
Colletotrichum echinochloae: A Potential Bioherbicide Agent for Control of Barnyardgrass (Echinochloa crus-galli (L.) Beauv.)

Abstract
:1. Introduction
2. Results
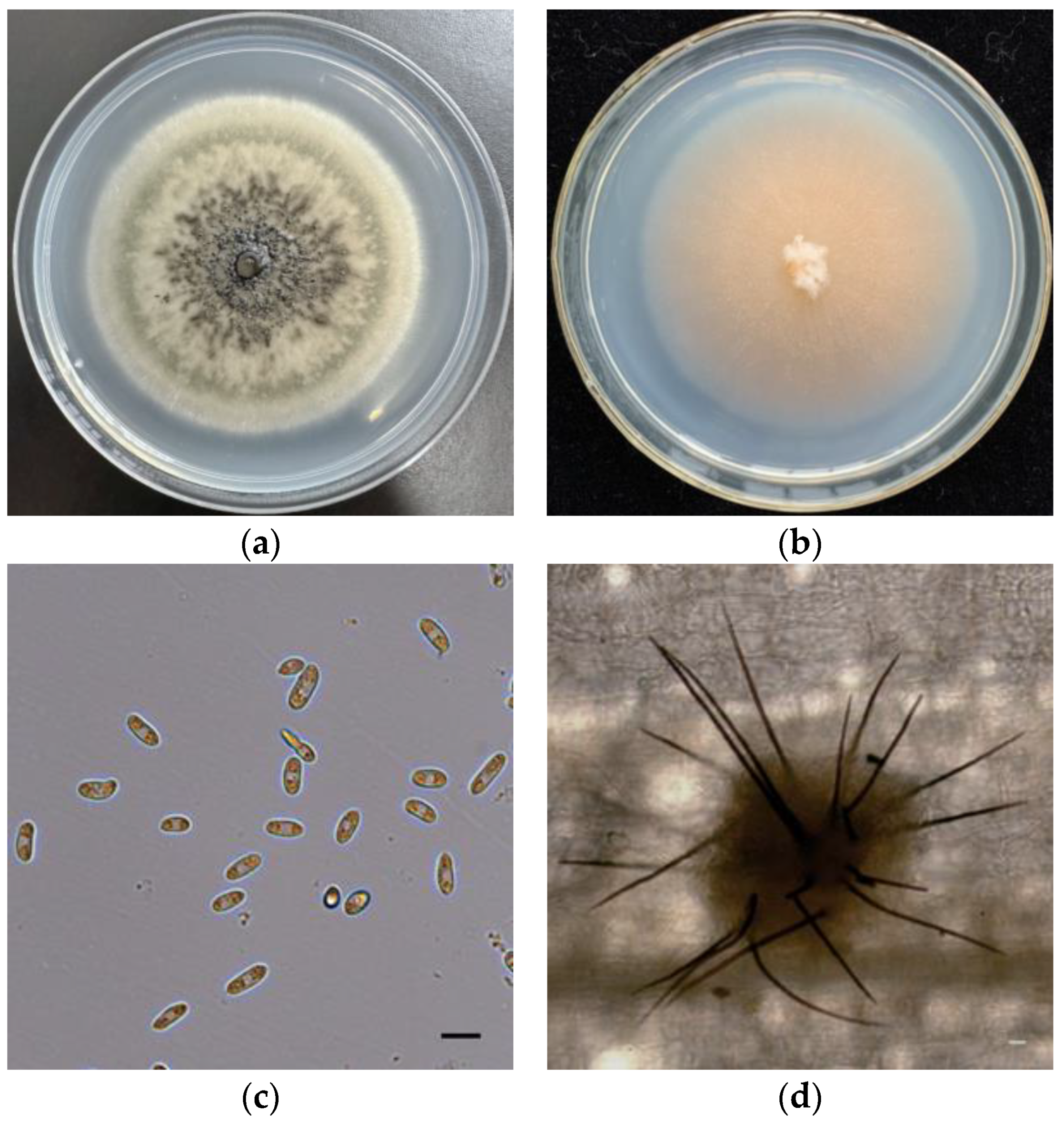
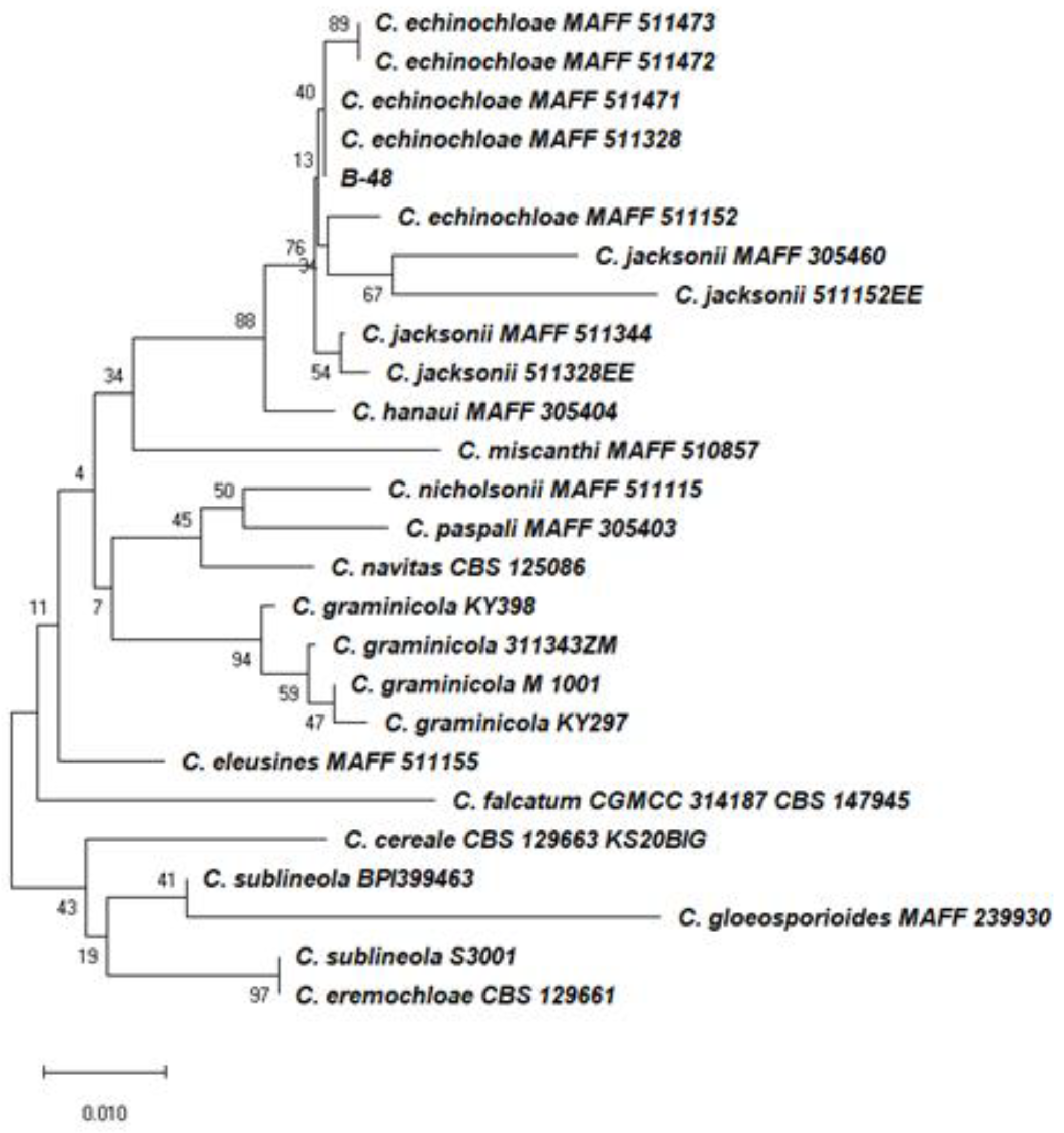
2.1. Isolation and Identification of Fungal Pathogens for the Control of Barnyardgrass
2.2. Host-Range Studies
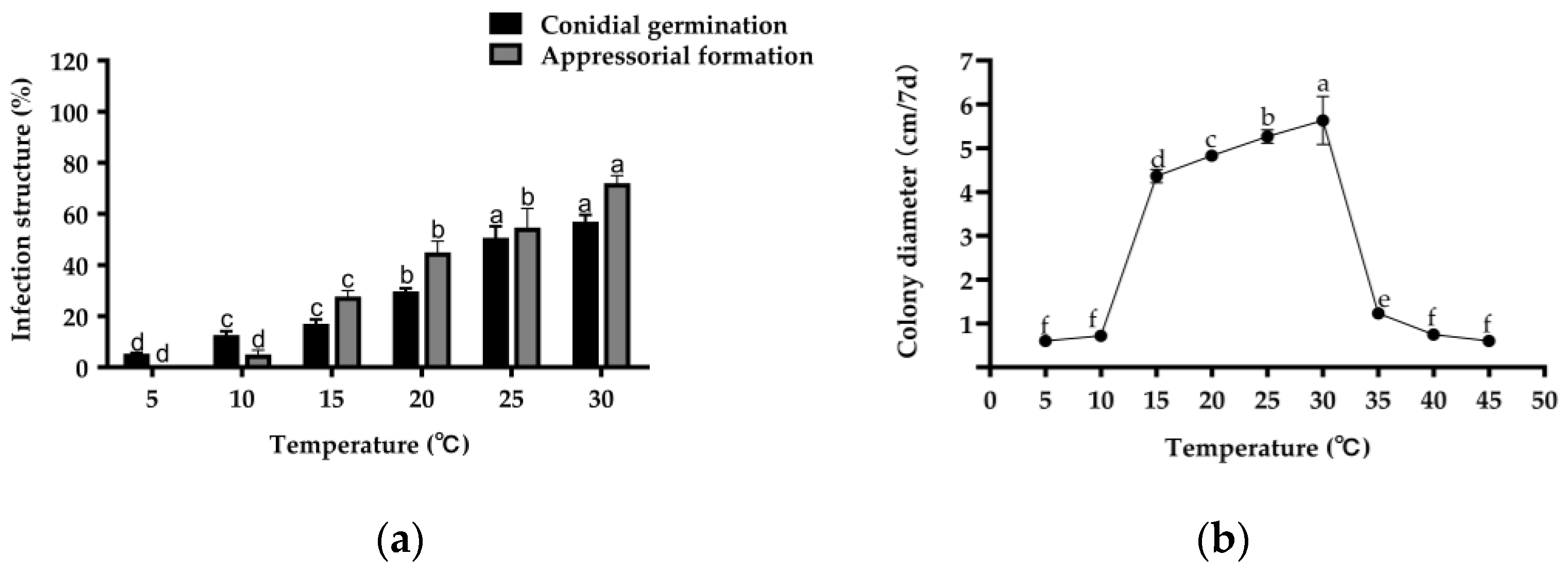
2.3. Effect of Different Temperatures on C. echinochloae Conidial Germination, Appressorial Formation, and Mycelial Growth
2.4. Effect of C. echinochloae Conidia Concentration on Weed Control
2.5. Effect of Weed Growth Stage on Biocontrol Efficacy
2.6. Effect of Dew Period on Biocontrol Efficacy
3. Discussion
4. Materials and Methods
4.1. Isolation, Identification, and Characterization of C. echinochloae B-48
4.2. Plant Preparation, Pathogen Inoculum Preparation, and Plant Inoculation
4.3. Disease Assessment
4.4. Host-Range Studies
4.5. Effect of Temperature on C. echinochloae Conidial Germination, Appressorial Formation, and Mycelial Growth
4.6. Effect of C. echinochloae Conidial Concentration on Weed Control
4.7. Effect of Weed Growth Stage on Weed Biocontrol Efficacy
4.8. Effect of the Dew Period on Weed Biocontrol Efficacy
4.9. Data Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bajwa, A.A.; Jabran, K.; Shahid, M.; Ali, H.H.; Chauhan, B.S. Eco-biology and management of Echinochloa crus-galli. Crop Prot. 2015, 75, 151–162. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Johnson, D.E. Ecological studies on Echinochloa crus-galli and the implications for weed management in direct-seeded rice. Crop Prot. 2011, 30, 1385–1391. [Google Scholar] [CrossRef]
- Maun, M.A.; Barrett, S.C.H. The biology of Canadian weeds: 77. Echinochloa crus-galli (L.) Beauv. Can. J. Plant Sci. 1986, 66, 739–759. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, J.; Gu, T.; Yang, X.; Peng, Q.; Bai, L.; Li, Y. Co-planted barnyardgrass reduces rice yield by inhibiting plant above-and belowground-growth during post-heading stages. Crop J. 2021, 9, 1198–1207. [Google Scholar] [CrossRef]
- Selig, C.; de Mol, F.; Westerman, P.R.; Gerowitt, B. Quantifying seed and establishment limitation to seedling recruitment of arable weeds: An example of barnyardgrass (Echinochloa crus-galli). Weed Sci. 2022, 70, 87–94. [Google Scholar] [CrossRef]
