

Drought-Stress-Induced Changes in Chloroplast Gene Expression in Two Contrasting Strawberry Tree (Arbutus unedo L.) Genotypes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Photosynthetic Activity and Chlorophyll Content
2.2. Chloroplast Gene Expression
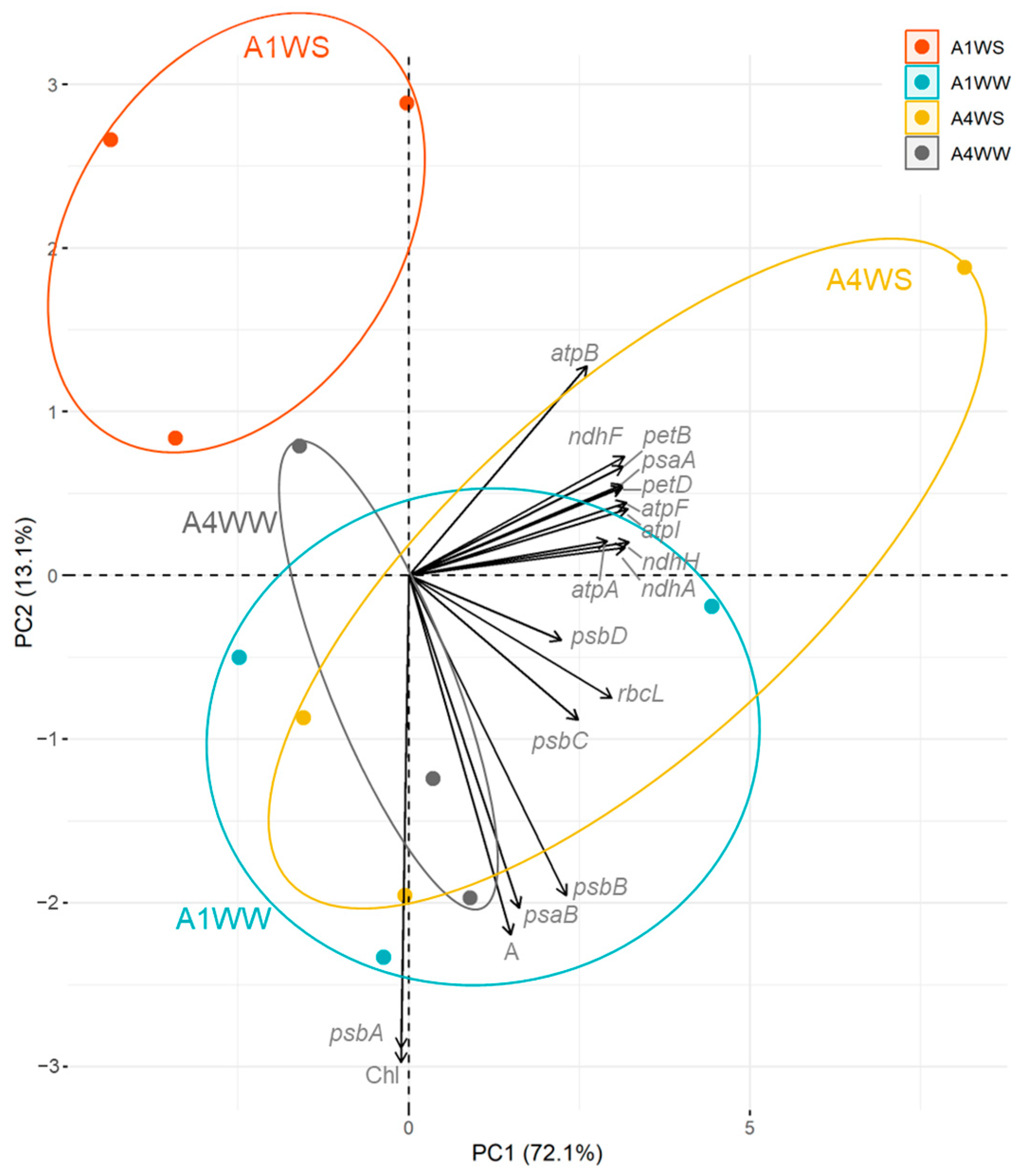
2.3. Correlation between Net CO2 Assimilation, Chlorophyll Content, and Gene Expression
2.4. Predictive Models of Drought Tolerance
3. Discussion
3.1. Photosynthetic Activity and Chlorophyll Content
3.2. Chloroplast Gene Expression
3.3. Correlation between Net CO2 Assimilation, Chlorophyll Content, and Gene Expression
3.4. Predictive Models of Drought Tolerance
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Drought Stress Assay
5.2. Photosynthetic Activity and Chlorophyll Content
5.2.1. Photosynthetic Activity
5.2.2. Chlorophyll Content
5.3. Chloroplast Gene Expression
5.3.1. RNA Extraction and cDNA Synthesis
5.3.2. qPCR
5.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fahad, S.; Bajwa, A.; Nazir, U.; Anjum, S.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, Y.; Ji, P.; Wu, P.; Sheffield, J.; Otkin, J.A. A global transition to flash droughts under climate change. Science 2023, 380, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Yao, T.; Zhang, J.; Xie, M.; Yuan, G.; Tschaplinski, T.; Muchero, W.; Chen, J. Transcriptional regulation of drought response in Arabidopsis and woody plants. Front. Plant Sci. 2021, 11, 572137. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, X.; Shen, X. Regulatory network established by transcription factors transmits drought stress signals in plant. Stress Biol. 2022, 2, 26. [Google Scholar] [CrossRef]
- Bobik, K.; Burch-Smith, T.M. Chloroplast signaling within, between and beyond cells. Front. Plant Sci. 2015, 6, 163163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, A.; Li, X.; Lu, C. The role of chloroplast gene expression in plant responses to environmental stress. Int. J. Mol. Sci. 2020, 21, 6082. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef]
- Zheng, H.; Xin, L.; Guo, J.; Mao, J.; Han, X.; Jia, L.; Zheng, B.; Du, C.; Elmore, R.; Yang, Q.; et al. Adaptation of photosynthesis to water deficit in the reproductive phase of a maize (Zea mays L.) inbred line. Photosynthetica 2019, 57, 399–408. [Google Scholar] [CrossRef]
- Tamburino, R.; Vitale, M.; Ruggiero, A.; Sassi, M.; Sannino, L.; Arena, S.; Costa, A.; Batelli, G.; Zambrano, N.; Scaloni, A.; et al. Chloroplast proteome response to drought stress and recovery in tomato (Solanum lycopersicum L.). BMC Plant Biol. 2017, 17, 40. [Google Scholar] [CrossRef]
- Zadražnik, T.; Moen, A.; Šuštar-Vozlič, J. Chloroplast proteins involved in drought stress response in selected cultivars of common bean (Phaseolus vulgaris L.). 3 Biotech 2019, 9, 331. [Google Scholar] [CrossRef]
- Torres, J.; Valle, F.; Pinto, C.; García-Fuentes, A.; Salazar, C.; Cano, E. Arbutus unedo L. communities in southern Iberian Peninsula mountains. Plant Ecol. 2002, 160, 207–223. [Google Scholar] [CrossRef]
- Martins, J.; Pinto, G.; Canhoto, J. Biotechnology of the multipurpose tree species Arbutus unedo: A review. J. For. Res. 2021, 33, 377–390. [Google Scholar] [CrossRef]
