Scholar] [CrossRef]
- Towett, E.K.; Shepherd, K.D.; Lee Drake, B. Plant Elemental Composition and Portable X-ray Fluorescence (PXRF) Spectroscopy: Quantification under Different Analytical Parameters. X-Ray Spectrom. 2016, 45, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Soares, T.M.; Carvalho, H.W.P.; Almeida, E.; Costa, G.T.; Pavinato, P.S. Phosphorus Quantification in Sugar Cane (Saccharum Officinarum) Leaves in Vivo by Portable X-ray Fluorescence Spectroscopy. ACS Agric. Sci. Technol. 2021, 1, 479–487. [Google Scholar] [CrossRef]
- Mancini, M.; Weindorf, D.C.; Silva, S.H.G.; Chakraborty, S.; Teixeira, A.F.d.S.; Guilherme, L.R.G.; Curi, N. Parent Material Distribution Mapping from Tropical Soils Data via Machine Learning and Portable X-Ray Fluorescence (PXRF) Spectrometry in Brazil. Geoderma 2019, 354, 113885. [Google Scholar] [CrossRef]
- Mulenga, C.; Clarke, C.; Meincken, M. Bioaccumulation of Cu, Fe, Mn and Zn in Native Brachystegia Longifolia Naturally Growing in a Copper Mining Environment of Mufulira, Zambia. Environ. Monit. Assess. 2022, 194, 8. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Raven, J.A. Nitrogen and Sulfur Assimilation in Plants and Algae. Aquat. Bot. 2014, 118, 45–61. [Google Scholar] [CrossRef]
- Andrade, R.; Faria, W.M.; Silva, S.H.G.; Chakraborty, S.; Weindorf, D.C.; Mesquita, L.F.; Guilherme, L.R.G.; Curi, N. Prediction of Soil Fertility via Portable X-Ray Fluorescence (PXRF) Spectrometry and Soil Texture in the Brazilian Coastal Plains. Geoderma 2020, 357, 113960. [Google Scholar] [CrossRef]
- Silva, S.H.G.; Teixeira, A.F.d.S.; de Menezes, M.D.; Guilherme, L.R.G.; Moreira, F.M.d.S.; Curi, N. Multiple Linear Regression and Random Forest to Predict and Map Soil Properties Using Data from Portable X-Ray Fluorescence Spectrometer (PXRF). Ciência E Agrotecnologia 2017, 41, 648–664. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R.; Gregory, P.J. Managing the Nutrition of Plants and People. Appl. Environ. Soil Sci. 2012, 2012, 104826. [Google Scholar] [CrossRef] [Green Version]
- Huber, D.; Römheld, V.; Weinmann, M. Relationship between Nutrition, Plant Diseases and Pests. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 283–298. ISBN 978-0-12-384905-2. [Google Scholar]
- Dordas, C. Role of Nutrients in Controlling Plant Diseases in Sustainable Agriculture: A Review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Gaurat, J.O.; de Barros Silva, E.; Monteiro Cruz, M.d.C.; Esdras Lima, J. Foliar Sampling Time and Critical Level Diagnosis of Nutrients for Blackberry. J. Plant Nutr. 2022, 1–12. [Google Scholar] [CrossRef]
