
Inhibitory Activity of Saussurea costus Extract against Bacteria, Candida, Herpes, and SARS-CoV-2

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Analysis of the Organic Chemical Compounds in Saussurea costus
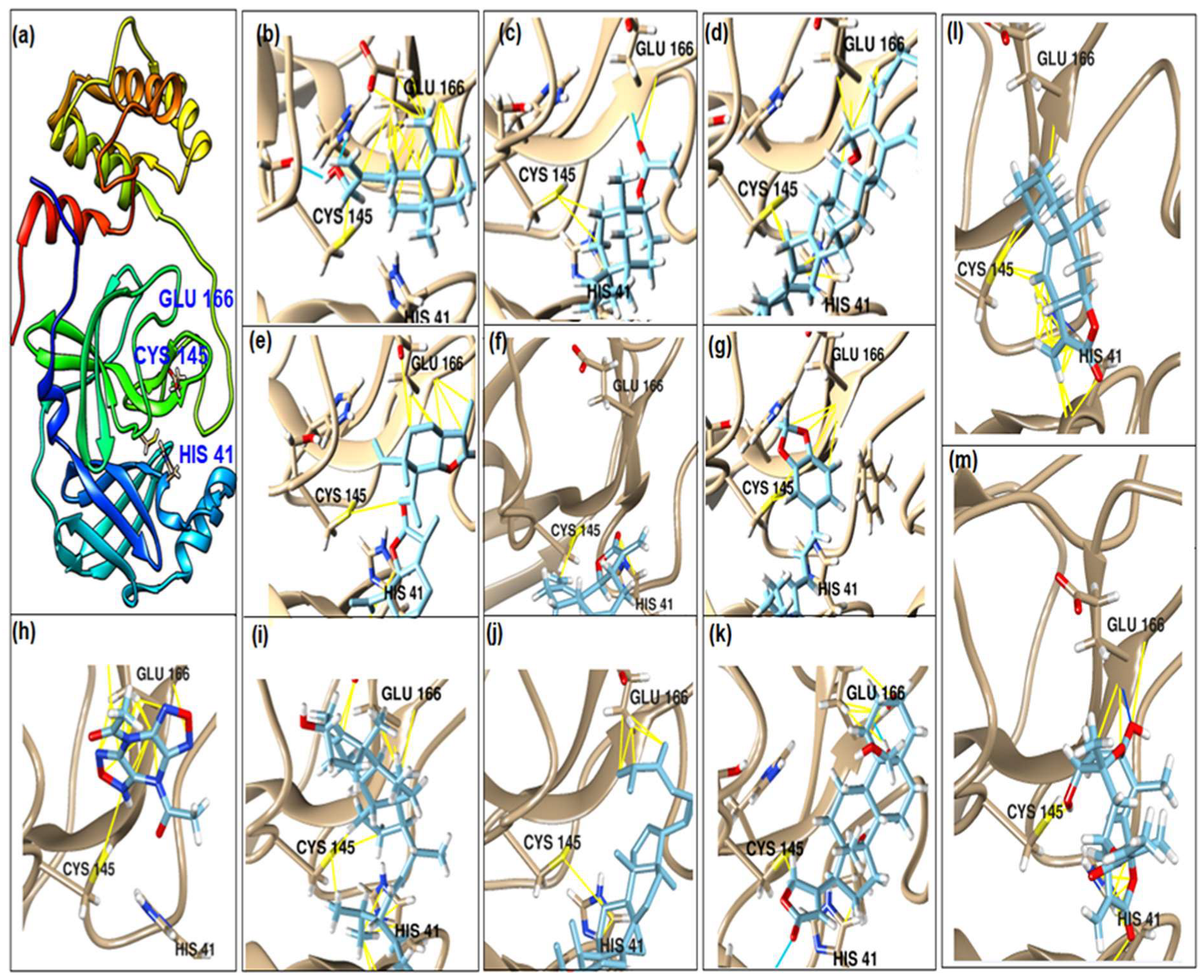
2.2. Molecular Docking
2.3. Antimicrobial Activity
2.4. Antiviral Activity by Saussurea costus Acetic Extract
3. Materials and Methods
3.1. Chemicals
3.2. Sample Preparation and Extraction
3.3. Gas Chromatography–Mass Spectrometry (GC/MS) Analysis
3.4. Molecular Docking
3.5. Microorganisms
3.6. Disc-Diffusion Test
3.7. Minimum Inhibitory Concentration MIC Assay
3.8. Minimum Bactericidal Concentration or Minimum Fungicidal Concentration MBC or MFC Assay
3.9. Virological Assessments
3.9.1. Cytotoxicity Assays
3.9.2. HIV-1-Based SARS-CoV-2 Pseudotyped Particles
3.9.3. Inhibition of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Entry Assay
3.9.4. Antiviral Activity against Herpes Simplex Virus Type-1 (HSV-1)
3.9.5. Time-of-Addition Assay of HSV-1
3.10. Statistical Analysis
3.11. Research Limitations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdallah, E.M. Plants: An alternative source for antimicrobials. J. Appl. Pharm. Sci. 2011, 1, 16–20. [Google Scholar]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi, B.; Abbaszadeh, S.; Shahsavari, S.; Alizadeh, M.; Beyranvand, F. The most useful medicinal herbs to treat diabetes. Biomed. Res. Ther. 2018, 5, 2538–2551. [Google Scholar] [CrossRef]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Shukla, S.; Mehta, A. Anticancer potential of medicinal plants and their phytochemicals: A review. Braz. J. Bot. 2015, 38, 199–210. [Google Scholar] [CrossRef]
- Vashist, H.; Jindal, A. Antimicrobial activities of medicinal plants–Review. Int. J. Res. Pharm. Biomed. Sci. 2012, 3, 222–230. [Google Scholar]
- Umeta Chali, B.; Melaku, T.; Berhanu, N.; Mengistu, B.; Milkessa, G.; Mamo, G.; Alemu, S.; Mulugeta, T. Traditional medicine practice in the context of COVID-19 pandemic: Community claim in jimma zone, oromia, Ethiopia. Infect Drug Resist. 2021, 14, 3773–3783. [Google Scholar] [CrossRef]
- Gwari, G.; Bhandari, U.; Andola, H.C.; Lohani, H.; Chauhan, N. Volatile constituents of Saussurea costus roots cultivated in Uttarakhand Himalayas, India. Pharmacogn. Res. 2013, 5, 179–182. [Google Scholar]
- Ansari, S.; Siddiqui, M.; Malhotra, S.; Maaz, M. Antiviral Efficacy of Qust (Saussurea lappa) and Afsanteen (Artemisia absinthium) for Chronic Hepatitis B: A Prospective Single-Arm Pilot Clinical Trial. Pharmacogn. Res. 2018, 10, 282. [Google Scholar] [CrossRef]
- Hassan, R.; Masoodi, M.H. Saussurea lappa: A comprehensive review on its pharmacological activity and phytochemistry. Curr. Tradit. Med. 2020, 6, 13–23. [Google Scholar] [CrossRef]
- WHO. WHO COVID-19 Preparedness and Response Progress Report-1 February to 30 June 2020; WHO: Geneva, Switzerland, 2020.
