
Detailed Seed Cone Morpho-Anatomy Provides New Insights into Seed Cone Origin and Evolution of Podocarpaceae; Podocarpoid and Dacrydioid Clades

Abstract
:1. Introduction
2. Material and Methods
2.1. Seed Cone Collection
2.2. Morphology and Distribution of the Investigated Taxa
- The South African Afrocarpus falcatus (Thunb.) C.N. Page, which forms large trees [21].
- Two species of Nageia, N. nagi (Thunb.) Kuntze, which is an evergreen tall tree, distributed in China (introduced), Japan, Taiwan (introduced) and Vietnam [21,22] and N. wallichiana (Presl.) Kuntze, which is a tall tree distributed in Brunei Darussalam, Cambodia, China, India, Indonesia, Laos, Malaysia, Myanmar, Papua New Guinea, the Philippines, Thailand and Vietnam [21,22]).
- Three species of Podocarpus subgenus Podocarpus (P. henkelii Stapf., P. elongatus (Aiton) L’Herit. Ex Persoon and P. oleifolius D. Don). Podocarpus henkelii and P. elongatus occur in southern Africa, while P. oleifolius is found in Central America [23].
- Two species of Podocarpus subgenus Foliolatus (P. spinulosus (Smith) R. Br. Ex Mirbel and P. elatus R. Br. ex Endlicher). Podocarpus spinulosus and P. elatus are dioecious species from northeastern Australia [24].
- Retrophyllum comptonii (Buchholz) C.N. Page, which is a tree endemic to New Caledonia [25].
- Dacrycarpus dacrydioides (A. Richard.) de Laub., a tall tree endemic to New Zealand.
- Dacrydium cupressinum Solander ex G. Forst., a tall tree endemic to New Zealand.
- Falcatifolium papuanum de Laub., a tall tree endemic to Papua New Guinea [26].
2.3. Taxon Processing and Sectioning
2.4. Measurements and Trait Reconstructions
3. Results
3.1. Afrocarpus Seed Cone Morpho-Anatomy
3.2. Nageia Seed Cone Morpho-Anatomy
3.3. Podocarpus Seed Cone Morpho-Anatomy
3.3.1. Podocarpus henkelii
3.3.2. Podocarpus elongatus
3.3.3. Podocarpus oleifolius
3.3.4. Podocarpus spinulosus
3.3.5. Podocarpus elatus
3.4. Retrophyllum Seed Cone Morpho-Anatomy
3.5. Acmopyle Seed Cone Morpho-Anatomy
3.6. Dacrycarpus Seed Cone Morpho-Anatomy
3.7. Dacrydium Seed Cone Morpho-Anatomy
3.8. Falcatifolium Seed Cone Morpho-Anatomy
3.9. Comparison of Seed Cone Morpho-Anatomical Characters
3.10. Seed Cone Morpho-Anatomical Traits and Structures
3.10.1. Fleshy Seed Cones
3.10.2. Epimatium Morphology