- Netto, A.G.; Christoffoleti, P.; VanGessel, M.; Carvalho, S.J.; Nicolai, M.; Brunharo, C. Seed Production, Dissemination, and Weed Seedbanks. In Persistence Strategy of Weeds; John Wiley & Sons, Inc.: Hoboken, USA, 2022; pp. 19–42. [Google Scholar]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Mahajan, G.; Chauhan, B.S. Nonconventional weed management strategies for modern agriculture. Weed Sci. 2015, 63, 723–747. [Google Scholar] [CrossRef]
- Mennan, H.; Ngouajio, M.; Sahin, M.; Isk, D.; Altop, E.K. Competitiveness of rice (Oryza sativa L.) cultivars against Echinochloa crus-galli (L.) Beauv. in water-seeded production systems. Crop Prot. 2012, 41, 1–9. [Google Scholar] [CrossRef]
- Kacan, K.; Tursun, N.; Ullah, H.; Datta, A. Barnyardgrass (Echinochloa crus-galli (L.) P. Beauv.) resistance to acetolactate synthase-inhibiting and other herbicides in rice in Turkey. Plant Soil Environ. 2020, 66, 357–365. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhang, Z.Y.; Xu, X.M.; Li, G. The density of barnyard grass affects photosynthesis and physiological characteristics of rice. Photosynthetica 2019, 57, 705–711. [Google Scholar] [CrossRef]
- Hashimi, M.H.; Hashimi, R.; Ryan, Q. Toxic effects of pesticides on humans, plants, animals, pollinators and beneficial organisms. APRJ 2020, 5, 37–47. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Mohsin, S.M.; Bhuyan, M.B.; Bhuiyan, T.F.; Anee, T.I.; Masud, A.A.C.; Nahar, K. Phytotoxicity, environmental and health hazards of herbicides: Challenges and ways forward. In Agrochemicals Detection Treatment and Remediation; Butterworth-Heinemann: Oxford, UK, 2020; pp. 55–99. [Google Scholar]
- Rashid, B.; Husnain, T.; Riazuddin, S. Herbicides and pesticides as potential pollutants: A global problem. In Plant Adaptation and Phytoremediation; Springer: Dordrecht, The Netherlands, 2010; pp. 427–447. [Google Scholar]
- Cerdeira, A.L.; Duke, S.O. The current status and environmental impacts of glyphosate-resistant crops: A review. J. Environ. Qual. 2006, 35, 1633–1658. [Google Scholar] [CrossRef] [PubMed]
- Mithila, J.; Hall, J.C.; Johnson, W.G.; Kelley, K.B.; Riechers, D.E. Evolution of resistance to auxinic herbicides: Historical perspectives, mechanisms of resistance, and implications for broadleaf weed management in agronomic crops. Weed Sci. 2011, 59, 445–457. [Google Scholar] [CrossRef]
- Heap, I. Herbicide resistant weeds. In Integrated Pest Management; Springer: Dordrecht, The Netherlands, 2014; pp. 281–301. [Google Scholar]