- Gao, Y.; Cai, G.-Y.; Fang, W.; Li, H.-Y.; Wang, S.-Y.; Chen, L.; Yu, Y.; Liu, D.; Xu, S.; Cui, P.-F.; et al. Machine learning based early warning system enables accurate mortality risk prediction for COVID-19. Nat. Commun. 2020, 11, 5033. [Google Scholar] [CrossRef] [PubMed]
- Nickbakhsh, S.; Mair, C.; Matthews, L.; Reeve, R.; Johnson, P.C.D.; Thorburn, F.; von Wissmann, B.; Reynolds, A.; McMenamin, J.; Gunson, R.N.; et al. Virus–virus interactions impact the population dynamics of influenza and the common cold. Proc. Natl. Acad. Sci. 2019, 116, 27142–27150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatkin, D.; Macrae, C.; Sasaki, T.; Wolff, M.R.; Porcu, M.; Frenneaux, M.; Atherton, J.; Vidaillet, H.J., Jr.; Spudich, S.; De Girolami, U.; et al. Missense mutations in the rod domain of the lamin A/C gene as causes of dilated cardiomyopathy and conduction-system disease. New Engl. J. Med. 1999, 341, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, Z.; Chen, W.; Cai, Z.; Ge, X.; Zhu, H.; Jiang, T.; Tan, W.; Peng, Y. Predicting the receptor-binding domain usage of the coronavirus based on kmer frequency on spike protein. Infect. Genet. Evol. 2018, 61, 183. [Google Scholar] [CrossRef]
- Hutchison, C.J.; Alvarez-Reyes, M.; Vaughan, O.A. Lamins in disease: Why do ubiquitously expressed nuclear envelope proteins give rise to tissue-specific disease phenotypes? J. Cell Sci. 2001, 114, 9–19. [Google Scholar] [CrossRef]
- Zhang, X.; Kousoulas, K.G.; Storz, J. The hemagglutinin/esterase glycoprotein of bovine coronaviruses: Sequence and functional comparisons between virulent and avirulent strains. Virology 1991, 185, 847–852. [Google Scholar] [CrossRef]
- Salazar, E.; Christensen, P.A.; Graviss, E.A.; Nguyen, D.T.; Castillo, B.; Chen, J.; Lopez, B.V.; Eagar, T.N.; Yi, X.; Zhao, P.; et al. Treatment of coronavirus disease 2019 patients with convalescent plasma reveals a signal of significantly decreased mortality. Am. J. Pathol. 2020, 190, 2290–2303. [Google Scholar] [CrossRef]
- Holmes, E.A.; O’Connor, R.C.; Perry, V.H.; Tracey, I.; Wessely, S.; Arseneault, L.; Ballard, C.; Christensen, H.; Silver, R.C.; Everall, I.; et al. Multidisciplinary research priorities for the COVID-19 pandemic: A call for action for mental health science. Lancet Psychiatry 2020, 7, 547–560. [Google Scholar] [CrossRef]
- Mintah, S.O.; Asafo-Agyei, T.; Archer, M.A.; Junior, P.A.A.; Boamah, D.; Kumadoh, D.; Appiah, A.; Ocloo, A.; Boakye, Y.D.; Agyare, C. Medicinal plants for treatment of prevalent diseases. In Pharmacognosy-Medicinal Plants; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar]
- Tahir, A.H.; Javed, M.M.; Hussain, Z. Nutraceuticals and herbal extracts: A ray of hope for COVID-19 and related infections. Int. J. Funct. Nutr. 2020, 1, 6. [Google Scholar] [CrossRef]
- Glatthaar-Saalmüller, B.; Sacher, F.; Esperester, A. Antiviral activity of an extract derived from roots of Eleutherococcus senticosus. Antivir. Res. 2001, 50, 223–228. [Google Scholar] [CrossRef]
- Karagöz, A.; Arda, N.; Gören, N.; Nagata, K.; Kuru, A. Antiviral activity of Sanicula europaea L. extracts on multiplication of human parainfluenza virus type 2. Phytother. Res. 1999, 13, 436–438. [Google Scholar] [PubMed]