3.10.3. Receptaculum Morphology
3.10.4. Testa Morpho-Anatomy
3.10.5. Ovule and Embryo Traits
4. Discussion
4.1. The Dominance of Fleshy Seed Cones in Living Podocarps and Fossil Evidence
4.2. Shifts in the Fleshy Structures
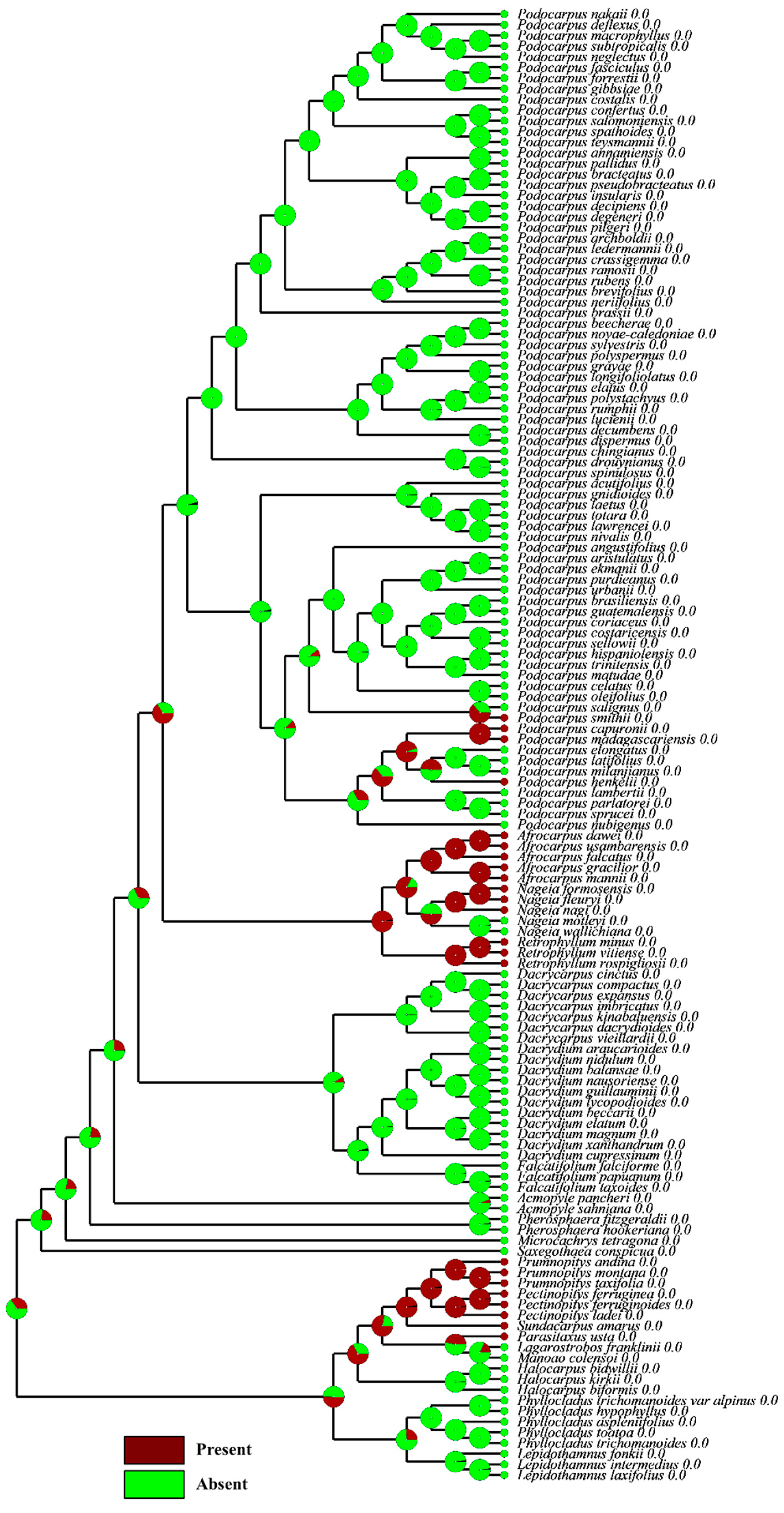
4.3. Significance of the Evolutionary Reconstruction of Fleshiness
4.4. Broader Perspective with other Podocarps
4.5. Evolution of Morphological Structures and Animal Dispersal
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Brodribb, T.J. A functional analysis of podocarp ecology. In Smithsonian Contributions to Botany; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011. [Google Scholar]
- Biffin, E.; Conran, J.G.; Lowe, A.J. Podocarp evolution: A molecular phylogenetic perspective. In Ecology of the Podocarpaceae in tropical Forests; Turner, B.L., Cernusak, L.A., Eds.; Smithsonian Contributions to Botany; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011. [Google Scholar]
- Biffin, E.; Brodribb, T.J.; Hill, R.S.; Thomas, P.; Lowe, A.J. Leaf evolution in Southern Hemisphere conifers tracks the angiosperm ecological radiation. Proc. R. Soc. B Biol. Sci. 2012, 279, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Leslie, A.B.; Beaulieu, J.; Holman, G.; Campbell, C.S.; Mei, W.; Raubeson, L.R.; Mathews, S. An overview of extant conifer evolution from the perspective of the fossil record. Am. J. Bot. 2018, 105, 1531–1544. [Google Scholar] [CrossRef]
- Leslie, A.B.; Benson, R.B.J. Neontological and paleontological congruence in the evolution of Podocarpaceae (coniferales) reproductive morphology. Front. Ecol. Evol. 2022, 10, 1058746. [Google Scholar] [CrossRef]
- Klaus, K.V.; Matzke, N.J. Statistical comparison of trait-dependent biogeographical models indicates that Podocarpaceae dispersal is influenced by both seed cone traits and geographical distance. Syst. Biol. 2020, 69, 61–75. [Google Scholar] [CrossRef]
- Rudall, P. Colourful cones: How did flower colour first evolve? J. Exp. Bot. 2020, 71, 759. [Google Scholar] [CrossRef]
- Herting, J.; Stützel, T.; Klaus, K.V. The Ancestral Conifer Cone: What Did It Look Like? A Modern Trait-Evolution Approach. Int. J. Plant Sci. 2020, 181, 871–886. [Google Scholar] [CrossRef]