- Cai, X.; Chen, J.; Wang, X.; Gao, H.; Xiang, B.; Dong, L. Mefenacet resistance in multiple herbicide-resistant Echinochloa crus-galli L. populations. Pestic. Biochem. Physiol. 2022, 182, 105038. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.I.; Norsworthy, J.K.; González-Torralva, F.; Priess, G.L.; Barber, L.T.; Butts, T.R. Non-target-site resistance mechanism of barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.] to florpyrauxifen-benzyl. Pest Manag. Sci. 2022, 78, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Y.; Wang, L.; Liu, W.; Wang, J. Baseline sensitivity of Echinochloa crus-galli (L.) P. Beauv. to tripyrasulfone a new HPPD-inhibiting herbicide, in China. Crop Prot. 2022, 158, 105993. [Google Scholar] [CrossRef]
- Iwakami, S.; Hashimoto, M.; Matsushima, K.I.; Watanabe, H.; Hamamura, K.; Uchino, A. Multiple-herbicide resistance in Echinochloa crus-galli var. formosensis an allohexaploid weed species, in dry-seeded rice. Pestic. Biochem. Physiol. 2015, 119, 1–8. [Google Scholar] [CrossRef]
- Steingrímsdóttir, M.M.; Petersen, A.; Fantke, P. A screening framework for pesticide substitution in agriculture. J. Clean. Prod. 2018, 192, 306–315. [Google Scholar] [CrossRef]
- Morin, L. Progress in biological control of weeds with plant pathogens. Annu. Rev. Phytopathol. 2020, 58, 201–223. [Google Scholar] [CrossRef]
- Hinz, H.L.; Winston, R.L.; Schwarzländer, M. How safe is weed biological control? A global review of direct nontarget attack. Q. Rev. Biol. 2019, 94, 1–27. [Google Scholar] [CrossRef]
- Kumar, V.I.K.A.S.; Singh, M.A.N.O.J.; Sehrawat, N.I.R.M.A.L.A.; Atri, N.I.D.H.I.; Singh, R.; Upadhyay, S.K.; Yadav, M.U.K.E.S.H. Mycoherbicide control strategy: Concept, constraints, and advancements. Biopestic. Int. 2021, 17, 29–40. [Google Scholar]
- Goodwin, P.H. A molecular weed-mycoherbicide interaction: Colletotrichum gloeosporioides f. sp. malvae and round-leaved mallow, Malva pusilla. Can. J. Plant Pathol. 2001, 23, 28–35. [Google Scholar]
- Kirkpatrick, T.L.; Templeton, G.E.; TeBeest, D.O.; Smith, R.J., Jr. Potential of Colletotrichum malvarum for biological control of prickly sida. Plant Dis. 1983, 66, 323–325. [Google Scholar] [CrossRef]
- Bowers, R.C. Commercialization of Collego™–An Industrialist’s View. Weed Sci. 1986, 34, 24–25. [Google Scholar]