- Al-Mijalli, S.H.; Mrabti, N.N.; Ouassou, H.; Sheikh, R.; Abdallah, E.; Assaggaf, H.; Bakrim, S.; Alshahrani, M.M.; Al Awadh, A.A.; Qasem, A.; et al. Phytochemical Variability, In Vitro and In Vivo Biological Investigations, and In Silico Antibacterial Mechanisms of Mentha piperita Essential Oils Collected from Two Different Regions in Morocco. Foods 2022, 11, 3466. [Google Scholar] [CrossRef]
- Razzaque, M.S. Commentary: Microbial resistance movements: An overview of global public health threats posed by antimicrobial resistance, and how best to counter. Front. Public Health 2021, 8, 629120. [Google Scholar] [CrossRef] [PubMed]
- Jassim, S.A.; Naji, M.A. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harazem, R.; El Rahman, S.A.; El-Kenawy, A. Evaluation of Antiviral Activity of Allium Cepa and Allium Sativum Extracts Against Newcastle Disease Virus. Alex. J. Vet. Sci. 2019, 61, 108. [Google Scholar] [CrossRef]
- Badam, L.; Joshi, S.; Bedekar, S. ‘In vitro’ antiviral activity of neem (Azadirachta indica A. Juss) leaf extract against group B coxsackieviruses. J. Commun. Dis. 1999, 31, 79–90. [Google Scholar]
- Sharma, P.; Dwivedee, B.P.; Bisht, D.; Dash, A.K.; Kumar, D. The chemical constituents and diverse pharmacological importance of Tinospora cordifolia. Heliyon 2019, 5, e02437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayan, P.; Raghu, C.; Ashok, G.; A Dhanaraj, S.; Suresh, B. Antiviral activity of medicinal plants of Nilgiris. Indian J. Med. Res. 2004, 120, 24–29. [Google Scholar]
- Kochan, E.; Wasiela, M.; Sienkiewicz, M. The production of ginsenosides in hairy root cultures of American Ginseng, Panax quinquefolium L. and their antimicrobial activity. Vitr. Cell. Dev. Biol. Plant 2013, 49, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Maideen, N.M.P. Prophetic medicine-Nigella Sativa (Black cumin seeds)–potential herb for COVID-19? J. Pharmacopunct. 2020, 23, 62. [Google Scholar] [CrossRef]
- Kim, H.J.; Yoo, H.S.; Kim, J.C.; Park, C.S.; Choi, M.S.; Kim, M.; Choi, H.; Min, J.S.; Kim, Y.S.; Yoon, S.W.; et al. Antiviral effect of Curcuma longa Linn extract against hepatitis B virus replication. J. Ethnopharmacol. 2009, 124, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Torabian, G.; Valtchev, P.; Adil, Q.; Dehghani, F. Anti-influenza activity of elderberry (Sambucus nigra). J. Funct. Foods 2019, 54, 353–360. [Google Scholar] [CrossRef]
- Kumar, D.V.; Geethanjali, B.; Avinash, K.O.; Kumar, J.R.; Basalingappa, K.M. Tinospora cordifolia: The antimicrobial property of the leaves of amruthaballi. J. Bacteriol. Mycol. Open Access 2017, 5, 363–371. [Google Scholar]
- Saha, S.; Ghosh, S. Tinospora cordifolia: One plant, many roles. Anc. Sci. Life 2012, 31, 151. [Google Scholar]
- Panday, J.; Singh, L.; Saxena, G.; Devkota, H.P. Environmental Challenges for Himalayan Medicinal Plants. In Environmental Challenges and Medicinal Plants; Aftab, T., Ed.; Springer: Cham, Switzerland; pp. 29–47.