- Khan, R.; Hill, R.S. Morpho-anatomical affinities and evolutionary relationships of three paleoendemic podocarp genera based on seed cone traits. Ann. Bot. 2021, 128, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Hill, R.S.; Dörken, V.M.; Biffin, E. Detailed seed cone morpho-anatomy of the Prumnopityoid clade: An insight into the origin and evolution of Podocarpaceae seed cones. Ann. Bot. 2022, 130, 637–655. [Google Scholar] [CrossRef]
- Andruchow-Colombo, A.; Wilf, P.; Escapa, I.H. A South American fossil relative of Phyllocladus: Huncocladus laubenfelsii gen. et sp. nov. (Podocarpaceae), from the early Eocene of Laguna del Hunco, Patagonia, Argentina. Aust. Syst. Bot. 2019, 32, 290–309. [Google Scholar] [CrossRef]
- Morley, R.J. Dispersal and paleoecology of tropical podocarps. In Smithsonian Contributions to Botany; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011; pp. 21–41. [Google Scholar]
- Khan, R.; Hill, R.S.; Liu, J.; Biffin, E. Diversity, Distribution, Systematics and Conservation Status of Podocarpaceae. Plants 2023, 12, 1171. [Google Scholar] [CrossRef]
- Tomlinson, P. Aspects of cone morphology and development in Podocarpaceae (Coniferales). Int. J. Plant Sci. 1992, 153, 572–588. [Google Scholar] [CrossRef]
- Quiroga, M.P.; Mathiasen, P.; Iglesias, A.; Mill, R.R.; Premoli, A.C. Molecular and fossil evidence disentangle the biogeographical history of Podocarpus, a key genus in plant geography. J. Biogeogr. 2016, 43, 372–383. [Google Scholar] [CrossRef]
- Mundry, I. Morphologische und morphogenetische Untersuchungen zur-Evolution der Gymnospermen. Bibl. Bot. 2000, 152, 1–90. [Google Scholar]
- Restemeyer, J. Morphologische und morphogenetische Untersuchungen zur Phylogenie und Evolution der Podocarpaceae und Phyllocladaceae. Ph.D. Thesis, Ruhr-Universität Bochum, Bochum, Germany, 2002. [Google Scholar]
- Mill, R.R.; Möller, M.; Glidewell, S.M.; Masson, D.; Williamson, B. Comparative anatomy and morphology of fertile complexes of Prumnopitys and Afrocarpus species (Podocarpaceae) as revealed by histology and NMR imaging, and their relevance to systematics. Bot. J. Linn. Soc. 2004, 145, 295–316. [Google Scholar] [CrossRef]
- Chen, L.; Jin, W.-T.; Liu, X.-Q.; Wang, X.-Q. New insights into the phylogeny and evolution of Podocarpaceae inferred from transcriptomic data. Mol. Phylogenetics Evol. 2022, 166, 107341. [Google Scholar] [CrossRef]