- Chakraborty, A.; Ray, P. Mycoherbicides for the Noxious Meddlesome: Can Colletotrichum be a Budding Candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Li, X.H.; Liu, M.; Zhang, W.; Yan, J.Y. Mycosphere essay 16: Colletotrichum: Biological control, biocatalyst, secondary metabolites and toxins. Mycosphere 2016, 7, 1164–1176. [Google Scholar] [CrossRef]
- Boyette, C.D.; Gealy, D.; Hoagland, R.E.; Vaughn, K.C.; Bowling, A.J. Hemp sesbania (Sesbania exaltata) control in rice (Oryza sativa) with the bioherbicidal fungus Colletotrichum gloeosporioides f. sp. aeschynomene formulated in an invert emulsion. Biocontrol. Sci. Technol. 2011, 21, 1399–1407. [Google Scholar] [CrossRef]
- Auld, B.A.; Say, M.M.; Ridings, H.I.; Andrews, J. Field applications of Colletotrichum orbiculare to control Xanthium spinosum. Agr. Ecosyst. Environ. 1990, 32, 315–323. [Google Scholar] [CrossRef]
- Auld, B.A.; McRae, C.F.; Say, M.M. Potential for Xanthium spinosum control by Colletotrichum orbiculare as a mycoherbicide. In Proceedings VII Symposium of the Biological Control of Weeds; Delfosse, E.S., Ed.; Istituto Sperimentale Patologia Vegetale MAF: Rome, Italy, 1989; pp. 435–443. [Google Scholar]
- Li, J.; Li, M.; GAO, X.X.; Fang, F.; DONG, L.H. Biological characteristics of Lubao No. 1 biological control agent (Colletotrichum gloeosporioides) and construction of a T-DNA insertional mutant library. Acta Prataculturae Sin. 2017, 26, 142. [Google Scholar]
- Morris, M.J. A method for controlling Hakea sericea Schrad. seedlings using the fungus Colletotrichum gloeosporioides (Penz.) Sacc. Weed Res. 1989, 29, 449–454. [Google Scholar] [CrossRef]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O’Connell, R.J.; Narusaka, Y.; Shirasu, K. Comparative genomic and transcriptomic analyses reveal the hemibiotrophic stage shift of Colletotrichum fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K. Auxin herbicides: Current status of mechanism and mode of action. Pest Manag Sci. 2010, 66, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Mathur, M.; Gehlot, P. Recruit the Plant Pathogen for Weed Management: Bioherbicide–A Sustainable Strategy. In Fungi and Their Role in Sustainable Development: Current Perspectives; Springer: Singapore, 2018; pp. 159–181. [Google Scholar]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An eco-friendly tool for sustainable weed management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Auld, B.A.; Hetherington, S.D.; Smith, H.E. Advances in bioherbicide formulation. Weed Biol. Manag. 2003, 3, 61–67. [Google Scholar] [CrossRef]