- Pandey, M.M.; Rastogi, S.; Rawat, A. Saussurea costus: Botanical, chemical and pharmacological review of an ayurvedic medicinal plant. J. Ethnopharmacol. 2007, 110, 379–390. [Google Scholar] [CrossRef]
- Rathore, S.; Debnath, P.; Kumar, R. Kuth Saussurea costus (Falc.) Lipsch.: A critically endangered medicinal plant from Himalaya. J. Appl. Res. Med. Aromat. Plants 2021, 20, 100277. [Google Scholar]
- Singh, D.K.; Tóth, R.; Gácser, A. Mechanisms of pathogenic Candida species to evade the host complement attack. Front. Cell. Infect. Microbiol. 2020, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Simón-Manso, Y.; Lowenthal, M.S.; Kilpatrick, L.E.; Sampson, M.L.; Telu, K.H.; Rudnick, P.A.; Mallard, W.G.; Bearden, D.W.; Schock, T.B.; Tchekhovskoi, D.V.; et al. Metabolite profiling of a NIST Standard Reference Material for human plasma (SRM 1950): GC-MS, LC-MS, NMR, and clinical laboratory analyses, libraries, and web-based resources. Anal. Chem. 2013, 85, 11725–11731. [Google Scholar] [CrossRef]
- Khan, A.; Shah, A.H.; Ali, N. In-vitro propagation and phytochemical profiling of a highly medicinal and endemic plant species of the Himalayan region (Saussurea costus). Sci. Rep. 2021, 11, 23575. [Google Scholar] [CrossRef]
- Kumar, J.; Pundir, M. Phytochemistry and pharmacology of Saussurea genus (Saussurea lappa, Saussurea costus, Saussurea obvallata, Saussurea involucrata). Mater. Today Proc. 2022, 56, 1173–1181. [Google Scholar] [CrossRef]
- Zahnit, W.; Smara, O.; Bechki, L.; Bensouici, C.; Messaoudi, M.; Benchikha, N.; Larkem, I.; Awuchi, C.G.; Sawicka, B.; Simal-Gandara, J. Phytochemical Profiling, Mineral Elements, and Biological Activities of Artemisia campestris L. Grown in Algeria. Horticulturae 2022, 8, 914. [Google Scholar] [CrossRef]
- Mammate, N.; El oumari, F.E.; Imtara, H.; Belchkar, S.; Lahrichi, A.; Alqahtani, A.S.; Noman, O.M.; Tarayrah, M.; Houssaini, T.S. Antioxidant and Anti-Urolithiatic Activity of Aqueous and Ethanolic Extracts from Saussurea costus (Falc) Lispich Using Scanning Electron Microscopy. Life 2022, 12, 1026. [Google Scholar] [CrossRef]
- Ali, S.I.; Venkatesalu, V. Botany, traditional uses, phytochemistry and pharmacological properties of Saussurea costus–An endangered plant from Himalaya-A review. Phytochem. Lett. 2022, 47, 140–155. [Google Scholar] [CrossRef]
- Nadda, R.K.; Ali, A.; Goyal, R.C.; Khosla, P.K.; Goyal, R. Aucklandia costus (Syn. Saussurea costus): Ethnopharmacology of an endangered medicinal plant of the himalayan region. J. Ethnopharmacol. 2020, 263, 113199. [Google Scholar] [CrossRef] [PubMed]
- Madhuri, K.; Elango, K.; Ponnusankar, S. Saussurea lappa (Kuth root): Review of its traditional uses, phytochemistry and pharmacology. Orient. Pharm. Exp. Med. 2012, 12, 1–9. [Google Scholar] [CrossRef]
- Savadi, S.; Vazifedoost, M.; Didar, Z.; Nematshahi, M.M.; Jahed, E. Phytochemical analysis and antimicrobial/antioxidant activity of Cynodon dactylon (L.) Pers. rhizome methanolic extract. J. Food Qual. 2020, 2020, 5946541. [Google Scholar] [CrossRef] [Green Version]
- Dolmatova, L.S.; Dolmatov, I.Y. Tumor-Associated Macrophages as Potential Targets for Anti-Cancer Activity of Marine Invertebrate-Derived Compounds. Curr. Pharm. Des. 2021, 27, 3139–3160. [Google Scholar] [CrossRef]
- Quimque, M.T.J.; Notarte, K.I.R.; de Leon, V.N.O.; Manzano, J.A.H.; Munoz, J.E.R.; Pilapil IV, D.Y.H.; Lim, J.A.K.; Macabeo, A.P.G. Computationally Repurposed Natural Products Targeting SARS-CoV-2 Attachment and Entry Mechanisms. In Frontiers of COVID-19; Adibi, S., Griffin, P., Sanicas, M., Rashidi, M., Lanfranchi, F., Eds.; Springer: Cham, Switzerland, 2022; pp. 505–537. [Google Scholar]
- Khan, J.; Sakib, S.A.; Mahmud, S.; Khan, Z.; Islam, M.N.; Sakib, M.A.; Emran, T.B.; Simal-Gandara, J. Identification of potential phytochemicals from Citrus Limon against main protease of SARS-CoV-2: Molecular docking, molecular dynamic simulations and quantum computations. J. Biomol. Struct. Dyn. 2021, 40, 10741–10752. [Google Scholar] [CrossRef]