- Leslie, A.B. How many ways can you build a conifer cone? A commentary on ‘Origin and evolution of Podocarpaceae seed cones’. Ann. Bot. 2022, 130, i–iii. [Google Scholar] [CrossRef]
- Farjon, A. The Kew review: Conifers of the world. Kew Bull. 2018, 73, 8. [Google Scholar] [CrossRef]
- Farjon, A. Taxonomic treatment of genera and species. A Handbook of the World’s Conifers; Brill: Leiden, The Netherlands, 2017; Volume 2. [Google Scholar]
- Ornelas, J.F.; Ortiz-Rodriguez, A.E.; Ruiz-Sanchez, E.; Sosa, V.; Pérez-Farrera, M.Á. Ups and downs: Genetic differentiation among populations of the Podocarpus (Podocarpaceae) species in Mesoamerica. Mol. Phylogenetics Evol. 2019, 138, 17–30. [Google Scholar] [CrossRef]
- Mellick, R.; Rossetto, M.; Porter, C. Isolation and characterisation of polymorphic microsatellite loci from Podocarpus elatus (Podocarpaceae). Mol. Ecol. Resour. 2009, 9, 1460–1466. [Google Scholar]
- Mill, R. A monographic revision of Retrophyllum (Podocarpaceae). Edinb. J. Bot. 2016, 73, 171–261. [Google Scholar] [CrossRef]
- De Laubenfels David, J. Coniferales. In Flora Malesiana; Kluwer Academic: Dordrecht, The Netherlands, 1988; Series I; Volume 10. [Google Scholar]
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers: An Analysis of Their Distribution, Biogeography, Diversity and Conservation Status; Brill: Leiden, The Netherlands, 2013. [Google Scholar]
- Yu, Y.; Blair, C.; He, X. RASP 4: Ancestral state reconstruction tool for multiple genes and characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.; Maddison, D. Mesquite: A Modular System for Evolutionary Analysis, Version 3.51; Mesquite Project: Arlington, AZ, USA, 2019. [Google Scholar]
- Buchholz, J.T. Embryogeny of the Podocarpaceae. Bot. Gaz. 1941, 103, 1–37. [Google Scholar] [CrossRef]
- Doyle, J.; Looby, W. Embryogeny in Saxegothaea and its relation to other podocarps. Sci. Proc. R. Dublin Soc. 1939, 22, 127–147. [Google Scholar]
- Peters, M.D. A taxonomic Analysis of a Middle Cretaceous Megafossil Plant Assemblage from Queensland, Australia. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 1985. [Google Scholar]