- Kremer, R.J. The role of bioherbicides in Weed Management. Biopestic. Int. 2005, 1, 127–141. [Google Scholar]
- Rosskopf, E.N.; Yandoc, C.B.; Charudattan, R.; DeValerio, J.T. Influence of epidemiological factors on the bioherbicidal efficacy of Phomopsis amaranthicola on Amaranthus hybridus. Plant Dis. 2005, 89, 1295–1300. [Google Scholar] [CrossRef]
- Makowski, R.M.D. Effect of inoculum concentration, temperature, dew period, and plant growth stage on disease of round-leaved mallow and velvetleaf by Colletotrichum gloeosporioides f. sp. malvae. Phytopathology 1993, 83, 1229–1234. [Google Scholar] [CrossRef]
- Bradley, D.J.; Gilbert, G.S.; Parker, I.M. Susceptibility of clover species to fungal infection: The interaction of leaf surface traits and environment. Am. J. Bot. 2003, 90, 857–864. [Google Scholar] [CrossRef]
- Shabana, Y.M. The use of oil emulsions for improvising the efficacy of Alternaria eichhorniae as a mycoherbicide for water hyacinth (Eichhornia crassipes). Biol. Control. 2005, 32, 78–89. [Google Scholar] [CrossRef]
- Boyette, C.D.; Hoagland, R.E. Adjuvant and refined corn oil formulation effects on conidial germination, appressorial formation and virulence of the bioherbicide, Colletotrichum truncatum. Plant Pathol. J. 2013, 12, 50–60. [Google Scholar] [CrossRef]
- Crouch, J.A.; Clarke, B.B.; White, J.F., Jr.; Hillman, B.I. Systematic analysis of the falcate-spored graminicolous Colletotrichum and a description of six new species from warm-season grasses. Mycologia 2009, 101, 717–732. [Google Scholar] [CrossRef] [PubMed]
- Crouch, J.A.; Clarke, B.B.; Hillman, B.I. What is the value of ITS sequence data in Colletotrichum systematics and species diagnosis? A case study using the falcate-spored graminicolous Colletotrichum group. Mycologia 2009, 101, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Templeton, G.E. Use of Colletotrichum strains as mycoherbicides. In Colletotrichum: Biology, Pathology and Control; CAB International: Wallingford, UK, 1992; pp. 358–380. [Google Scholar]
- Di Tommaso, A.; Watson, A.K.; Hallett, S.G. Infection by the fungal pathogen Colletotrichum coccodes affects velvetleaf (Abutilon theophrasti)-soybean competition in the field. Weed Sci. 1996, 44, 924–933. [Google Scholar] [CrossRef]