- Mej, J.; Zhou, Y.; Yang, X.; Zhang, F.; Liu, X.; Yu, B. Active components in Ephedra sinica stapf disrupt the interaction between ACE2 and SARS-CoV-2 RBD: Potent COVID-19 therapeutic agents. J. Ethnopharmacol. 2021, 278, 114303. [Google Scholar]
- Idriss, H.; Siddig, B.; Maldonado, P.G.; Elkhair, H.M.; Alakhras, A.I.; Abdallah, E.M.; Torres, P.H.S.; Elzupir, A.O. Phytochemical Discrimination, Biological Activity and Molecular Docking of Water-Soluble Inhibitors from Saussurea costus Herb against Main Protease of SARS-CoV-2. Molecules 2022, 27, 4908. [Google Scholar] [CrossRef]
- Romero-Martínez, B.S.; Montaño, L.M.; Solís-Chagoyán, H.; Sommer, B.; Ramírez-Salinas, G.L.; Pérez-Figueroa, G.E.; Flores-Soto, E. Possible beneficial actions of caffeine in SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 5460. [Google Scholar] [CrossRef] [PubMed]
- Rolta, R.; Salaria, D.; Sharma, B.; Awofisayo, O.; Fadare, O.; Sharma, S.; Patel, C.; Kumar, V.; Sourirajan, A.; Baumler, D.; et al. Methylxanthines as Potential Inhibitor of SARS-CoV-2: An In Silico Approach. Curr. Pharmacol. Rep. 2022, 8, 149–170. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; El-Halawany, A.; Sirwi, A.; El-Araby, A.; Mohamed, G.; Ibrahim, S.; Koshak, A.; Asfour, H.; Awan, Z.; Elfaky, M.A. Repurposing of some natural product isolates as SARS-COV-2 main protease inhibitors via in vitro cell free and cell-based antiviral assessments and molecular modeling approaches. Pharmaceuticals 2021, 14, 213. [Google Scholar] [CrossRef]
- van de Sand, L.; Bormann, M.; Alt, M.; Schipper, L.; Heilingloh, C.S.; Steinmann, E.; Todt, D.; Dittmer, U.; Elsner, C.; Witzke, O.; et al. Glycyrrhizin effectively inhibits SARS-CoV-2 replication by inhibiting the viral main protease. Viruses 2021, 13, 609. [Google Scholar] [CrossRef] [PubMed]
- Lockbaum, G.J.; Reyes, A.; Lee, J.M.; Tilvawala, R.; Nalivaika, E.; Ali, A.; Yilmaz, N.K.; Thompson, P.; Schiffer, C. Crystal structure of SARS-CoV-2 main protease in complex with the non-covalent inhibitor ML188. Viruses 2021, 13, 174. [Google Scholar] [CrossRef]
- Elzupir, A.O. Inhibition of SARS-CoV-2 main protease 3CLpro by means of α-ketoamide and pyridone-containing pharmaceuticals using in silico molecular docking. J. Mol. Struct. 2020, 1222, 128878. [Google Scholar] [CrossRef]
- Elzupir, A.O. Molecular Docking and Dynamics Investigations for Identifying Potential Inhibitors of the 3-Chymotrypsin-like Protease of SARS-CoV-2: Repurposing of Approved Pyrimidonic Pharmaceuticals for COVID-19 Treatment. Molecules 2021, 26, 7458. [Google Scholar] [CrossRef]
- Khan, A.; Heng, W.; Wang, Y.; Qiu, J.; Wei, X.; Peng, S.; Saleem, S.; Khan, M.; Ali, S.S.; Wei, D.-Q. In silico and in vitro evaluation of kaempferol as a potential inhibitor of the SARS-CoV-2 main protease (3CLpro). Phytother. Res. 2021, 35, 2841–2845. [Google Scholar] [CrossRef]
- Quimque, M.T.J.; Notarte, K.I.; Fernandez, R.A.; Mendoza, M.A.; Liman, R.A.; Lim, J.A.; Pilapil, L.A.; Ong, J.K.; Pastrana, A.; Khan, A.; et al. Virtual screening-driven drug discovery of SARS-CoV2 enzyme inhibitors targeting viral attachment, replication, post-translational modification and host immunity evasion infection mechanisms. J. Biomol. Struct. Dyn. 2021, 39, 4316–4333. [Google Scholar] [CrossRef]
- Khan, M.T.; Ali, A.; Wang, Q.; Irfan, M.; Khan, A.; Zeb, M.T.; Zhang, Y.-J.; Chinnasamy, S.; Wei, D.-Q. Marine natural compounds as potents inhibitors against the main protease of SARS-CoV-2—A molecular dynamic study. J. Biomol. Struct. Dyn. 2021, 39, 3627–3637. [Google Scholar] [CrossRef]
- Koulgi, S.; Jani, V.; Uppuladinne, M.; Sonavane, U.; Nath, A.K.; Darbari, H.; Joshi, R. Drug repurposing studies targeting SARS-CoV-2: An ensemble docking approach on drug target 3C-like protease (3CLpro). J. Biomol. Struct. Dyn. 2021, 39, 5735–5755. [Google Scholar] [CrossRef] [PubMed]