- Hill, R.; Carpenter, R. Evolution of Acmopyle and Dacrycarpus (Podocarpaceae) foliage as inferred from macrofossils in south-eastern Australia. Aust. Syst. Bot. 1991, 4, 449–479. [Google Scholar] [CrossRef]
- Mittre, V. Studies on the fossil flora of Nipania, Rajmahal Series, India—Pteridophyta and general observations on Nipania fossil flora. Palaeobotanist 1959, 7, 47–66. [Google Scholar]
- Rao, A. Nipaniostrobus, a New Genus of Dacrydium-Like Sed-Bearing Cones, and Other Silicifield Plants from the Rajmahal Series. Proc. Natl. Acad. Sci. USA 1943, 13, 113–133. [Google Scholar]
- Townrow, J.A. Conifer from the Jurassic of east Antarctica. Pap. Proc. R. Soc. Tasman. 1967, 101, 137–149. [Google Scholar]
- Miller, C.N. Mesozoic conifers. Bot. Rev. 1977, 43, 217–280. [Google Scholar] [CrossRef]
- Reymanówna, M. A Jurassic podocarp from Poland. Rev. Palaeobot. Palynol. 1987, 51, 133–143. [Google Scholar] [CrossRef]
- Stewart, W.N.; Stewart, W.N.; Stewart, W.M.; Rothwell, G.W. Paleobotany and the Evolution of Plants; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Wu, X.K.; Zavialova, N.E.; Kodrul, T.M.; Liu, X.Y.; Gordenko, N.V.; Maslova, N.P.; Quan, C.; Jin, J.H. Northern Hemisphere megafossil of Dacrycarpus (Podocarpaceae) from the Miocene of South China and its evolutionary and paleoecological implications. J. Syst. Evol. 2021, 59, 352–374. [Google Scholar] [CrossRef]
- Wilf, P. Rainforest conifers of Eocene Patagonia: Attached cones and foliage of the extant Southeast Asian and Australasian genus Dacrycarpus (Podocarpaceae). Am. J. Bot. 2012, 99, 562–584. [Google Scholar] [CrossRef]
- Lewis, E.K.; Drinnan, A.N. The Miocene conifer flora of Balcombe Bay, Victoria, Australia. Aust. Syst. Bot. 2013, 26, 145–155. [Google Scholar] [CrossRef]
- Geldenhuys, C. Reproductive biology and population structures of Podocarpus falcatus and P. latifolius in southern Cape forests. Bot. J. Linn. Soc. 1993, 112, 59–74. [Google Scholar] [CrossRef]
- Mill, R. A new sectional combination in Nageia Gaertn. (Podocarpaceae). Edinb. J. Bot. 2001, 58, 499. [Google Scholar] [CrossRef]
- Contreras, D.; Duijnstee, I.; Ranks, S.; Marshall, C.; Looy, C. Evolution of dispersal strategies in conifers: Functional divergence and convergence in the morphology of diaspores. Perspect. Plant Ecol. Evol. Syst. 2017, 24, 93–117. [Google Scholar] [CrossRef]