- Wymore, L.A.; Watson, A.K.; Gotlieb, A.R. Interaction between Colletotrichum coccodes and thidiazuron for control of velvetleaf (Abutilon theophrasti). Weed Sci. 1987, 35, 377–383. [Google Scholar] [CrossRef]
- Singh, J.; Quereshi, S.; Banerjee, N.; Pandey, A.K. Production and extraction of phytotoxins from Colletotrichum dematium FGCC# 20 effective against Parthenium hysterophorus L. Braz Arch Biol Technol. 2010, 53, 669–678. [Google Scholar]
- Vieira, B.S.; Dias, L.V.S.A.; Langoni, V.D.; Lopes, E.A. Liquid fermentation of Colletotrichum truncatum UFU 280, a potential mycoherbicide for beggartick. Australas Plant Pathol. 2018, 47, 277–283. [Google Scholar] [CrossRef]
- Roberts, J.; Florentine, S.; Fernando, W.D.; Tennakoon, K.U. Achievements, developments and future challenges in the field of bioherbicides for weed control: A global review. Plants 2022, 11, 2242. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Tan, Y.P. Colletotrichum—Names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Moriwaki, J.; Tsukiboshi, T. Colletotrichum echinochloae, a new species on Japanese barnyard millet (Echinochloa utilis). Mycoscience 2009, 50, 273–280. [Google Scholar] [CrossRef]
- Crouch, J.A.; Beirn, L.A. Anthracnose of cereals and grasses. Fungal Divers. 2009, 39, 19. [Google Scholar]
- Obanor, F.O.; Walter, M.; Jones, E.E.; Jaspers, M.V. Effect of temperature, relative humidity, leaf wetness and leaf age on Spilocaea oleagina conidium germination on olive leaves. Eur. J. Plant Pathol. 2008, 120, 211–222. [Google Scholar] [CrossRef]
- Denner, F.D.N.; Kotzé, J.M.; Putterill, J.F. The effect of temperature on spore germination, growth and appressorium formation of Colletotrichum gloeosporioides and Dothiorella aromatica. South Arican Avocado Grow. Assoc. Yearb. 1986, 9, 19–22. [Google Scholar]
- Boyetchko, S.; Peng, G. Challenges and strategies for development of mycoherbicides. Fungal Biotechnol. Agric. Food Environ. Appl. 2004, 21, 111–121. [Google Scholar]
- Hong, J.K.; Hwang, B.K. Influence of inoculum density, wetness duration, plant age, inoculation method, and cultivar resistance on infection of pepper plants by Colletotrichum coccodes. Plant Dis. 1998, 82, 1079–1083. [Google Scholar] [CrossRef]
- Jackson, M.A. Optimizing nutritional conditions for the liquid culture production of effective fungal biological control agents. J. Ind. Microbiol. Biotechnol. 1997, 19, 180–187. [Google Scholar] [CrossRef]
