
Emphasizing the Role of Long Non-Coding RNAs (lncRNA), Circular RNA (circRNA), and Micropeptides (miPs) in Plant Biotic Stress Tolerance



Abstract
:1. Introduction
2. Types of Non-Coding RNA (ncRNA) and Their Biological Relevance
3. Structural Complexity of Non-Coding RNA
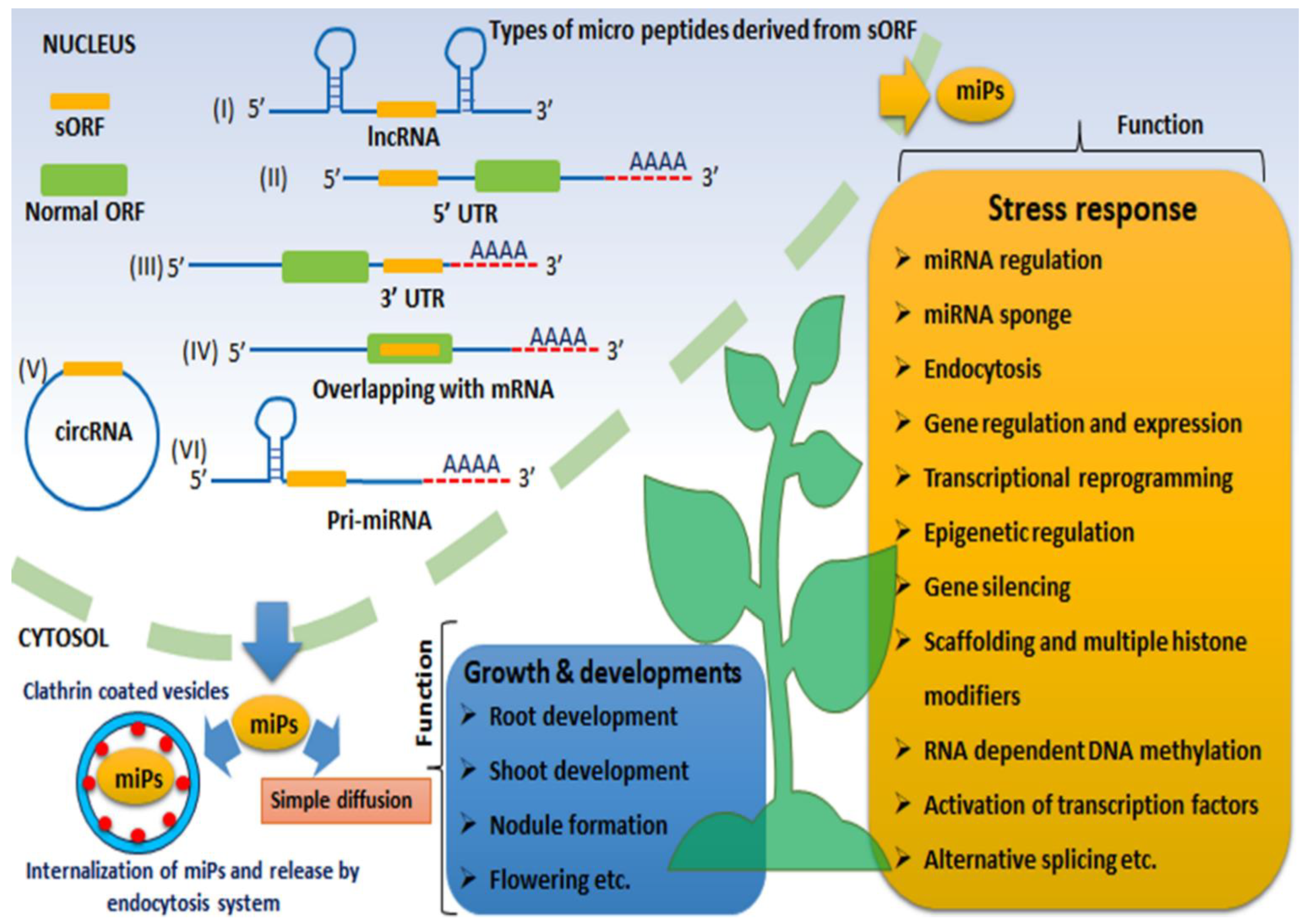
3.1. Short Open Reading Frames (sORFs)
3.2. Long Non-Coding RNAs (lncRNA)
3.3. Circular RNAs (circRNA)
3.4. MiRNA-Derived Peptides (miPEPs)

{kind=link}
{kind=link}
| Sl. No. | Name of Database | Function | References |
|---|---|---|---|
| 1. | PlncRNADB | This is a searchable database of lncRNA sequences and annotation in plants. | [61] |
| 2. | PLNlncRbase | Literature-based database for plant lncRNA for easy curation and determination of biological functions. | [62] |
| 3. | NONCODEV6 | Repository of non-coding RNAs in plants and animals. Tissue-specific expression profile of lncRNA. | [63] |
| 4. | PLncDB | Plant lncRNA database. It includes tissues, developmental stages, mutants, stress treatments, and epigenetic regulation of lncRNA. | [64] |
| 5. | Green Non-Coding Database. GREENC | Pipeline to annotate a large number of plant-specific lncRNAs, including algae. | [65] |
| 6. | MiPepid | A Python-based detection software of sORF using FASTA genomic sequences. | [66] |
| 7. | FuncPEP | This database provides functional peptide identification from non-coding portions of the genome. | [67] |
4. Micropeptides (miPs): Emerging Stars from the “Dark Matter” of Biological Sciences
5. The Regulatory Function of Non-Coding RNA in Biotic Stress Tolerance in Plants
5.1. The Role of lncRNA in Biotic Stress Tolerance in Plants
5.2. The Roles of circRNA in Biotic Stress Tolerance Circuitry in Plants
5.3. The miPEPs and Biotic Stress Tolerance in Plants
6. The Modes of Action of Non-Coding RNAs in Plant Immune Response
6.1. Plant Immunity
6.2. Immune Sensing and Signaling
6.3. Reactive Oxygen Species (ROS) and Hormonal Cross-Talk: A Key Player in Non-Coding-RNA-Mediated Defense Signaling in Plants
7. Conclusions and Future Directions
- The complete micropeptide map of major crop plants and model plants is necessary.