- Sahni, B.; Mitra, A. Notes on the anatomy of some New Zealand species of Dacrydium. Ann. Bot. 1927, 41, 75–89. [Google Scholar] [CrossRef]
- Herting, J.; Stützel, T. Morphogenesis of the seed cone of Araucaria ancheri (Molina) K. Koch and the evolution of the coniferous seed scale. Flora 2020, 273, 151719. [Google Scholar] [CrossRef]
- Field, T.S.; Brodribb, T.J.; Iglesias, A.; Chatelet, D.S.; Baresch, A.; Upchurch, G.R.; Gomez, B.; Mohr, B.A.; Coiffard, C.; Kvacek, J. Fossil evidence for Cretaceous escalation in angiosperm leaf vein evolution. Proc. Natl. Acad. Sci. USA 2011, 108, 8363–8366. [Google Scholar] [CrossRef] [PubMed]
- Galetti, M.; Pizo, M.A.; Morellato, L.P.C. Diversity of functional traits of fleshy fruits in a species-rich Atlantic rain forest. Biota Neotrop. 2011, 11, 181–193. [Google Scholar] [CrossRef]
- Leslie, A.B.; Beaulieu, J.M.; Mathews, S. Variation in seed size is structured by dispersal syndrome and cone morphology in conifers and other nonflowering seed plants. New Phytol. 2017, 216, 429–437. [Google Scholar] [CrossRef]
- Del Fueyo, G.M. Cone and ovule development in the Podocarpus species from Argentina. Phytomorphology 1999, 49, 49–60. [Google Scholar]
- Mill, R.; Möller, M.; Christie, F.; Glidewell, S.; Masson, D.; Williamson, B. Morphology, anatomy and ontogeny of female cones in Acmoanchericheri (Brongn. & Gris) Pilg. (Podocarpaceae). Ann. Bot. 2001, 88, 55–67. [Google Scholar]
- Blendinger, P.G. Functional equivalence in seed dispersal effectiveness of Podocarpus parlatorei in Andean fruit-eating bird assemblages. Front. Ecol. Evol. 2017, 5, 57. [Google Scholar] [CrossRef]
- Enright, N.J.; Jaffré, T. Ecology and distribution of the Malesian podocarps. In Ecology of the Podocarpaceae in Tropical Forests; Turner, B.L., Cernusak, L.A., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011. [Google Scholar]
- Koen, J. The effect of rodent granivory on recruitment of the irregularly fruiting Podocarpus falcatus in the southern Cape. S. Afr. For. J. 1991, 159, 25–28. [Google Scholar] [CrossRef]
- Eckenwalder, J.E. Conifers of the World: The Complete Reference; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Wilf, P.; Donovan, M.P.; Cúneo, N.R.; Gandolfo, M.A. The fossil flip-leaves (Retrophyllum, Podocarpaceae) of southern South America. Am. J. Bot. 2017, 104, 1344–1369. [Google Scholar] [CrossRef]