- Nordzieke, D.E.; Sanken, A.; Antelo, L.; Raschke, A.; Deising, H.B.; Pöggeler, S. Specialized infection strategies of falcate and oval conidia of Colletotrichum graminicola. Fungal Genet Biol. 2019, 133, 103276. [Google Scholar] [CrossRef] [PubMed]
- Babu, R.M.; Sajeena, A.; Seetharaman, K. Bioassay of the potentiality of Alternaria alternata (Fr.) keissler as a bioherbicide to control water hyacinth and other aquatic weeds. Crop Prot. 2003, 22, 1005–1013. [Google Scholar] [CrossRef]
- Peng, G.; Bailey, K.L.; Hinz, H.L.; Byer, K.N. Colletotrichum sp: A potential candidate for biocontrol of scentless chamomile (Matricaria perforata) in western Canada. Biocontrol. Sci. Technol. 2005, 15, 497–511. [Google Scholar] [CrossRef]
- Peng, G.; Byer, K.N.; Bailey, K.L. Pyricularia setariae: A potential bioherbicide agent for control of green foxtail (Setaria viridis). Weed Sci. 2004, 52, 105–114. [Google Scholar] [CrossRef]
- Figliola, S.S.; Camper, N.D.; Ridings, W.H. Potential biological control agents for goosegrass (Eleusine indica). Weed Sci. 1988, 36, 830–835. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum–current status and future directions. Stud. Mycol. 2007, 59, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer & Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Abdessemed, N.; Bahet, Y.A.; Zermane, N. Mycoherbicide potential of Alternaria alternata (Fries.) Kiessler and its formulations on the host weed Xanthium strumarium L. Biocontrol. Sci. Technol. 2020, 30, 1300–1315. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species Treated | Plant Reaction |
|---|---|---|
| Gramineae | Maize | I |
| Rice | HR | |
| Wheat | I | |
| Malvaceae | Cotton | I |
| Solanaceae | Tobacco | I |
| Chili | I | |
| Liliaceae | Leek | I |
| Dioscoreaceae | Chinese yam | I |
| Fabaceae | Cowpea | I |
| Family | Plant Species | Plant Reaction |
|---|---|---|
| Amaranthaceae | Celosia argentea L. | I |
| Alternanthera philoxeroides (Mart.) Griseb. | I | |
| Achyranthes bidentata Blume | I | |
| Chenopod | Chenopodium ficifolium Sm. | I |
| Convolvulaceae | Convolvulus arvensis L. | I |
| Malvaceae | Abutilon theophrasti Medikus | I |
| Polygonaceae | Persicaria hydropiper (L.) Spach | I |
| Portulacaceae | Portulaca oleracea L. | I |
| Rubiaceae | Scleromitrion diffusum (Willd.) R.J. Wang | I |
| Scrophulariaceae | Lindernia crustacea (L.) F. Muell. | I |