- A stringent yet feasible toolkit and advancing the presently available bioinformatic platform are required.
- An in-depth study is required to decipher the precise functions of miPs in plant–microbe interaction.
- The updating of the existing plant immune system with the proper incorporation of miPs in pathogen recognition, interaction with pattern-triggered immunity (PTI), effector-triggered immunity (ETI), and intracellular signaling is also required.
- Plant immunity is multifaceted signaling. Hence, the interaction of stress-associated miPs with other signaling intermediates and hormonal cross-talk will provide more insights.
- The interaction networks between miP and miRNA need more dissection and follow-ups.
- The potential targets of miPs for genetic engineering and genome editing tools, e.g., clustered regularly interspaced short palindromic repeats (CRISPR) targets of miPs, may revolutionize plant science research in the Anthropocene.
- Dedicated studies on the diversity, conservation, and evolution of miPs among plant species may unveil the eco-evolutionary aspects of plant–pathogen interactions.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, S.; Bhar, A.; Chatterjee, M.; Das, S. Fusarium oxysporum f. sp. ciceri race 1 induced redox state alterations are coupled to downstream defense signaling in root tissues of chickpea (Cicer arietinum L.). PLoS ONE 2013, 8, e73163. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Bhar, A.; Chatterjee, M.; Ghosh, A.; Das, S. Transcriptomic dissection reveals wide spread differential expression in chickpea during early time points of Fusarium oxysporum f. sp. ciceri Race 1 attack. PLoS ONE 2017, 12, e0178164. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Bhar, A.; Chakraborty, A.; Roy, A. Plant responses to biotic stress: Old memories matter. Plants 2022, 11, 84. [Google Scholar] [CrossRef] [PubMed]
- Šečić, E.; Kogel, K.-H.; Ladera-Carmona, M.J. Biotic stress-associated microRNA families in plants. J. Plant Physiol. 2021, 263, 153451. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.E.; Farkas, M.H. Micropeptide. PLoS Genet. 2018, 14, e1007764. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; Guedes, S.; Amado, F.; Santos, M.; Akimitsu, N. The role of micropeptides in biology. Cell. Mol. Life Sci. 2021, 78, 3285–3298. [Google Scholar] [CrossRef] [PubMed]
- Crappé, J.; Van Criekinge, W.; Menschaert, G. Little things make big things happen: A summary of micropeptide encoding genes. EuPA Open Proteom. 2014, 3, 128–137. [Google Scholar] [CrossRef]
- Anderson, D.M.; Anderson, K.M.; Chang, C.-L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Bi, P.; Ramirez-Martinez, A.; Li, H.; Cannavino, J.; McAnally, J.R.; Shelton, J.M.; Sánchez-Ortiz, E.; Bassel-Duby, R.; Olson, E.N. Control of muscle formation by the fusogenic micropeptide myomixer. Science 2017, 356, 323–327. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, M.P.; Krude, T. Functional roles of non-coding Y RNAs. Int. J. Biochem. Cell Biol. 2015, 66, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Rastetter, R.H.; Wilhelm, D. Non-coding RNAs: An introduction. In Non-Coding RNA and the Reproductive System; Springer: Berlin, Germany, 2016; pp. 13–32. [Google Scholar]
- Kovalchuk, I. Non-coding RNAs in genome integrity. In Genome Stability; Elsevier: Amsterdam, The Netherlands, 2021; pp. 453–475. [Google Scholar]
- Makarewich, C.A.; Olson, E.N. Mining for micropeptides. Trends Cell Biol. 2017, 27, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-Z.; Zhu, Q.-F.; Xue, J.; Chen, P.; Yu, Y. Shining in the dark: The big world of small peptides in plants. aBIOTECH 2023, 4, 238–256. [Google Scholar] [CrossRef] [PubMed]
- Leong, A.Z.-X.; Lee, P.Y.; Mohtar, M.A.; Syafruddin, S.E.; Pung, Y.-F.; Low, T.Y. Short open reading frames (sORFs) and microproteins: An update on their identification and validation measures. J. Biomed. Sci. 2022, 29, 19. [Google Scholar] [CrossRef] [PubMed]
- Brunet, M.A.; Leblanc, S.; Roucou, X. Reconsidering proteomic diversity with functional investigation of small ORFs and alternative ORFs. Exp. Cell Res. 2020, 393, 112057. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Hu, J.; Xu, H. Distribution of micropeptide-coding sORFs in transcripts. Chin. Chem. Lett. 2018, 29, 1029–1032. [Google Scholar] [CrossRef]
- Biswas, A.; Sen, B.; Bandyopadhyay, S.; Mal, C. Co-regulatory functions of miRNA and lncRNA in adapting biotic and abiotic stress in economically important dicot plants. Plant Gene 2021, 26, 100275. [Google Scholar] [CrossRef]