- Gammeltoft, K.A.; Zhou, Y.; Hernandez, C.D.; Galli, A.; Offersgaard, A.; Costa, R.; Pham, L.; Fahnøe, U.; Feng, S.; Scheel, T.; et al. Hepatitis C virus protease inhibitors show differential efficacy and interactions with remdesivir for treatment of SARS-CoV-2 in vitro. Antimicrob. Agents Chemother. 2021, 65, e02680–20. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.I.; Darwish, K.; Soltane, R.; Chrouda, A.; Mostafa, A.; Shama, N.A.; Elhady, S.; Abulkhair, H.; Khodir, A.; Elmaaty, A.A.; et al. β-Blockers bearing hydroxyethylamine and hydroxyethylene as potential SARS-CoV-2 Mpro inhibitors: Rational based design, in silico, in vitro, and SAR studies for lead optimization. RSC Adv. 2021, 11, 35536–35558. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Yeh, C.-T.; Hsu, C.-W.; Lin, Y.-H. Reduction of ACE2 Serum Concentrations by Telbivudine in Chronic Hepatitis B Patients. Curr. Mol. Med. 2022, 23, 420–424. [Google Scholar] [CrossRef]
- Hussein, R.K.; Khouqeer, G.; Alkaoud, A.M.; El-Khayatt, A.M. Probing the Action of Screened Anticancer Triazole–Tetrazole Derivatives Against COVID-19 Using Molecular Docking and DFT Investigations. Nat. Prod. Commun. 2022, 17, 1934578X221093915. [Google Scholar] [CrossRef]
- Baig, A.; Srinivasan, H. SARS-CoV-2 Inhibitors from Nigella Sativa. Appl. Biochem. Biotechnol. 2022, 194, 1051–1090. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.; Bhardwaj, V.K.; Chauhan, M.; Chauhan, V.; Kumar, A.; Purohit, R.; Kumar, A.; Kumar, S. A ricin-based peptide BRIP from Hordeum vulgare inhibits Mpro of SARS-CoV-2. Sci. Rep. 2022, 12, 12802. [Google Scholar] [CrossRef] [PubMed]
- Mercorelli, B.; Desantis, J.; Celegato, M.; Bazzacco, A.; Siragusa, L.; Benedetti, P.; Eleuteri, M.; Croci, F.; Cruciani, G.; Goracci, L.; et al. Discovery of novel SARS-CoV-2 inhibitors targeting the main protease Mpro by virtual screenings and hit optimization. Antivir. Res. 2022, 204, 105350. [Google Scholar] [CrossRef]
- Silva, S.C.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida species biofilms’ antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef]
- Abdellatif, A.A.H.; Alhathloul, S.S.; Aljohani, A.S.M.; Maswadeh, H.; Abdallah, E.M.; Musa, K.H.; El Hamd, M.A. Green Synthesis of Silver Nanoparticles Incorporated Aromatherapies Utilized for Their Antioxidant and Antimicrobial Activities against Some Clinical Bacterial Isolates. Bioinorg. Chem. Appl. 2022, 2022, 1–14. [Google Scholar] [CrossRef]
- Abdallah, E.; Qureshi, K.; Ali, A.M.H.; Elhassan, G.O. Evaluation of some biological properties of Saussurea costus crude root extract. Biosci. Biotechnol. Res. Commun. 2017, 10, 601–611. [Google Scholar] [CrossRef]
- Siddiqui, Z.N.; Farooq, F.; Musthafa, T.M.; Ahmad, A.; Khan, A.U. Synthesis, characterization and antimicrobial evaluation of novel halopyrazole derivatives. J. Saudi Chem. Soc. 2013, 17, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, J.A.; Sobel, J.D. Candidiasis. In Essentials of clinical mycology; Kauffman, C., Pappas, P., Sobel, J., Dismukes, W., Eds.; Springer: New York, NY, USA, 2011; pp. 167–206. [Google Scholar]
- Canillac, N.; Mourey, A. Antibacterial activity of the essential oil of Picea excelsa on Listeria, Staphylococcus aureus and coliform bacteria. Food Microbiol. 2001, 18, 261–268. [Google Scholar] [CrossRef]
- Hazen, K.C. Fungicidal versus fungistatic activity of terbinafine and itraconazole: An in vitro comparison. J. Am. Acad. Dermatol. 1998, 38, 37–41. [Google Scholar] [CrossRef] [PubMed]
- González-Maldonado, P.; Alvarenga, N.; Burgos-Edwards, A.; Flores-Giubi, M.E.; Barúa, J.; Romero-Rodríguez, M.C.; Soto-Rifo, R.; Valiente-Echeverría, F.; Langjahr, P.; Cantero-González, G.; et al. Screening of Natural Products Inhibitors of SARS-CoV-2 Entry. Molecules 2022, 27, 1743. [Google Scholar] [CrossRef] [PubMed]