- Azevedo Junior, R.R.; Dos Santos, J.B.; Baretta, D.; Ramos, A.C.; Otto, R.; Façanha, A.R.; Nogueira Cardoso, E.J.B. Discriminating Organic and Conventional Coffee Production Systems through Soil and Foliar Analysis Using Multivariate Approach. Commun. Soil Sci. Plant Anal. 2019, 50, 651–661. [Google Scholar] [CrossRef]
- Shedley, E.; Dell, B.; Grove, T. Diagnosis of Nitrogen Deficiency and Toxicity of Eucalyptus Globulus Seedlings by Foliar Analysis. Plant Soil 1995, 177, 183–189. [Google Scholar] [CrossRef]
- Sanchez-Gonzalez, E.I.; Soares, T.d.P.F.; Zarpelon, T.G.; Zauza, E.A.V.; Mafia, R.G.; Ferreira, M.A. Two New Species of Calonectria (Hypocreales, Nectriaceae) Causing Eucalyptus Leaf Blight in Brazil. Mycokeys 2022, 91, 169–197. [Google Scholar] [CrossRef]
- Bose, R.; Banerjee, S.; Pandey, A.; Bhandari, M.S.; Barthwal, S.; Pandey, S. Calonectria Leaf Blight of Eucalyptus: A Global Review. Ann. Appl. Biol. 2022, 182, 6–28. [Google Scholar] [CrossRef]
- Mohanty, P.S.; Pandey, A.; Arya, P.; Harsh, N.S.K. Molecular Variability in North Indian Isolates of Cylindrocladium Quinquieseptatum Causing Eucalyptus Leaf and Seedling Blight. Indian J. Microbiol. 2012, 52, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Poke, F.S.; Vaillancourt, R.E.; Potts, B.M.; Reid, J.B. Genomic Research in Eucalyptus. Genetica 2005, 125, 79–101. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, H.G.; Jacomine, P.K.T.; dos Anjos, L.H.C.; de Oliveira, V.Á.; Lumbreras, J.F.; Coelho, M.R.; de Almeida, J.A.; Filho, J.C.d.A.; de Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 5th ed.; Embrapa Solos: Brasília, Brazil, 2018. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA: Washington, DC, USA, 2014.
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Avaliação Do Estado Nutricional Das Plantas: Princípios e Aplicações, 2nd ed.; Associação Brasileira para pesquisa da Potassa e do Fosfato: Piracicaba, Brazil, 1997. [Google Scholar]
- Miller, R. Nitric-Perchloric Acid Wet Digestion in an Open Vessel. In Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1998; pp. 57–61. [Google Scholar]
- Weindorf, D.C.; Chakraborty, S. Portable Apparatus for Soil Chemical Characterization. U.S. Patent No. 10107770, 23 October 2018. [Google Scholar]