- Lopez-Bustins, J.; Rové, D.; Martin-Vide, J. Mediterranean-scale drought: Regional datasets for exceptional meteorological drought events during 1975–2019. Atmosphere 2021, 12, 941. [Google Scholar] [CrossRef]
- Tuel, A.; Eltahir, E. Why is the Mediterranean a climate change hot spot? J. Clim. 2020, 33, 5829–5843. [Google Scholar] [CrossRef]
- Castell, C.; Terradas, J. Effects of water and nutrient availability on water relations, gas exchange and growth rate of mature plants and resprouts of Arbutus unedo L. Ann. Bot. 1994, 73, 595–602. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Peñuelas, J. Drought-induced oxidative stress in strawberry tree (Arbutus unedo L.) growing in mediterranean field conditions. Plant Sci. 2004, 166, 1105–1110. [Google Scholar] [CrossRef]
- Martins, J.; Correia, S.; Correia, B.; Pinto, G.; Canhoto, J.M. Shoot proliferation and organogenesis on Arbutus unedo: Physiological analysis under water stress. Biol. Plant 2019, 63, 278–286. [Google Scholar] [CrossRef]
- Martins, J.; Monteiro, P.; Pinto, G.; Canhoto, J. Hybridization assays in strawberry tree toward the identification of plants displaying increased drought tolerance. Forests 2021, 12, 148. [Google Scholar] [CrossRef]
- Martins, J.; Pétriacq, P.; Flandin, A.; Gómez-Cadenas, A.; Monteiro, P.; Pinto, G.; Canhoto, J. Genotype determines Arbutus unedo L. physiological and metabolomic responses to drought and recovery. Front. Plant Sci. 2022, 13, 1011542. [Google Scholar] [CrossRef]
- Xu, D.; Tang, Q.; Xu, P.; Schäffner, A.; Leister, D.; Kleine, T. Response of the organellar and nuclear (post)transcriptomes of Arabidopsis to drought. Front. Plant Sci. 2023, 14, 1220928. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.; Zhang, C.; Kurjogi, M.; Pervaiz, T.; Zheng, T.; Zhang, C.; Lide, C.; Shangguan, L.; Fang, J. Insights into grapevine defense response against drought as revealed by biochemical, physiological and RNA-Seq analysis. Sci. Rep. 2017, 7, 13134. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, R.; Zhao, D.; Tao, J. Effects of drought stress on physiological responses and gene expression changes in herbaceous peony (Paeonia lactiflora Pall.). Plant Signal Behav. 2020, 15, 1746034. [Google Scholar] [CrossRef] [PubMed]
- Pelloux, J.; Jolivet, Y.; Fontaine, V.; Banvoy, J.; Dizengremel, P. Changes in Rubisco and Rubisco activase gene expression and polypeptide content in Pinus halepensis M. subjected to ozone and drought. Plant Cell Environ. 2001, 24, 123–131. [Google Scholar] [CrossRef]
- Ji, K.; Wang, Y.; Sun, W.; Lou, Q.; Mei, H.; Shen, S.; Chen, H. Drought-responsive mechanisms in rice genotypes with contrasting drought tolerance during reproductive stage. J. Plant Physiol. 2012, 169, 336–344. [Google Scholar] [CrossRef]
- Zadražnik, T.; Hollung, K.; Egge-Jacobsen, W.; Meglič, V.; Šuštar-Vozlič, J. differential proteomic analysis of drought stress response in leaves of common bean (Phaseolus vulgaris L.). J. Proteom. 2013, 78, 254–272. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Elias, E.; Nawrocki, W.; Croce, R. Drought affects both photosystems in Arabidopsis thaliana. New Phytol. 2023, 240, 663–675. [Google Scholar] [CrossRef]