- Fountain, D.W.; Holdsworth, J.M.; Outred, H.A. The dispersal unit of Dacrycarpus dacrydioides (A. Rich.) de Laubenfels (Podocarpaceae) and the significance of the fleshy receptacle. Bot. J. Linn. Soc. 1989, 99, 197–207. [Google Scholar] [CrossRef]
- Robertson, H.A.; Hackwell, K.R. Habitat preferences of birds in seral kahikatea Dacrycarpus dacrydioides (Podocarpaceae) forest of South Westland, New Zealand. Biol. Conserv. 1995, 71, 275–280. [Google Scholar] [CrossRef]
- Norton, D.; Herbert, J.; Beveridge, A. The ecology of Dacrydium cupressinum: A review. N. Z. J. Bot. 1988, 26, 37–62. [Google Scholar] [CrossRef]
- Beveridge, A. Dispersal and destruction of seed in central North Island podocarp forests. N. Z. Ecol. Soc. 1964, 11, 48–55. [Google Scholar]
- Beveridge, A. Regeneration of podocarps in a central North Island Forest. N. Z. J. For. 1973, 18, 23–35. [Google Scholar]
- Warburton, B.; Kingsford, S.; Lewitt, D.; Spurr, E. Plant species preferences of birds in lowland rimu (Dacrydium cupressinum) forest-implications for selective-logging. N. Z. J. Ecol. 1992, 16, 119–126. [Google Scholar]
- Fidler, A.E.; Lawrence, S.B.; McNatty, K.P. An hypothesis to explain the linkage between kakapo (Strigops habroptilus) breeding and the mast fruiting of their food trees. Wildl. Res. 2008, 35, 1–7. [Google Scholar] [CrossRef]
- Thorsen, M.J.; Dickinson, K.J.; Seddon, P.J. Seed dispersal systems in the New Zealand flora. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 285–309. [Google Scholar] [CrossRef]
- Spurr, E.; Rod, S.; Tranter, K. Food preferences of the bellbird (Anthornis melanura) in native forest remnants on the Port Hills, Banks Peninsula, New Zealand. Notornis 2011, 58, 139–157. [Google Scholar]









{kind=link}
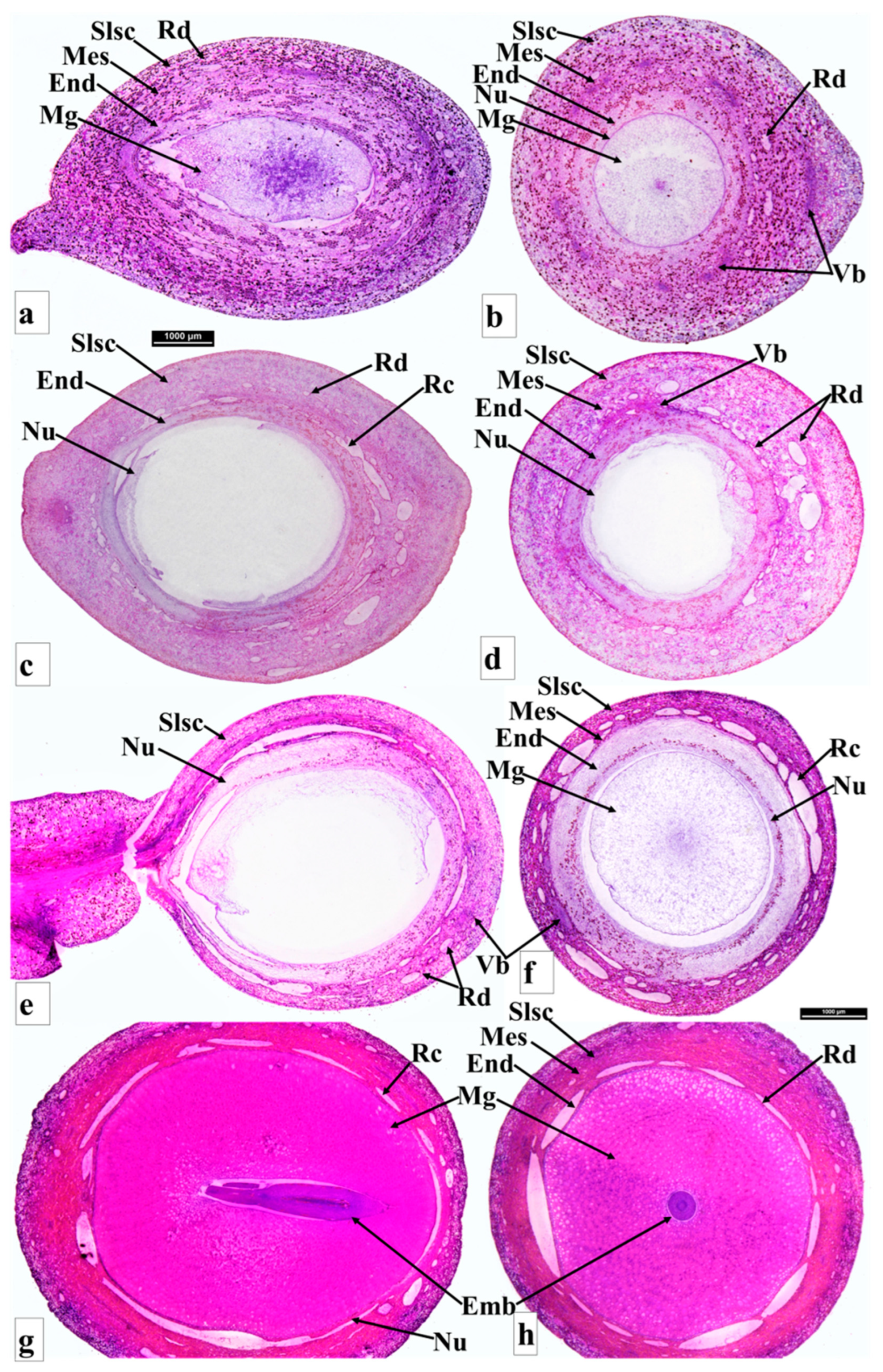{kind=link}
{kind=link}
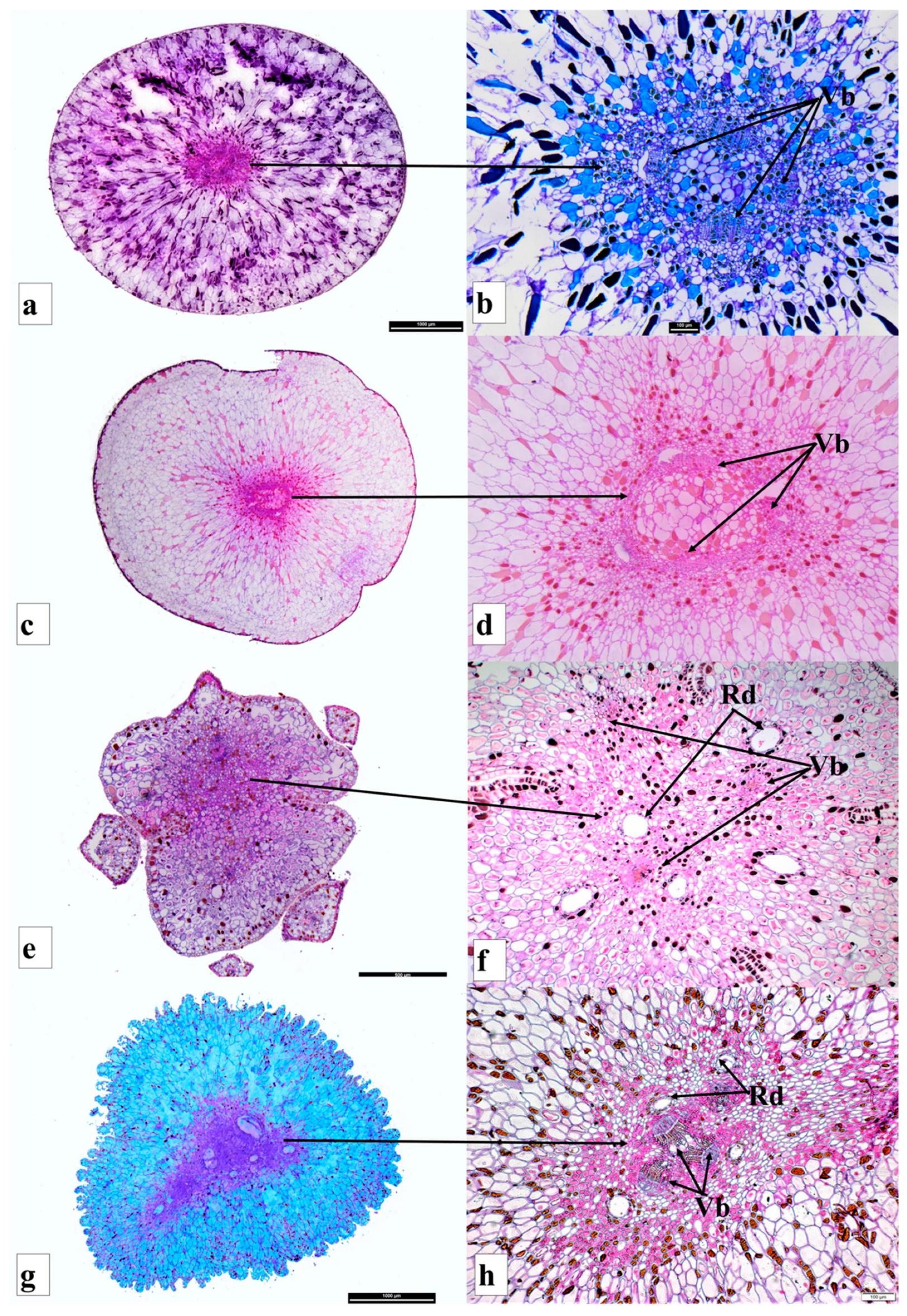{kind=link}
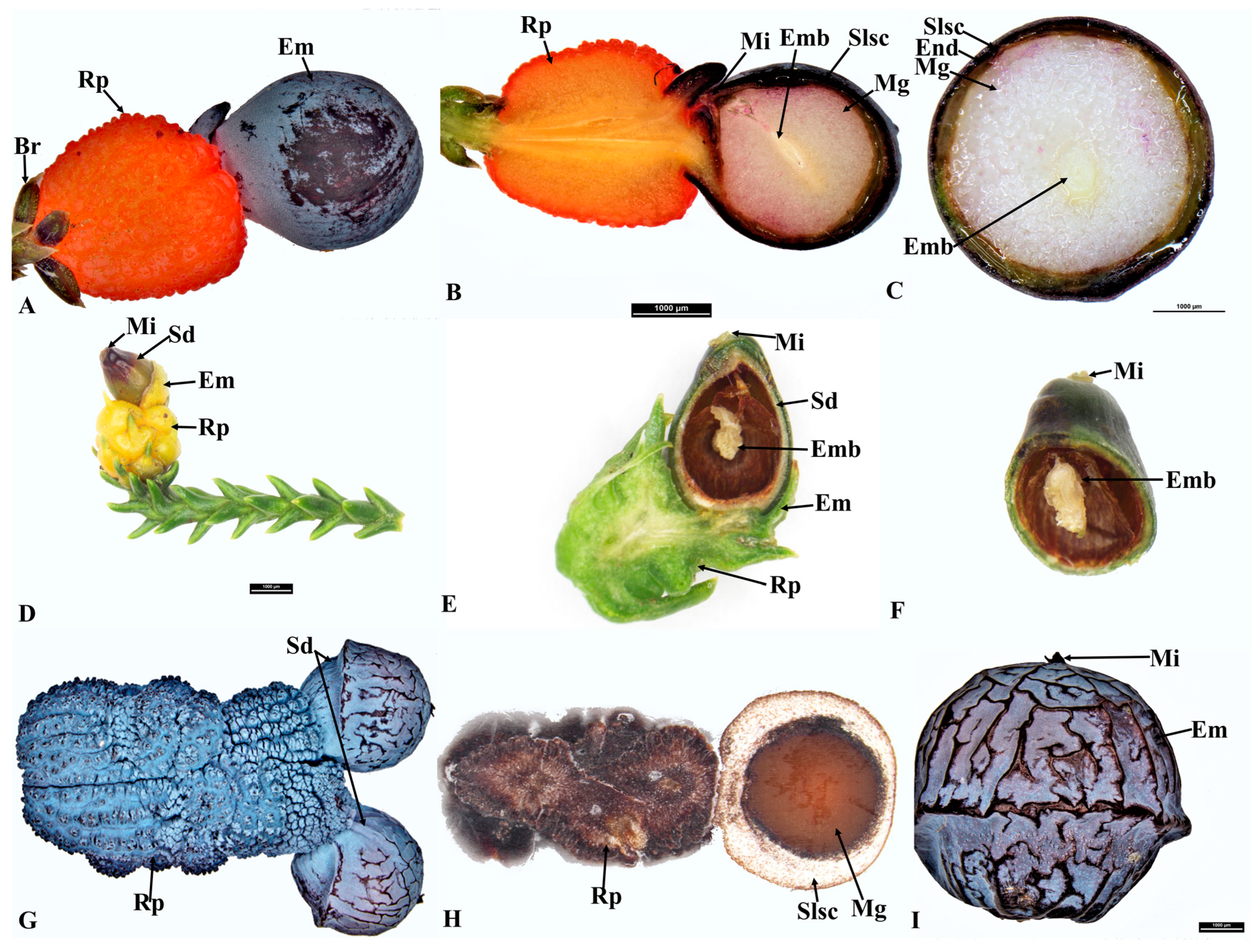{kind=link}
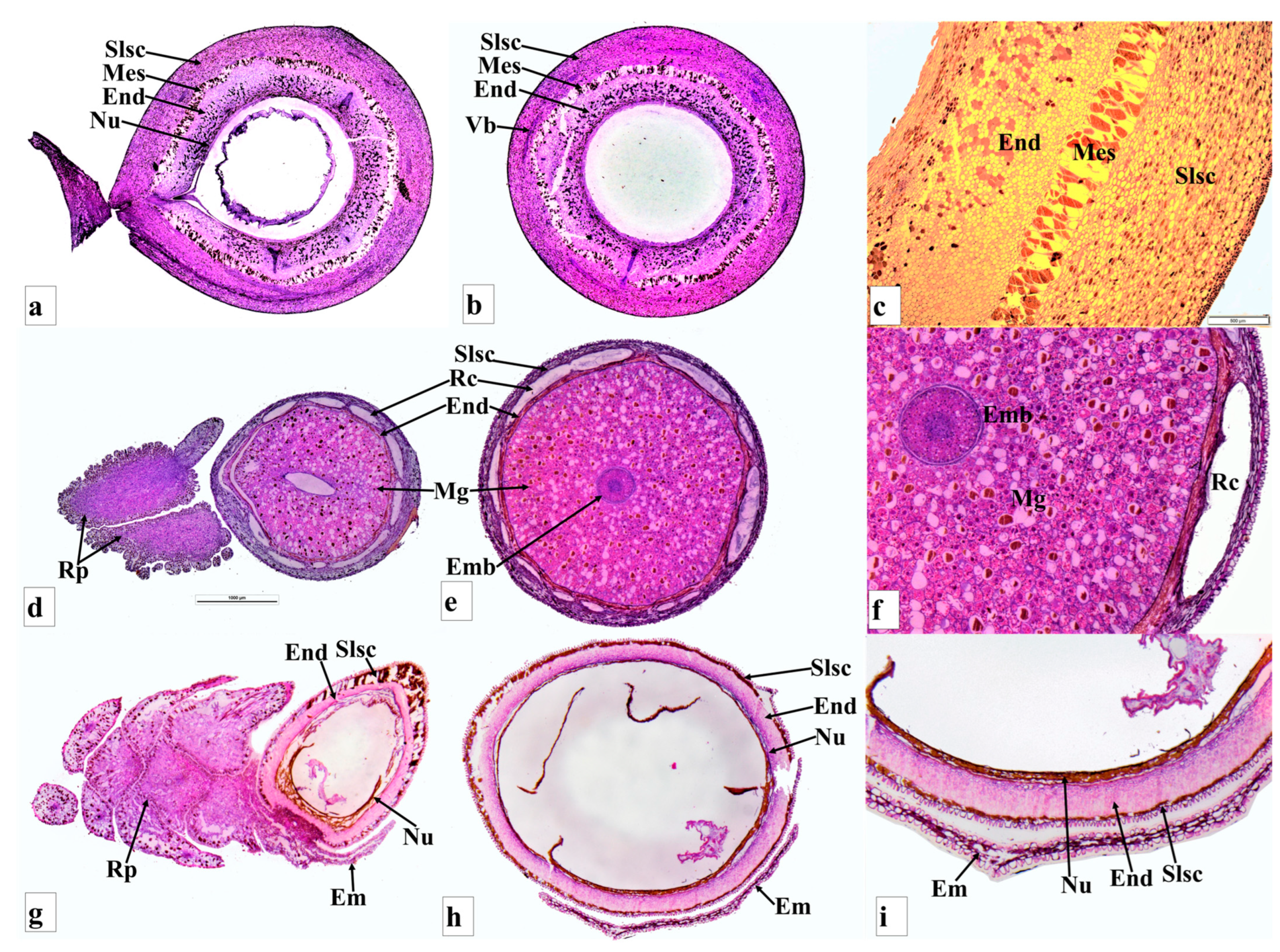{kind=link}
{kind=link}
{kind=link}