- Gai, Y.-P.; Yuan, S.-S.; Zhao, Y.-N.; Zhao, H.-N.; Zhang, H.-L.; Ji, X.-L. A novel LncRNA, MuLnc1, associated with environmental stress in mulberry (Morus multicaulis). Front. Plant Sci. 2018, 9, 669. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Zhang, H.; Zeng, M.; Chen, Q.; Wang, H.; An, H.; Luo, X.; Xu, D.; Wan, P. The long non-coding RNA GhlncRNA149. 1 improves cotton defense response to aphid damage as a positive regulator. Plant Cell Tissue Organ Cult. (PCTOC) 2023, 152, 517–527. [Google Scholar] [CrossRef]
- Ai, G.; Li, T.; Zhu, H.; Dong, X.; Fu, X.; Xia, C.; Pan, W.; Jing, M.; Shen, D.; Xia, A. BPL3 binds the long non-coding RNA nalncFL7 to suppress FORKED-LIKE7 and modulate HAI1-mediated MPK3/6 dephosphorylation in plant immunity. Plant Cell 2023, 35, 598–616. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Jiang, N.; Meng, J.; Yang, G.; Liu, W.; Zhou, X.; Ma, N.; Hou, X.; Luan, Y. LncRNA33732-respiratory burst oxidase module associated with WRKY1 in tomato-Phytophthora infestans interactions. Plant J. 2019, 97, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and characterization of wheat long non-protein coding RNAs responsive to powdery mildew infection and heat stress by using microarray analysis and SBS sequencing. BMC Plant Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, M.; Li, N.; Wang, H.; Qiu, P.; Pei, L.; Xu, Z.; Wang, T.; Gao, E.; Liu, J. Long noncoding RNA s involve in resistance to Verticillium dahliae, a fungal disease in cotton. Plant Biotechnol. J. 2018, 16, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Yan, W.; Li, X.; Yang, S.; Li, J.; Yin, Y.; Yuan, F.; Wang, H.; Kang, Z.; Han, D. Identification of Long Intergenic Noncoding RNAs in Rhizoctonia cerealis following Inoculation of Wheat. Microbiol. Spectr. 2023, 11, e03449-22. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Sharma, V.; Dubey, H.; Singh, P.K.; Kapoor, R.; Kumari, M.; Singh, J.; Pawar, D.V.; Bisht, D.; Solanke, A.U. Identification of long non-coding RNA in rice lines resistant to Rice blast pathogen Maganaporthe oryzae. Bioinformation 2017, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xue, H.-S.; Zhang, D.-D.; Wang, D.; Song, J.; Subbarao, K.V.; Klosterman, S.J.; Chen, J.-Y.; Dai, X.-F. Identification of long non-coding RNAs in Verticillium dahliae following inoculation of cotton. Microbiol. Res. 2022, 257, 126962. [Google Scholar] [CrossRef]
- Tang, J.; Chen, X.; Yan, Y.; Huang, J.; Luo, C.; Tom, H.; Zheng, L. Comprehensive transcriptome profiling reveals abundant long non-coding RNAs associated with development of the rice false smut fungus, Ustilaginoidea virens. Environ. Microbiol. 2021, 23, 4998–5013. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, G.; Zhang, F.; Ma, J.; Wen, C.; Li, H. Genome-wide Identification of Powdery Mildew Responsive Long Noncoding RNAs in Cucurbita pepo. Front. Genet. 2022, 13, 933022. [Google Scholar] [CrossRef]
- Cao, W.; Gan, L.; Wang, C.; Zhao, X.; Zhang, M.; Du, J.; Zhou, S.; Zhu, C. Genome-wide identification and characterization of potato long non-coding RNAs associated with Phytophthora infestans resistance. Front. Plant Sci. 2021, 12, 619062. [Google Scholar] [CrossRef]
- Li, Q.; Shen, H.; Yuan, S.; Dai, X.; Yang, C. miRNAs and lncRNAs in tomato: Roles in biotic and abiotic stress responses. Front. Plant Sci. 2023, 13, 1094459. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Song, Y.; Zhang, L.; Guo, D.; He, J.; Wang, L.; Song, S.; Xu, W.; Zhang, C.; Lers, A. Coding of non-coding RNA: Insights into the regulatory functions of Pri-MicroRNA-encoded peptides in plants. Front. Plant Sci. 2021, 12, 641351. [Google Scholar] [CrossRef] [PubMed]
- Ormancey, M.; Thuleau, P.; Combier, J.-P.; Plaza, S. The Essentials on microRNA-Encoded Peptides from Plants to Animals. Biomolecules 2023, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Gautam, H.; Sharma, A.; Trivedi, P.K. Plant microProteins and miPEPs: Small molecules with much bigger roles. Plant Sci. 2023, 326, 111519. [Google Scholar] [CrossRef] [PubMed]
- Ram, M.K.; Mukherjee, K.; Pandey, D.M. Identification of miRNA, their targets and miPEPs in peanut (Arachis hypogaea L.). Comput. Biol. Chem. 2019, 83, 107100. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, F.J. Small RNAs in Medicago truncatula: Introduction. In Model Legume Medicago Truncatula; Wiley Online Library: Hoboken, NJ, USA, 2020; p. 946. [Google Scholar]
- Chand Jha, U.; Nayyar, H.; Mantri, N.; Siddique, K.H. Non-coding RNAs in legumes: Their emerging roles in regulating biotic/abiotic stress responses and plant growth and development. Cells 2021, 10, 1674. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Guo, B.; Xu, L.; Li, L.; Song, X.; Wang, X.; Zeng, X.; Wu, L.; Niu, D. Exonic Circular RNAs Are Involved in Arabidopsis Immune Response Against Bacterial and Fungal Pathogens and Function Synergistically with Corresponding Linear RNAs. Phytopathology® 2022, 112, 608–619. [Google Scholar] [CrossRef]