- Liu, J.; Xie, S.; Luo, Y.; Zhu, G.; Du, L. Soluble expression of spinach PsbC gene in Escherichia coli and in vitro reconstitution of CP43 coupled with chlorophyll a only. Plant Physiol. Bioch. 2014, 79, 19–24. [Google Scholar] [CrossRef]
- Murray, J.; Duncan, J.; Barber, J. CP43-like chlorophyll binding proteins: Structural and evolutionary implications. Trends Plant Sci. 2006, 11, 152–158. [Google Scholar] [CrossRef]
- Bota, J.; Medrano, H.; Flexas, J. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytol. 2004, 162, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Parry, M.; Andralojc, P.; Mitchell, R.; Madgwick, P.; Keys, A. Manipulation of Rubisco: The amount, activity, function and regulation. J. Exp. Bot. 2003, 54, 1321–1333. [Google Scholar] [CrossRef]
- Evans, J.; Clarke, V. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef]
- Huo, Y.; Wang, M.; Wei, Y.; Xia, Z. Overexpression of the maize PsbA gene enhances drought tolerance through regulating antioxidant system, photosynthetic capability, and stress defense gene expression in tobacco. Front. Plant Sci. 2016, 6, 1223. [Google Scholar] [CrossRef]
- Leelavathi, S.; Bhardwaj, A.; Kumar, S.; Dass, A.; Pathak, R.; Pandey, S.; Tripathy, B.; Padmalatha, K.; Dhandapani, G.; Kanakachari, M.; et al. Genome-wide transcriptome and proteome analyses of tobacco PsaA and PsbA deletion mutants. Plant Mol. Biol. 2011, 76, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Sasi, S.; Venkatesh, J.; Daneshi, R.; Gururani, M. Photosystem II extrinsic proteins and their putative role in abiotic stress tolerance in higher plants. Plants 2018, 7, 100. [Google Scholar] [CrossRef]
- Landi, M.; Guidi, L. Effects of abiotic stress on Photosystem II proteins. Photosynthetica 2023, 61, 148–156. [Google Scholar] [CrossRef]
- Anderson, W.C. Tissue culture propagation of red and black raspberries, Rubus idaeus and R. occidentalis. Acta Hortic. 1980, 112, 13–20. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 1 October 2023).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2019. Available online: https://cran.r-project.org/package=factoextra (accessed on 1 October 2023).
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. GGally: Extension to “Ggplot2”. R Package Version 2.1.2. 2021. Available online: https://cran.r-project.org/package=GGally (accessed on 1 October 2023).
- Indahl, U.; Næs, T.; Liland, K. A similarity index for comparing coupled matrices. J. Chemom. 2018, 32, e3049. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, J.; Neves, M.; Canhoto, J. Drought-Stress-Induced Changes in Chloroplast Gene Expression in Two Contrasting Strawberry Tree (Arbutus unedo L.) Genotypes. Plants 2023, 12, 4133. https://doi.org/10.3390/plants12244133
Martins J, Neves M, Canhoto J. Drought-Stress-Induced Changes in Chloroplast Gene Expression in Two Contrasting Strawberry Tree (Arbutus unedo L.) Genotypes. Plants. 2023; 12(24):4133. https://doi.org/10.3390/plants12244133
Chicago/Turabian StyleMartins, João, Mariana Neves, and Jorge Canhoto. 2023. "Drought-Stress-Induced Changes in Chloroplast Gene Expression in Two Contrasting Strawberry Tree (Arbutus unedo L.) Genotypes" Plants 12, no. 24: 4133. https://doi.org/10.3390/plants12244133