- Al-Janabi, A.S.; Elzupir, A.O.; Yousef, T.A. Synthesis, anti-bacterial evaluation, DFT study and molecular docking as a potential 3-chymotrypsin-like protease (3CLpro) of SARS-CoV-2 inhibitors of a novel Schiff bases. J. Mol. Struct. 2021, 1228, 129454. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Al-Mijalli, S.H.; Assaggaf, H.; Qasem, A.; El-Shemi, A.G.; Abdallah, E.M.; Mrabti, H.N.; Bouyahya, A. Antioxidant, Antidiabetic, and Antibacterial Potentials and Chemical Composition of Salvia officinalis and Mentha suaveolens Grown Wild in Morocco. Adv. Pharmacol. Pharm. Sci. 2022, 2022, 1–10. [Google Scholar] [CrossRef]
- Ansel, H.C.; Norred, W.P.; Roth, I.L. Antimicrobial activity of dimethyl sulfoxide against Escherichia coli, Pseudomonas aeruginosa, and Bacillus megaterium. J. Pharm. Sci. 1969, 58, 836–839. [Google Scholar] [CrossRef]
- Hassan, A.S. The antibacterial activity of dimethyl sulfoxide (DMSO) with and without of some ligand complexes of the transitional metal ions of ethyl coumarin against bacteria isolate from burn and wound infection. J. Nat. Sci. Res. 2014, 4, 426–435. [Google Scholar]
- Czuprynski, C.J.; Henson, P.M.; Campbell, P.A. Effect of dimethyl sulfoxide on the in vitro and in vivo bactericidal activity of human and mouse neutrophils and mononuclear phagocytes. Inflammation 1984, 8, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Gabaglio, S.; Alvarenga, N.; Cantero-González, G.; Degen, R.; Ferro, E.A.; Langjahr, P.; Chnaiderman, J.; Sotelo, P.H. A quantitative PCR assay for antiviral activity screening of medicinal plants against Herpes simplex 1. Nat. Prod. Res. 2021, 35, 2926–2930. [Google Scholar] [CrossRef] [PubMed]
- Gulluce, M.; Aslan, A.; Sokmen, M.; Sahin, F.; Adiguzel, A.; Agar, G.; Sokmen, A. Screening the antioxidant and antimicrobial properties of the lichens Parmelia saxatilis, Platismatia glauca, Ramalina pollinaria, Ramalina polymorpha and Umbilicaria nylanderiana. Phytomedicine 2006, 13, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Gonzales-Wartz, K.K.; Huang, D.; Yuan, M.; Peterson, M.; Liang, J.; Beutler, N.; Torres, J.; Cong, Y.; Postnikova, E.; et al. Bispecific antibodies targeting distinct regions of the spike protein potently neutralize SARS-CoV-2 variants of concern. Sci. Transl. Med. 2021, 13, eabj5413. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Huang, D.; Lee, C.-C.; Wu, N.; Jackson, A.; Zhu, X.; Liu, H.; Peng, L.; van Gils, M.; Sanders, R.; et al. Structural and functional ramifications of antigenic drift in recent SARS-CoV-2 variants. Science 2021, 373, 818–823. [Google Scholar] [CrossRef]
- Kingston, R.E.; Chen, C.A.; Rose, J.K. Calcium phosphate transfection. Curr. Protoc. Mol. Biol. 2003, 63, 9. [Google Scholar] [CrossRef]
- Telu, K.H.; Yan, X.; Wallace, W.E.; Stein, S.E.; Simón-Manso, Y. Analysis of human plasma metabolites across different liquid chromatography/mass spectrometry platforms: Cross-platform transferable chemical signatures. Rapid Commun. Mass Spectrom. 2016, 30, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Alam, A.; Husain, S.; Ahmed, S.; Nazamuddin, M.; Ahmed, Z. A potent herb of Unani medicine: A review. Int. J. Curr. Pharm. Res. 2013, 5, 1–4. [Google Scholar]
- Tag, H.M.; Khaled, H.E.; Ismail, H.A.; El-Shenawy, N.S. Evaluation of anti-inflammatory potential of the ethanolic extract of the Saussurea lappa root (costus) on adjuvant-induced monoarthritis in rats. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 71–78. [Google Scholar] [CrossRef]
- Okubo, S.; Ohta, T.; Fujita, H.; Shoyama, Y.; Uto, T. Costunolide and dehydrocostuslactone from Saussurea lappa root inhibit autophagy in hepatocellular carcinoma cells. J. Nat. Med. 2021, 75, 240–245. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Chemical Structure | Score (kcal/mol) | Root-Mean-Square Deviation of Atomic Positions RMSD Range | Hydrogen Bonds (Number of Bonds/Number of Conformations), (Distance ≤ 4 Å) | Van der Waals (Number of Bonds/Number of Conformations), (Distance Range Å) |
|---|---|---|---|---|---|---|