- Kalwan, G.; Gill, S.S.; Priyadarshini, P.; Gill, R.; Yadava, Y.K.; Yadav, S.; Baruah, P.M.; Agarwala, N.; Gaikwad, K.; Jain, P.K. Approaches for identification and analysis of plant circular RNAs and their role in stress responses. Environ. Exp. Bot. 2023, 205, 105099. [Google Scholar] [CrossRef]
- Luo, J.; Wang, X.L.; Sun, Z.C.; Wu, D.; Zhang, W.; Wang, Z.J. Progress in circular RNAs of plants. Yi Chuan = Hered. 2018, 40, 467–477. [Google Scholar]
- Raja, I.; Tennyson, J. Small RNA–regulator of biotic stress and pathogenesis in food crops. In Plant Small RNA in Food Crops; Elsevier: Amsterdam, The Netherlands, 2023; pp. 233–269. [Google Scholar]
- Bordoloi, K.S.; Baruah, P.M.; Agarwala, N. Identification of circular RNAs in tea plant during Helopeltis theivora infestation. Plant Stress 2023, 8, 100150. [Google Scholar] [CrossRef]
- Rakoczy-Lelek, R.; Czernicka, M.; Ptaszek, M.; Jarecka-Boncela, A.; Furmanczyk, E.M.; Kęska-Izworska, K.; Grzanka, M.; Skoczylas, Ł.; Kuźnik, N.; Smoleń, S. Transcriptome Dynamics Underlying Planticine®-Induced Defense Responses of Tomato (Solanum lycopersicum L.) to Biotic Stresses. Int. J. Mol. Sci. 2023, 24, 6494. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.-H.; Meng, J.; Zhang, M.; Luan, Y.-S. Identification of tomato circular RNAs responsive to Phytophthora infestans. Gene 2020, 746, 144652. [Google Scholar] [CrossRef]
- Belousova, E.; Filipenko, M.; Kushlinskii, N. Circular RNA: New regulatory molecules. Bull. Exp. Biol. Med. 2018, 164, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Ebbesen, K.K.; Hansen, T.B.; Kjems, J. Insights into circular RNA biology. RNA Biol. 2017, 14, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Yang, L. Regulation of circRNA biogenesis. RNA Biol. 2015, 12, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chu, S.; Jiao, Y. Present scenario of circular RNAs (circRNAs) in plants. Front. Plant Sci. 2019, 10, 379. [Google Scholar] [CrossRef]
- Chu, Q.; Bai, P.; Zhu, X.; Zhang, X.; Mao, L.; Zhu, Q.-H.; Fan, L.; Ye, C.-Y. Characteristics of plant circular RNAs. Brief. Bioinform. 2020, 21, 135–143. [Google Scholar] [CrossRef]
- Liao, X.; Li, X.-J.; Zheng, G.-T.; Chang, F.-R.; Fang, L.; Yu, H.; Huang, J.; Zhang, Y.-F. Mitochondrion-encoded circular RNAs are widespread and translatable in plants. Plant Physiol. 2022, 189, 1482–1500. [Google Scholar] [CrossRef]
- Chu, Q.; Ding, Y.; Xu, X.; Ye, C.Y.; Zhu, Q.H.; Guo, L.; Fan, L. Recent origination of circular RNAs in plants. New Phytol. 2022, 233, 515–525. [Google Scholar] [CrossRef]
- Julkowska, M. Small but powerful: MicroRNA-derived peptides promote grape adventitious root formation. Am. Soc. Plant Biol. 2020, 183, 429–430. [Google Scholar] [CrossRef] [PubMed]
- Lauressergues, D.; Couzigou, J.-M.; Clemente, H.S.; Martinez, Y.; Dunand, C.; Bécard, G.; Combier, J.-P. Primary transcripts of microRNAs encode regulatory peptides. Nature 2015, 520, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Couzigou, J.-M.; André, O.; Guillotin, B.; Alexandre, M.; Combier, J.-P. Use of microRNA-encoded peptide miPEP172c to stimulate nodulation in soybean. New Phytol. 2016, 211, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-j.; Deng, B.-h.; Gao, J.; Zhao, Z.-y.; Chen, Z.-l.; Song, S.-r.; Wang, L.; Zhao, L.-p.; Xu, W.-p.; Zhang, C.-x. A miRNA-encoded small peptide, vvi-miPEP171d1, regulates adventitious root formation. Plant Physiol. 2020, 183, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Badola, P.K.; Bhatia, C.; Sharma, D.; Trivedi, P.K. miRNA-encoded peptide, miPEP858, regulates plant growth and development in Arabidopsis. bioRxiv 2019. [Google Scholar] [CrossRef]
- Ormancey, M.; Le Ru, A.; Duboé, C.; Jin, H.; Thuleau, P.; Plaza, S.; Combier, J.-P. Internalization of miPEP165a into Arabidopsis roots depends on both passive diffusion and endocytosis-associated processes. Int. J. Mol. Sci. 2020, 21, 2266. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Dai, X.; Ye, T.; Zhang, P.; Yan, X.; Gong, X.; Liang, S.; Chen, M. PlncRNADB: A repository of plant lncRNAs and lncRNA-RBP protein interactions. Curr. Bioinform. 2019, 14, 621–627. [Google Scholar] [CrossRef]
- Xuan, H.; Zhang, L.; Liu, X.; Han, G.; Li, J.; Li, X.; Liu, A.; Liao, M.; Zhang, S. PLNlncRbase: A resource for experimentally identified lncRNAs in plants. Gene 2015, 573, 328–332. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D. NONCODEV6: An updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef]
- Jin, J.; Liu, J.; Wang, H.; Wong, L.; Chua, N.-H. PLncDB: Plant long non-coding RNA database. Bioinformatics 2013, 29, 1068–1071. [Google Scholar] [CrossRef]