- Kursa, M.B.; Rudnicki, W.R. Feature Selection with the Boruta Package. J. Stat. Soft. 2010, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environmental for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kuhn, M. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Olsen, L.R. Creating Groups from Data. R Package Version 2.0.1. 2022. Available online: https://cran.r-project.org/web/packages/groupdata2/vignettes/automatic_groups_with_groupdata2.html (accessed on 4 September 2022).
- Chang, C.-W.; Laird, D.A.; Mausbach, M.J.; Hurburgh, C.R. Near-Infrared Reflectance Spectroscopy–Principal Components Regression Analyses of Soil Properties. Soil Sci. Soc. Am. J. 2001, 65, 480. [Google Scholar] [CrossRef] [Green Version]
- Congalton, R.G. A Review of Assessing the Accuracy of Classifications of Remotely Sensed Data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Dataset | Min | Max | Mean | STD 1 | CV (%) 2 |
|---|---|---|---|---|---|---|
| N | Total | 0.00 | 56.82 | 23.85 | 6.19 | 25.95 |
| Field | 0.00 | 37.84 | 22.99 | 5.54 | 24.12 | |
| Nursery | 2.36 | 56.82 | 29.52 | 7.16 | 24.25 | |
| P | Total | 0.62 | 5.14 | 1.64 | 0.70 | 42.74 |
| Field | 0.62 | 3.40 | 1.43 | 0.39 | 27.60 | |
| Nursery | 1.83 | 5.14 | 3.07 | 0.61 | 19.83 | |
| K | Total | 5.54 | 25.60 | 11.56 | 3.67 | 31.77 |
| Field | 5.54 | 24.36 | 10.41 | 2.12 | 20.34 | |
| Nursery | 12.67 | 25.60 | 19.11 | 2.60 | 13.61 | |
| Ca | Total | 2.32 | 29.04 | 8.19 | 3.29 | 40.23 |
| Field | 2.32 | 29.04 | 8.27 | 3.48 | 42.02 | |
| Nursery | 3.52 | 13.18 | 7.63 | 1.57 | 20.52 | |
| Mg | Total | 1.57 | 7.10 | 3.55 | 0.74 | 20.94 |
| Field | 1.57 | 7.10 | 3.58 | 0.77 | 21.45 | |
| Nursery | 2.23 | 4.64 | 3.32 | 0.49 | 14.89 | |
| S | Total | 0.98 | 4.06 | 2.10 | 0.41 | 19.79 |
| Field | 0.98 | 3.30 | 2.01 | 0.34 | 17.07 | |
| Nursery | 1.73 | 4.06 | 2.65 | 0.42 | 15.85 | |
| B | Total | 11.24 | 147.50 | 48.14 | 18.27 | 37.95 |
| Field | 11.24 | 147.50 | 49.48 | 18.87 | 38.14 | |
| Nursery | 21.82 | 79.74 | 39.33 | 9.90 | 25.18 | |
| Mn | Total | 29.74 | 4170.40 | 855.12 | 683.10 | 79.88 |
| Field | 62.22 | 4170.40 | 965.95 | 666.88 | 69.04 | |
| Nursery | 29.74 | 498.00 | 130.19 | 61.93 | 47.57 | |
| Fe | Total | 40.30 | 344.10 | 110.85 | 38.03 | 34.31 |
| Field | 40.30 | 344.10 | 114.39 | 38.22 | 33.42 | |
| Nursery | 53.13 | 224.90 | 87.72 | 27.21 | 31.01 | |
| Cu | Total | 0.07 | 27.41 | 5.65 | 2.71 | 47.96 |
| Field | 0.07 | 17.58 | 5.59 | 2.53 | 45.20 | |
| Nursery | 1.64 | 27.41 | 6.03 | 3.67 | 60.94 | |
| Zn | Total | 8.30 | 89.63 | 21.73 | 8.18 | 37.64 |
| Field | 8.30 | 89.63 | 19.86 | 5.48 | 27.59 | |
| Nursery | 17.17 | 80.20 | 33.91 | 11.73 | 34.58 |