| b | 2-(4a,8-Dimethyl-1,2,3,4,4a,5,6,7-octahydro-naphthalen-2-yl)-prop-2-en-1-ol |  | −5.6 | 27.83–29.77 | HIS 163 (1/1), SER 144 (1/1) | GLU 166 (19/1), (2.52–3.87) CYS 145 (1/1), (4.01) |
| d | 9beta-Acetoxy-3,5alpha,8-trimethyltricyclo[6.3.1.0(1,5)]dodec-3-ene |  | −5.6 | 33.48–36.10 | GLU 166 (1/1) | GLU 166 (5/2), (2.01–3.80) HIS 41 (6/2), (3.11–3.80) CYS 145 (4/2), (3.76–3.95) |
| c | 3-Oxatricyclo[20.8.0.0(7,16)]triaconta-1(22),7(16),9,13,23,29-hexaene |  | −7.8 | 27.51–30.19 | GLU 166 (1/1) | GLU 166 (6/1), (2.66–3.70) HIS 41 (2/1), (3.74–3.76) CYS 145 (2/1), (3.51 to 3.69) |
| d | 2(3H)-Benzofuranone, 6-ethenylhexahydro-3,6-dimethyl-7-(1-methylethenyl)-, [3S-(3.alpha.,3a.alpha.,6.alpha.,7.beta.,7a.beta.)]- |  | −5.6 | 0.00–0.00 | - | GLU 166 (8/1), (2.80–3.79) HIS 41 (5/1), (3.48–3.96) CYS 145 (1/1), (3.93) |
| e | Vanillosmin |  | −6.4 | 30.75–33.33 | - | HIS 41 (4/1), (2.65–3.75), CYS 145 (1/1), (3.81) |
| f | Piperine |  | −6.0 | 30.24–32.01 | - | GLU 166 (5/1), (2.15–3.85) CYS 145 (2/1), (3.64–3.82) |
| g | Stigmasterol |  | −6.8 | 31.13–33.56 | - | GLU 166 (7/1), (2.57–3.69) HIS 41 (5/1), (3.61–3.89) CYS 145 (3/1), (3.78–3.98) |
| h | Di(1,2,5-oxadiazolo)[3,4-b:3,4-E]pyrazine, 4,8-diacetyl- |  | −6.5–−5.7 | 26.80–33.89 | PHE 140 (1/1), HIS 163 (1/1), HIS 164 (1/1) | GLU 166 (14/2), (2.49–3.62) HIS 41 (2/1), (2.96–3.69) CYS 145 (1/1), (3.62) |
| i | Cycloeucalenol acetate |  | −7.0–6.4 | 25.29–34.14 | THR 26 (1/1) LYS 137 (1/1) MET 276 (1/1) | GLU 166 (8/2), (2.67–3.81) HIS 41 (18/3), (2.87–3.90) CYS 145 (2/2), (3.73–3.77) |
| k | Strophanthidol |  | −7.2–6.7 | 0.00–3.98 | THR 26 (1/1) GLU 166 (3/2) GLN 198 (1/1) | GLU 166 (27/3), (1.82–3.69) HIS 41 (8/3), (2.95–3.64) CYS 145 (2/2), (2.94–3.52) |
| l | Eudesma-5,11(13)-dien-8,12-olide |  | −6.1–5.9 | 29.56–33.61 | - | GLU 166 (2/1), (3.03–3.87) HIS 41(34/2), (1.70–3.96) CIS 145 (3/2), (3.77–3.88) |
| m | 2-Butenoic acid, 2-methyl-, 2-(acetyloxy)-1,1a,2,3,4,6,7,10,11,11a-decahydro-7,10-dihydroxy-1,1,3,6,9-pentamethyl-4a,7a-epoxy-5 |  | −6.6–6.1 | 29.19–32.40 | (ASN 142)2/2 GLU 166 | GLU 166 (6/2), (1.99–3.20) HIS 41(14/2), (1.62–3.78) CIS 145 (6/2), (2.54–3.64) |
| Microorganisms | Acetic Acid Extract (100 mg/mL) | Chloramphgenicol (2.5 mg/mL) | Clotrimazole (5 mg/mL) | DMSO (10% v/v) |
|---|---|---|---|---|
| Staphylococcus aureus | 16.5 ± 0.7 | 26.0 ± 1.4 | NA | 6.0 ± 0.0 |
| Bacillus cereus | 25.0 ± 2.8 | 22.5 ± 0.7 | NA | 6.0 ± 0.0 |
| Pseudomonas aeruginosa | 13.5 ± 0.7 | 15.5 ± 0.7 | NA | 6.0 ± 0.0 |
| Escherichia coli | 14.0 ± 1.4 | 24.5 ± 0.7 | NA | 6.0 ± 0.0 |
| Salmonella enterica | 18.0 ± 0.0 | 23.0 ± 1.4 | NA | 6.0 ± 0.0 |
| Candida albicans | 38.5 ± 2.1 | NA | 16.0 ± 1.4 | 6.0 ± 0.0 |
| Microorganism | MIC (mg/mL) | MBC or MFC (mg/mL) | MBC/MIC or MFC/MIC |
|---|---|---|---|
| Staphylococcus aureus | 25 | 25 | 1 |
| Bacillus cereus | 25 | 50 | 2 |
| Escherichia coli | 25 | 50 | 2 |
| Salmonella enterica | 25 | 50 | 2 |
| Pseudomonas aeruginosa | 25 | 50 | 2 |
| Candida albicans | 6.25 | 12.5 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idriss, H.; Siddig, B.; González-Maldonado, P.; Elkhair, H.M.; Alakhras, A.I.; Abdallah, E.M.; Elzupir, A.O.; Sotelo, P.H. Inhibitory Activity of Saussurea costus Extract against Bacteria, Candida, Herpes, and SARS-CoV-2. Plants 2023, 12, 460. https://doi.org/10.3390/plants12030460
Idriss H, Siddig B, González-Maldonado P, Elkhair HM, Alakhras AI, Abdallah EM, Elzupir AO, Sotelo PH. Inhibitory Activity of Saussurea costus Extract against Bacteria, Candida, Herpes, and SARS-CoV-2. Plants. 2023; 12(3):460. https://doi.org/10.3390/plants12030460
Chicago/Turabian StyleIdriss, Hajo, Babeker Siddig, Pamela González-Maldonado, H. M. Elkhair, Abbas I. Alakhras, Emad M. Abdallah, Amin O. Elzupir, and Pablo H. Sotelo. 2023. "Inhibitory Activity of Saussurea costus Extract against Bacteria, Candida, Herpes, and SARS-CoV-2" Plants 12, no. 3: 460. https://doi.org/10.3390/plants12030460