- Gallart, A.P.; Pulido, A.H.; de Lagrán, I.A.M.; Sanseverino, W.; Cigliano, R.A. GREENC: A Wiki-based database of plant lncRNAs. Nucleic Acids Res. 2016, 44, D1161. [Google Scholar]
- Zhu, M.; Gribskov, M. MiPepid: MicroPeptide identification tool using machine learning. BMC Bioinform. 2019, 20, 559. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, M.P.; Manyam, G.C.; Ott, L.F.; Berland, L.; Knutsen, E.; Ivan, C.; Lipovich, L.; Broom, B.M.; Calin, G.A. FuncPEP: A database of functional peptides encoded by non-coding RNAs. Non-Coding RNA 2020, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Chen, R.; Qu, L.; Cao, X. Noncoding RNA: From dark matter to bright star. Sci. China Life Sci. 2020, 63, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.H.; Stephen, S.; Taylor, J.; Helliwell, C.A.; Wang, M.B. Long noncoding RNA s responsive to Fusarium oxysporum infection in Arabidopsis thaliana. New Phytol. 2014, 201, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.S.; Sun, H.-X.; Park, B.S.; Huang, C.-H.; Yeh, S.-D.; Jung, C.; Chua, N.-H. ELF18-INDUCED LONG-NONCODING RNA associates with mediator to enhance expression of innate immune response genes in Arabidopsis. Plant Cell 2017, 29, 1024–1038. [Google Scholar] [CrossRef] [PubMed]
- Zafar, J.; Huang, J.; Xu, X.; Jin, F. Analysis of Long Non-Coding RNA-Mediated Regulatory Networks of Plutella xylostella in Response to Metarhizium anisopliae Infection. Insects 2022, 13, 916. [Google Scholar] [CrossRef]
- Fass, M.I.; Rivarola, M.; Ehrenbolger, G.F.; Maringolo, C.A.; Montecchia, J.F.; Quiroz, F.; García-García, F.; Blázquez, J.D.; Hopp, H.E.; Heinz, R.A. Exploring sunflower responses to Sclerotinia head rot at early stages of infection using RNA-seq analysis. Sci. Rep. 2020, 10, 13347. [Google Scholar] [CrossRef]
- Bhatia, G.; Upadhyay, S.K.; Singh, K. Vitis vinifera (grapevine) lncRNAs are potential regulators of response to necrotrophic fungus, Botrytis cinerea infection. Physiol. Mol. Plant Pathol. 2020, 112, 101553. [Google Scholar] [CrossRef]
- Choi, G.; Jeon, J.; Lee, H.; Zhou, S.; Lee, Y.-H. Genome-wide profiling of long non-coding RNA of the rice blast fungus Magnaporthe oryzae during infection. BMC Genom. 2022, 23, 132. [Google Scholar] [CrossRef]
- Zhou, X.; Cui, J.; Cui, H.; Jiang, N.; Hou, X.; Liu, S.; Gao, P.; Luan, Y.; Meng, J.; Luan, F. Identification of lncRNAs and their regulatory relationships with target genes and corresponding miRNAs in melon response to powdery mildew fungi. Gene 2020, 735, 144403. [Google Scholar] [CrossRef]
- Zhuo, X.; Yu, Q.; Russo, R.; Zhang, Y.; Wei, X.; Wang, Y.Z.; Holden, P.M.; Gmitter, F.G., Jr. Role of long non-coding RNA in regulatory network response to Candidatus Liberibacter asiaticus in citrus. Front. Plant Sci. 2023, 14, 1090711. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhu, J.; Qian, N.; Guo, J.; Yan, C. Bacillus subtilis SL18r induces tomato resistance against Botrytis cinerea, involving activation of long non-coding RNA, MSTRG18363, to decoy miR1918. Front. Plant Sci. 2021, 11, 634819. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhao, D.; Fan, H.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Xuan, Y.; Chen, L. Functional analysis of long non-coding RNAs reveal their novel roles in biocontrol of bacteria-induced tomato resistance to Meloidogyne incognita. Int. J. Mol. Sci. 2020, 21, 911. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhou, Y.F.; Feng, Y.Z.; He, H.; Lian, J.P.; Yang, Y.W.; Lei, M.Q.; Zhang, Y.C.; Chen, Y.Q. Transcriptional landscape of pathogen-responsive lnc RNA s in rice unveils the role of ALEX 1 in jasmonate pathway and disease resistance. Plant Biotechnol. J. 2020, 18, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Rawal, H.C.; Ali, S.; Mondal, T.K. Role of non-coding RNAs against salinity stress in Oryza species: Strategies and challenges in analyzing miRNAs, tRFs and circRNAs. Int. J. Biol. Macromol. 2023, 242, 125172. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zou, L.-H.; Wu, J.; Ramakrishnan, M.; Gao, Y.; Zhao, L.; Zhou, M. The pattern of DNA methylation alteration, and its association with the expression changes of non-coding RNAs and mRNAs in Moso bamboo under abiotic stress. Plant Sci. 2022, 325, 111451. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Y.; Kong, B.; Song, X.; Gao, Z.; Li, X. Identification and Characterization of circRNAs under Drought Stress in Moso Bamboo (Phyllostachys edulis). Forests 2022, 13, 426. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, Y.; Liu, H.; Liang, Z.; Zhang, M.; Zou, C.; Yuan, G.; Gao, S.; Pan, G.; Shen, Y. A Combination of a Genome-Wide Association Study and a Transcriptome Analysis Reveals circRNAs as New Regulators Involved in the Response to Salt Stress in Maize. Int. J. Mol. Sci. 2022, 23, 9755. [Google Scholar] [CrossRef]