| Statistics | Al | Ba | Br * | Ca | Cl | Cu * | Fe | K | Mg | Mn | Mo * | Ni * | P | Rb * | S | Si | Sr * | Ti * | V * | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | ||||||||||||||||||||
| Min | 0.04 | 0.01 | 0.02 | 0.53 | 0.02 | 0.03 | 0.00 | 0.31 | 0.20 | 0.00 | 0.02 | 0.01 | 0.03 | 0.02 | 0.01 | 0.01 | 0.04 | 0.03 | 0.01 | 0.00 |
| Max | 0.28 | 0.03 | 3.60 | 3.56 | 0.80 | 0.22 | 0.10 | 2.36 | 0.53 | 0.85 | 0.04 | 0.53 | 0.38 | 0.76 | 0.33 | 0.59 | 5.27 | 0.39 | 0.03 | 0.01 |
| Mean | 0.08 | 0.01 | 0.43 | 1.10 | 0.29 | 0.08 | 0.01 | 0.81 | 0.26 | 0.13 | 0.03 | 0.03 | 0.10 | 0.13 | 0.12 | 0.09 | 0.45 | 0.05 | 0.01 | 0.00 |
| STD 1 | 0.03 | 0.00 | 0.42 | 0.33 | 0.13 | 0.02 | 0.01 | 0.32 | 0.05 | 0.12 | 0.01 | 0.03 | 0.04 | 0.09 | 0.03 | 0.06 | 0.37 | 0.03 | 0.00 | 0.00 |
| CV (%) 2 | 32.24 | 32.07 | 98.38 | 29.77 | 45.25 | 28.94 | 54.38 | 38.98 | 20.94 | 86.64 | 21.71 | 96.50 | 35.46 | 70.91 | 23.65 | 61.47 | 81.77 | 50.35 | 24.74 | 41.15 |
| Field | ||||||||||||||||||||
| Min | 0.04 | 0.01 | 0.02 | 0.53 | 0.02 | 0.03 | 0.00 | 0.31 | 0.20 | 0.01 | 0.02 | 0.01 | 0.03 | 0.03 | 0.01 | 0.01 | 0.04 | 0.03 | 0.01 | 0.00 |
| Max | 0.28 | 0.03 | 3.60 | 3.56 | 0.80 | 0.22 | 0.10 | 1.80 | 0.53 | 0.85 | 0.04 | 0.53 | 0.20 | 0.76 | 0.33 | 0.59 | 5.27 | 0.39 | 0.03 | 0.01 |
| Mean | 0.09 | 0.01 | 0.48 | 1.11 | 0.28 | 0.08 | 0.01 | 0.71 | 0.26 | 0.15 | 0.03 | 0.03 | 0.09 | 0.13 | 0.12 | 0.10 | 0.48 | 0.05 | 0.01 | 0.00 |
| STD 1 | 0.03 | 0.00 | 0.43 | 0.34 | 0.13 | 0.02 | 0.01 | 0.19 | 0.06 | 0.11 | 0.01 | 0.03 | 0.02 | 0.09 | 0.03 | 0.06 | 0.38 | 0.03 | 0.00 | 0.00 |
| CV (%) 2 | 32.00 | 32.00 | 91.18 | 30.87 | 46.98 | 28.92 | 54.95 | 25.95 | 21.31 | 75.35 | 23.22 | 96.42 | 23.47 | 69.16 | 23.06 | 56.68 | 79.47 | 50.35 | 24.74 | 27.34 |
| Nursery | ||||||||||||||||||||
| Min | 0.05 | 0.01 | 0.04 | 0.55 | 0.16 | 0.05 | 0.01 | 0.85 | 0.20 | 0.00 | 0.03 | 0.00 | 0.09 | 0.02 | 0.08 | 0.01 | 0.08 | 0.00 | 0.00 | 0.00 |
| Max | 0.11 | 0.01 | 0.28 | 1.65 | 0.74 | 0.21 | 0.03 | 2.36 | 0.33 | 0.08 | 0.04 | 0.00 | 0.38 | 0.06 | 0.28 | 0.29 | 0.63 | 0.00 | 0.00 | 0.01 |
| Mean | 0.07 | 0.01 | 0.12 | 1.01 | 0.39 | 0.08 | 0.01 | 1.45 | 0.24 | 0.02 | 0.03 | 0.00 | 0.17 | 0.04 | 0.15 | 0.04 | 0.24 | 0.00 | 0.00 | 0.00 |
| STD 1 | 0.02 | 0.00 | 0.05 | 0.17 | 0.10 | 0.02 | 0.00 | 0.25 | 0.04 | 0.01 | 0.00 | 0.00 | 0.04 | 0.01 | 0.03 | 0.03 | 0.10 | 0.00 | 0.00 | 0.00 |
| CV (%) 2 | 23.25 | 12.79 | 44.11 | 16.75 | 26.50 | 29.17 | 33.05 | 17.39 | 17.73 | 66.13 | 14.81 | 0.00 | 23.83 | 28.51 | 19.81 | 89.18 | 42.12 | 0.00 | 0.00 | 40.47 |
| Nutrient | Model | RMSE | R2 | RPD | Nutrient | Model | RMSE | R2 | RPD |