| Solanaceae | Solanum nigrum L. | I |
| Violaceae | Viola japonica Langsd. ex DC. | I |
| Asteraceae | Eclipta prostrata (L.) L. | I |
| Bidens Pilosa L. | I | |
| Bidens frondose L. | I | |
| Erigeron canadensis L. | I | |
| Symphyotrichum subulatum (Michx.) G. L. Nesom | I | |
| Commelinaceae | Commelina communis L. | I |
| Commelina benghalensis L. | I | |
| Cyperaceae | Cyperus rotundus L. | HR |
| Gramineae | Digitaria sanguinalis (L.) Scop. | I |
| Eleusine indica (L.) Gaertn. | I | |
| Leptochloa chinensis (L.) Nees | I | |
| Setaria viridis (L.) P. Beauv. | I | |
| Leptochloa panicea (Retz.) Ohwi | I |
| Conidial Concentration | Fresh Weight Inhibition Rate (%) | Disease Severity (%) |
|---|---|---|
| 0 | 0 ± 0 d | 0 ± 0 e |
| 104 | 10.07 ± 5.79 c | 8.58 ± 0.09 d |
| 105 | 23.96 ± 0.59 b | 27.99 ± 0.11 c |
| 106 | 34.43 ± 1.71 b | 62.50 ± 0.05 b |
| 107 | 78.34 ± 3.78 a | 73.87 ± 0.04 ab |
| 108 | 84.33 ± 6.50 a | 78.96 ± 0.02 a |
| Leaf Stage | Fresh Weight Inhibition Rate (%) | Disease Severity (%) |
|---|---|---|
| 4 | 79.89 ± 2.66 a | 89.67 ± 0.88 a |
| 5 | 56.34 ± 3.69 b | 83.66 ± 3.17 a |
| 6 | 57.80 ± 2.37 ab | 66.34 ± 4.25 b |
| 7 | 28.91 ± 3.89 c | 64.67 ± 3.48 b |
| 8 | 27.69 ± 2.10 c | 36.16 ± 2.60 c |
| 9 | 24.71 ± 4.47 c | 15.32 ± 4.25 d |
| Wetness Period (h) | Fresh Weight Inhibition Rate (%) | Disease Severity (%) |
|---|---|---|
| 0 | 1.27 ± 1.07 d | 0 ± 0 d |
| 6 | 10.51 ± 7.17 c | 11.67 ± 3.51 c |
| 8 | 21.93 ± 13.09 c | 32.38 ± 4.73 b |
| 10 | 48.34 ± 6.73 b | 78.33 ± 4.04 a |
| 12 | 68.54 ± 5.09 a | 82.67 ± 4.16 a |
| 24 | 79.50 ± 3.01 a | 84.32 ± 4.72 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, Q.; Chu, S.; Huang, Q.; Chen, A.; Li, L.; Li, R. Colletotrichum echinochloae: A Potential Bioherbicide Agent for Control of Barnyardgrass (Echinochloa crus-galli (L.) Beauv.). Plants 2023, 12, 421. https://doi.org/10.3390/plants12030421
Gu Q, Chu S, Huang Q, Chen A, Li L, Li R. Colletotrichum echinochloae: A Potential Bioherbicide Agent for Control of Barnyardgrass (Echinochloa crus-galli (L.) Beauv.). Plants. 2023; 12(3):421. https://doi.org/10.3390/plants12030421
Chicago/Turabian StyleGu, Qiongnan, Shihai Chu, Qichao Huang, Anan Chen, Lin Li, and Ruhai Li. 2023. "Colletotrichum echinochloae: A Potential Bioherbicide Agent for Control of Barnyardgrass (Echinochloa crus-galli (L.) Beauv.)" Plants 12, no. 3: 421. https://doi.org/10.3390/plants12030421