- Sun, J.; Dong, Y.; Wang, C.; Xiao, S.; Jiao, Z.; Gao, C. Identification and characterization of melon circular RNAs involved in powdery mildew responses through comparative transcriptome analysis. PeerJ 2021, 9, e11216. [Google Scholar] [CrossRef]
- Ghorbani, A.; Izadpanah, K.; Tahmasebi, A.; Afsharifar, A.; Moghadam, A.; Dietzgen, R.G. Characterization of maize miRNAs responsive to maize Iranian mosaic virus infection. 3 Biotech 2022, 12, 69. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, P.; Fan, Y.; Lu, Q.; Li, Q.; Yan, J.; Muehlbauer, G.J.; Schnable, P.S.; Dai, M.; Li, L. Circular RNAs mediated by transposons are associated with transcriptomic and phenotypic variation in maize. New Phytol. 2018, 217, 1292–1306. [Google Scholar] [CrossRef] [PubMed]
- Basso, M.F.; Ferreira, P.C.G.; Kobayashi, A.K.; Harmon, F.G.; Nepomuceno, A.L.; Molinari, H.B.C.; Grossi-de-Sa, M.F. Micro RNAs and new biotechnological tools for its modulation and improving stress tolerance in plants. Plant Biotechnol. J. 2019, 17, 1482–1500. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Hanada, K.; Kondo, T.; Shinozaki, K. Hormone-like peptides and small coding genes in plant stress signaling and development. Curr. Opin. Plant Biol. 2019, 51, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Castellano, S.; Andrade-Marcial, M.; Aguilar-Méndez, E.D.; Loyola-Vargas, V.M.; de Folter, S.; De-la-Pena, C. MiRNA expression analysis during somatic embryogenesis in Coffea canephora. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 150, 177–190. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Bhar, A.; Gupta, S.; Chatterjee, M.; Das, S. Redox Regulatory Networks in Response to Biotic Stress in Plants: A New Insight Through Chickpea-Fusarium Interplay. Mech. Plant Horm. Signal. Under Stress 2017, 2, 23–43. [Google Scholar]
- Bhar, A.; Chakraborty, A.; Roy, A. The captivating role of calcium in plant-microbe interaction. Front. Plant Sci. 2023, 14, 1138252. [Google Scholar] [CrossRef]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Arenas-Huertero, C.; Chua, N.-H. Genome-wide analysis uncovers regulation of long intergenic noncoding RNAs in Arabidopsis. Plant Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef]
- Rosli, H.G.; Sirvent, E.; Bekier, F.N.; Ramos, R.N.; Pombo, M.A. Genome-wide analysis uncovers tomato leaf lncRNAs transcriptionally active upon Pseudomonas syringae pv. tomato challenge. Sci. Rep. 2021, 11, 24523. [Google Scholar] [CrossRef]
- Shivaprasad, P.V.; Chen, H.-M.; Patel, K.; Bond, D.M.; Santos, B.A.; Baulcombe, D.C. A microRNA superfamily regulates nucleotide binding site–leucine-rich repeats and other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Shi, Y.; Yang, G.; Zhou, X.; Hou, X.; Meng, J.; Luan, Y. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate NBS-LRR genes by decoying miR482b in the tomato-Phytophthora infestans interaction. Hortic. Res. 2019, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Cui, J.; Hou, X.; Yang, G.; Xiao, Y.; Han, L.; Meng, J.; Luan, Y. Sl-lncRNA15492 interacts with Sl-miR482a and affects Solanum lycopersicum immunity against Phytophthora infestans. Plant J. 2020, 103, 1561–1574. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cui, J.; Luan, Y. Overexpression of lncRNA08489 enhances tomato immunity against Phytophthora infestans by decoying miR482e-3p. Biochem. Biophys. Res. Commun. 2022, 587, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.K.; Gupta, P.; Sreevathsa, R.; Pradhan, S.K.; Sanjay, T.D.; Mohanty, M.R.; Roul, P.K.; Singh, N.K.; Rai, R. Phylogenomic Analysis of micro-RNA Involved in Juvenile to Flowering-Stage Transition in Photophilic Rice and Its Sister Species. Cells 2023, 12, 1370. [Google Scholar] [CrossRef]
- Dash, P.K.; Gupta, P.; Pradhan, S.K.; Shasany, A.K.; Rai, R. Analysis of homologous regions of small RNAs MIR397 and MIR408 reveals the conservation of microsynteny among rice crop-wild relatives. Cells 2022, 11, 3461. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Barik, S.R.; Dash, P.K.; Lenka, D.; Pradhan, K.C.; Raj, K.R.; Mohanty, S.P.; Monhaty, M.R.; Sahoo, A.; Jena, B.K.; et al. Molecular Breeding for Incorporation of Submergence Tolerance and Durable Bacterial Blight Resistance into the Popular Rice Variety ‘Ranidhan’. Biomolecules 2023, 13, 198. [Google Scholar] [CrossRef]
- Liu, N.; Xu, Y.; Li, Q.; Cao, Y.; Yang, D.; Liu, S.; Wang, X.; Mi, Y.; Liu, Y.; Ding, C. A lncRNA fine-tunes salicylic acid biosynthesis to balance plant immunity and growth. Cell Host Microbe 2022, 30, 1124–1138.e1128. [Google Scholar] [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Zhang, D.; Yin, D.; Zhao, Y.; Ji, C.; Zhao, X.; Li, X.; He, Q.; Chen, R. A novel antisense long noncoding RNA, TWISTED LEAF, maintains leaf blade flattening by regulating its associated sense R2R3-MYB gene in rice. New Phytol. 2018, 218, 774–788. [Google Scholar] [CrossRef]