|---|---|---|---|---|---|---|---|---|---|
| N | PPR | 4.46 | 0.51 | 1.42 | B | PPR | 15.48 | 0.26 | 1.16 |
| PLS | 4.56 | 0.49 | 1.39 | PLS | 15.63 | 0.24 | 1.15 | ||
| RF | 3.81 | 0.64 | 1.66 | RF | 14.82 | 0.33 | 1.21 | ||
| SVM | 4.49 | 0.51 | 1.41 | SVM | 15.65 | 0.26 | 1.15 | ||
| XGB | 4.28 | 0.55 | 1.48 | XGB | 16.46 | 0.21 | 1.09 | ||
| Cubist | 3.97 | 0.62 | 1.60 | Cubist | 15.47 | 0.26 | 1.16 | ||
| P | PPR | 0.19 | 0.94 | 3.94 | Cu | PPR | 1.59 | 0.63 | 1.62 |
| PLS | 0.19 | 0.93 | 3.85 | PLS | 1.57 | 0.64 | 1.64 | ||
| RF | 0.18 | 0.94 | 4.13 | RF | 1.70 | 0.57 | 1.51 | ||
| SVM | 0.20 | 0.93 | 3.81 | SVM | 1.52 | 0.66 | 1.69 | ||
| XGB | 0.24 | 0.90 | 3.12 | XGB | 1.97 | 0.45 | 1.31 | ||
| Cubist | 0.20 | 0.93 | 3.72 | Cubist | 1.64 | 0.60 | 1.57 | ||
| K | PPR | 0.89 | 0.95 | 4.40 | Fe | PPR | 16.04 | 0.81 | 2.31 |
| PLS | 0.93 | 0.94 | 4.22 | PLS | 17.40 | 0.80 | 2.13 | ||
| RF | 0.92 | 0.95 | 4.25 | RF | 19.39 | 0.78 | 1.91 | ||
| SVM | 0.92 | 0.94 | 4.24 | SVM | 14.58 | 0.85 | 2.54 | ||
| XGB | 0.90 | 0.95 | 4.35 | XGB | 19.21 | 0.74 | 1.93 | ||
| Cubist | 0.86 | 0.95 | 4.59 | Cubist | 15.61 | 0.83 | 2.37 | ||
| Ca | PPR | 1.05 | 0.89 | 3.05 | Mn | PPR | 113.00 | 0.97 | 5.88 |
| PLS | 1.06 | 0.89 | 3.02 | PLS | 132.00 | 0.96 | 5.04 | ||
| RF | 1.01 | 0.90 | 3.17 | RF | 129.34 | 0.96 | 5.14 | ||
| SVM | 1.04 | 0.90 | 3.08 | SVM | 128.51 | 0.96 | 5.17 | ||
| XGB | 1.11 | 0.88 | 2.89 | XGB | 154.38 | 0.95 | 4.31 | ||
| Cubist | 1.03 | 0.90 | 3.10 | Cubist | 123.07 | 0.97 | 5.40 | ||
| Mg | PPR | 0.59 | 0.40 | 1.29 | Zn | PPR | 4.67 | 0.67 | 1.73 |
| PLS | 0.63 | 0.31 | 1.20 | PLS | 4.82 | 0.64 | 1.68 | ||
| RF | 0.55 | 0.49 | 1.39 | RF | 4.22 | 0.75 | 1.92 | ||
| SVM | 0.64 | 0.30 | 1.19 | SVM | 4.84 | 0.65 | 1.67 | ||
| XGB | 0.61 | 0.36 | 1.25 | XGB | 4.27 | 0.72 | 1.90 | ||
| Cubist | 0.55 | 0.49 | 1.40 | Cubist | 4.56 | 0.69 | 1.77 | ||
| S | PPR | 0.18 | 0.81 | 2.27 | |||||
| PLS | 0.19 | 0.79 | 2.17 | ||||||
| RF | 0.17 | 0.85 | 2.52 | ||||||
| SVM | 0.18 | 0.82 | 2.34 | ||||||
| XGB | 0.20 | 0.78 | 2.05 | ||||||
| Cubist | 0.16 | 0.85 | 2.62 |
| Nutrient | Model | OA | CKC | Nutrient | Model | OA | CKC |
|---|---|---|---|---|---|---|---|
| N | C5.0 | 0.67 | 0.50 | B | C5.0 | 0.63 | 0.33 |
| PLS | 0.64 | 0.43 | PLS | 0.57 | 0.20 | ||
| RF | 0.75 | 0.60 | RF | 0.61 | 0.30 | ||
| SVM | 0.70 | 0.53 | SVM | 0.59 | 0.23 | ||
| XGB | 0.72 | 0.56 | XGB | 0.62 | 0.32 | ||
| BC | 0.73 | 0.57 | BC | 0.63 | 0.35 | ||
| P | C5.0 | 0.91 | 0.83 | Cu | C5.0 | 0.80 | 0.59 |
| PLS | 0.79 | 0.53 | PLS | 0.65 | 0.13 | ||
| RF | 0.91 | 0.83 | RF | 0.80 | 0.59 | ||
| SVM | 0.88 | 0.77 | SVM | 0.80 | 0.56 | ||
| XGB | 0.89 | 0.80 | XGB | 0.79 | 0.58 | ||
| BC | 0.89 | 0.81 | BC | 0.76 | 0.54 | ||
| K | C5.0 | 0.80 | 0.64 | Fe | C5.0 | 0.87 | 0.72 |
| PLS | 0.68 | 0.38 | PLS | 0.85 | 0.64 | ||
| RF | 0.80 | 0.65 | RF | 0.88 | 0.73 | ||
| SVM | 0.80 | 0.64 | SVM | 0.88 | 0.71 | ||
| XGB | 0.81 | 0.67 | XGB | 0.87 | 0.69 | ||
| BC | 0.79 | 0.63 | BC | 0.87 | 0.72 | ||
| Ca | C5.0 | 0.80 | 0.68 | Mn | C5.0 | 0.94 | 0.90 |
| PLS | 0.73 | 0.58 | PLS | 0.88 | 0.80 | ||
| RF | 0.83 | 0.73 | RF | 0.94 | 0.91 | ||
| SVM | 0.83 | 0.73 | SVM | 0.95 | 0.92 | ||
| XGB | 0.80 | 0.68 | XGB | 0.96 | 0.93 | ||
| BC | 0.79 | 0.66 | BC | 0.94 | 0.91 | ||
| Mg | C5.0 | 0.62 | 0.32 | Zn | C5.0 | 0.69 | 0.49 |
| PLS | 0.67 | 0.38 | PLS | 0.67 | 0.41 | ||
| RF | 0.70 | 0.45 | RF | 0.72 | 0.52 | ||
| SVM | 0.65 | 0.35 | SVM | 0.75 | 0.58 | ||
| XGB | 0.68 | 0.40 | XGB | 0.68 | 0.48 | ||
| BC | 0.65 | 0.38 | BC | 0.71 | 0.51 | ||
| S | C5.0 | 0.93 | 0.75 | ||||
| PLS | 0.85 | 0.34 | |||||
| RF | 0.91 | 0.67 | |||||
| SVM | 0.91 | 0.70 | |||||
| XGB | 0.92 | 0.69 | |||||
| BC | 0.91 | 0.68 |
| Identification of Figure 6 | SiBCS | Soil Taxonomy |
|---|---|---|
| C | Cambisol | Inceptisol |
| F | Plinthosol | Oxisol |
| G | Gleysol | Entisol (Aquent) |
| M | Chernosol | Mollisol |
| N | Nitosol | Ultisol |
| P | Argisol | Ultisol |
| R | Neosol | Entisol |
| S | Planosol | Alfisol |
| T | Luvisol | Alfisol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, R.; Silva, S.H.G.; Benedet, L.; de Araújo, E.F.; Carneiro, M.A.C.; Curi, N. A Proximal Sensor-Based Approach for Clean, Fast, and Accurate Assessment of the Eucalyptus spp. Nutritional Status and Differentiation of Clones. Plants 2023, 12, 561. https://doi.org/10.3390/plants12030561
Andrade R, Silva SHG, Benedet L, de Araújo EF, Carneiro MAC, Curi N. A Proximal Sensor-Based Approach for Clean, Fast, and Accurate Assessment of the Eucalyptus spp. Nutritional Status and Differentiation of Clones. Plants. 2023; 12(3):561. https://doi.org/10.3390/plants12030561
Chicago/Turabian StyleAndrade, Renata, Sérgio Henrique Godinho Silva, Lucas Benedet, Elias Frank de Araújo, Marco Aurélio Carbone Carneiro, and Nilton Curi. 2023. "A Proximal Sensor-Based Approach for Clean, Fast, and Accurate Assessment of the Eucalyptus spp. Nutritional Status and Differentiation of Clones" Plants 12, no. 3: 561. https://doi.org/10.3390/plants12030561