- Seo, J.S.; Diloknawarit, P.; Park, B.S.; Chua, N.H. ELF18-INDUCED LONG NONCODING RNA 1 evicts fibrillarin from mediator subunit to enhance PATHOGENESIS-RELATED GENE 1 (PR1) expression. New Phytol. 2019, 221, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Jiang, N.; Hou, X.; Wu, S.; Zhang, Q.; Meng, J.; Luan, Y. Genome-wide identification of lncRNAs and analysis of ceRNA networks during tomato resistance to Phytophthora infestans. Phytopathology 2020, 110, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I.K. How to cope with the challenges of environmental stresses in the era of global climate change: An update on ROS stave off in plants. Int. J. Mol. Sci. 2022, 23, 1995. [Google Scholar] [CrossRef] [PubMed]
- Bhar, A.; Gupta, S.; Chatterjee, M.; Sen, S.; Das, S. Differential expressions of photosynthetic genes provide clues to the resistance mechanism during Fusarium oxysporum f. sp. ciceri race 1 (Foc1) infection in chickpea (Cicer arietinum L.). Eur. J. Plant Pathol. 2017, 148, 533–549. [Google Scholar] [CrossRef]
- Thomma, B.P.; Nürnberger, T.; Joosten, M.H. Of PAMPs and effectors: The blurred PTI-ETI dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lnc RNA 16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zeng, Q.; She, L.; Yuan, H.; Luo, Y.; Wang, R.; She, Y.; Wang, W.; Wang, C.; Pan, X. Wolbachia utilizes lncRNAs to activate the anti-dengue toll pathway and balance reactive oxygen species stress in Aedes aegypti through a competitive endogenous RNA network. Front. Cell. Infect. Microbiol. 2022, 11, 1464. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, J.; Deng, F.; Wang, W.; Cheng, Y.; Song, L.; Hu, M.; Shen, J.; Xu, Q.; Shen, F. The long non-coding RNA lncRNA973 is involved in cotton response to salt stress. BMC Plant Biol. 2019, 19, 459. [Google Scholar] [CrossRef]
- Bhatia, G.; Upadhyay, S.K.; Upadhyay, A.; Singh, K. Investigation of long non-coding RNAs as regulatory players of grapevine response to powdery and downy mildew infection. BMC Plant Biol. 2021, 21, 265. [Google Scholar] [CrossRef]
- Zhang, T.; Liang, Q.; Li, C.; Fu, S.; Kundu, J.K.; Zhou, X.; Wu, J. Transcriptome analysis of rice reveals the lncRNA–mRNA regulatory network in response to rice black-streaked dwarf virus infection. Viruses 2020, 12, 951. [Google Scholar] [CrossRef]
- Kang, S.-H.; Sun, Y.-D.; Atallah, O.O.; Huguet-Tapia, J.C.; Noble, J.D.; Folimonova, S.Y. A long non-coding RNA of Citrus tristeza virus: Role in the virus interplay with the host immunity. Viruses 2019, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.-N.; Li, Y.-T.; Wu, Y.-Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.-L. Plant disease resistance-related signaling pathways: Recent progress and future prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, S.; Zhang, Q.; Cui, M.; Zhao, M.; Li, N.; Wang, S.; Wu, R.; Zhang, L.; Cao, Y. The interaction of ABA and ROS in plant growth and stress resistances. Front. Plant Sci. 2022, 13, 1050132. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.J., Jr.; Fichman, Y.; Zandalinas, S.I.; Mittler, R. Jasmonic acid and salicylic acid modulate systemic reactive oxygen species signaling during stress responses. Plant Physiol. 2023, 191, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Bhar, A.; Chatterjee, M.; Gupta, S.; Das, S. Salicylic acid regulates systemic defense signaling in chickpea during Fusarium oxysporum f. sp. ciceri race 1 infection. Plant Mol. Biol. Report. 2018, 36, 162–175. [Google Scholar] [CrossRef]
- Yadav, M.; Pandey, J.; Chakraborty, A.; Hassan, M.I.; Kundu, J.K.; Roy, A.; Singh, I.K.; Singh, A. A comprehensive analysis of calmodulin-like proteins of glycine max indicates their role in calcium signaling and plant defense against insect attack. Front. Plant Sci. 2022, 13, 817950. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Wei, Q.; Hu, Y.; Chen, M.; Chen, Z.; Zheng, S.; Ma, Q.; Luo, Z. Physiological and Full-Length Transcriptome Analyses Reveal the Dwarfing Regulation in Trifoliate Orange (Poncirus trifoliata L.). Plants 2023, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Li, S.; Hu, L.; Zhang, C. Genome-wide analysis of long non-coding RNAs (lncRNAs) in two contrasting rapeseed (Brassica napus L.) genotypes subjected to drought stress and re-watering. BMC Plant Biol. 2020, 20, 81. [Google Scholar] [CrossRef]
- Brant, E.J.; Budak, H. Plant small non-coding RNAs and their roles in biotic stresses. Front. Plant Sci. 2018, 9, 1038. [Google Scholar] [CrossRef]
- Cui, J. lncRNA in plants: Function, mechanisms and applications. Front. Plant Sci. 2023, 14, 1238185. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, Q.-H.; Kaufmann, K. Long non-coding RNAs in plants: Emerging modulators of gene activity in development and stress responses. Planta 2020, 252, 92. [Google Scholar] [CrossRef]


| Sl. No | Name of Plant | Name of Pathogen | Interaction | References |
|---|---|---|---|---|
| lncRNA | ||||
| 1. | Plant | Biotic stress | miRNA–lncRNA interaction | [20] |
| 2. | Mulberry (Morus multicaulis) | Botrytis cinerea and Pseudomonas syringae pv tomato DC3000 | MuLnc1-driven inactivation of calmodulin-like protein gene CML27 | [21] |
| 3. | Cotton (Gossypium hirsutum L.) | Aphid | GhlncRNA149.1 interacts with the CC-NBS-LRR family gene GhA01G0129 as a potential target. | [22] |
| 4. | Arabidopsis thaliana | Phytophthora capsici | nalncFL7 negatively regulates FORKED-LIKE7 (FL7). | [23] |
| 5. | Tomato (Solanum lycopersicum L.) | Phytophthora infestans | lncRNA33732 interaction with WRKY1 | [24] |
| 6. | Wheat (Triticum aestivum) | Blumeria graminis f. sp. tritici | Non-coding RNA profiling | [25] |
| 7. | Cotton (Gossypium hirsutum L.) | Verticillium dahliae and Botrytis cinerea | GhlncNAT-ANX2- and GhlncNAT-RLP7 control the expression of LOX1 and LOX2. | [26] |
| 8. | Wheat (Triticum aestivum) | Rhizoctonia cerealis | MSTRG.4380.1 in growth retardation of fungi | [27] |
| 9. | Rice (Oryza sativa) | Magnaporthae oryzae | Intergenic lncRNA candidates for resistance | [28] |
| 10. | Cotton (Gossypium hirsutum L.) | Verticillium dahliae | Overexpression of lncRNA012077 and down regulation of lncRNA007722 | [29] |
| 11. | Rice (Oryza sativa) | Ustilaginoidea virens | UvlncNAT-MFS, development of smut fungus | [30] |
| 12. | Pumpkin (Cucurbita pepo L.) | Phytophthora xanthii | lncRNA modulates immune responsive pathway, MAPK pathway, and hormonal cross-talk. | [31] |
| 13. | Potato (Solanum tuberosum L.) | Phytophthora infestans | Genome-wide analysis of lncRNA and their interrelationship | [32] |
| 14. | Tomato (Solanum lycopersicum L.) | miRNA–lncRNA interaction, biotic/abiotic stress tolerance | [33] | |
| miPEPs | ||||
| 15. | Plants (Arabidopsis, grapevine, soybean, and Medicago) | - | Interaction network of miPEPs as transcription factor, endocytosis, and transcriptional activator | [34] |
| 16. | Plants | - | Discovery of miPEPs in their probable role in plants and animals | [35] |
| 17. | Plants | - | miPEPs in growth, development, and stress response | [36] |
| 18. | Peanuts (Arachis hypogaea) | - | Mining of miRNA and their potential targets of miPEPs | [37] |
| 19. | Medicago truncatula and Arabidopsis thaliana | - | Finding the role of miPEPs in regulating the expression of miRNA and development of tasi-RNA and phasi-RNA | [38] |
| CircRNA | ||||
| 20. | Legume crops | - | Interaction with DNA, RNA, and protein, modulation of target protein | [39] |
| 21. | Arabidopsis thaliana | Pseudomonas syringae and Botrytis cinerea | circR194 and circR4022 involved in resistance to P. syringae, and circR11208 protecting from B. cinerea | [40] |
| 22. | Plants | - | Bioinformatic mining of circRNA and their potential roles in biotic and abiotic stress factors | [41] |
| 23. | Plants | - | Identification of circRNA in plants with reference to biotic/abiotic stress | [42] |
| 24. | Plants | - | Post-transcriptional modification of gene expression | [43] |
| 25. | Tea plant (Camellia sinensis) | Helopeltis theivora | Activation of secondary metabolites, endogenous target mimics (eTMs) of target genes, e.g., aspartyl protease, phospholipase, lectin receptor, etc. | [44] |
| 26. | Tomato (Solanum lycopersicum L.) | Planticine®-induced defense responses | Upregulation of circRNA whitefly-induced gp91 | [45] |
| 27. | Tomato (Solanum lycopersicum L.) | Phytophthora infestans | circRNA45 and circRNA47, positive regulators of resistance response in tomato | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhar, A.; Roy, A. Emphasizing the Role of Long Non-Coding RNAs (lncRNA), Circular RNA (circRNA), and Micropeptides (miPs) in Plant Biotic Stress Tolerance. Plants 2023, 12, 3951. https://doi.org/10.3390/plants12233951
Bhar A, Roy A. Emphasizing the Role of Long Non-Coding RNAs (lncRNA), Circular RNA (circRNA), and Micropeptides (miPs) in Plant Biotic Stress Tolerance. Plants. 2023; 12(23):3951. https://doi.org/10.3390/plants12233951
Chicago/Turabian StyleBhar, Anirban, and Amit Roy. 2023. "Emphasizing the Role of Long Non-Coding RNAs (lncRNA), Circular RNA (circRNA), and Micropeptides (miPs) in Plant Biotic Stress Tolerance" Plants 12, no. 23: 3951. https://doi.org/10.3390/plants12233951