| Characters | Afrocarpus falcatus | Nageia wallichiana | Nageia nagi | Podocarpus henkelii (subgenus Podocarpus) | Podocarpus elongatus (subgenus Podocarpus) | Podocarpus oleifolius (subgenus Podocarpus) | Podocarpus spinulosus (subgenus Foliolatus) | Podocarpus elatus (subgenus Foliolatus) | Retrophyllum comptonii | Dacrydium cupressinum | Dacrycarpus dacrydioides | Falcatifolium papuanum | Acmopyle pancheri |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Reproductive cycle | 1 year | 1 year | 1 year | 1 year | 1 year | 1 year | 1 year | 1 year | 1 year | 2 years | 1 year | 1 year | 1–2 years |
| Cone shape | obovoid | ovoid | globose | obovoid | ovoid | ovoid | ovoid | ovoid | ovoid-subglobose | obovoid | ovoid | obovoid | ovoid |
| Cone size (mm) | 12–20 × 8–16 | 18–38 × 6–10 | 12–18 × 10–16 | 15–20 × 8–12 | 13–20 × 4–8 | 12–22 × 4–7 | 11–24 × 6–7 | 25–45 × 10–20 | 10–20 × 7–15 | 6–10 × 2–4 | 10–18 × 4–8 | 10–16 × 4–6 | 18–34 × 10–18 |
| Colour | yellowish | reddish | brown | yellowish | reddish | reddish | dark purple | dark purple | reddish | reddish | reddish | reddish | reddish-brown |
| Number of seeds per cone | 1 | 1–2 | 1 | 1 | 1 | 1–2 | 1 | 1–2 | 1–2 | 1 | 1–2 | 1 | 1–2 |
| Seed size (mm) | 8–18 × 6–14 | 14–20 × 10–16 | 10–15 × 8–12 | 12–17 × 4–6 | 6–12 × 3–5 | 5–8 × 3–6 | 8–10 × 5–7 | 15–20 × 12–15 | 7–17 × 5–12 | 2.5–6 × 2–3 | 6–10 × 4–8 | 6–7 × 4–5 | 6–8 × 5–7 |
| Seed surface | rugose | rugose | rugose | rugose | smooth | smooth | rugose | rugose | rugose | smooth | smooth | smooth | rugose |
| Seed colour | brown | brown | brown | light brown | brown | brown | brown | purplish black | brown | brownish black | purplish black | dark brown | brown |
| Ovule orientation | inverted | inverted | inverted | inverted | inverted | inverted | inverted | inverted | inverted | inverted | inverted | inverted | erect |
| Cuticle | thick | thick | thick | thick | thin | thin | thin | thin | thick | thick | thin | thin | thick |
| Epidermal layers | 1 | 1–2 | 1–2 | 1 | 1 | 1 | 1–2 | 1 | 1 | 1 | 1 | 1 | 1 |
| Epidermal cell shape | rectangular | round–triangular | round–triangular | round–triangular | round–triangular | rectangular | round–triangular | round–triangular | isodiametric | dome-shaped | dome-shaped | dome-shaped | triangular |
| Exotesta | fleshy | coriaceous | coriaceous | fleshy | coriaceous | coriaceous | coriaceous | coriaceous | fleshy | coriaceous | coriaceous | coriaceous | coriaceous |
| Exotesta layers of cells | 10–18 | 14–22 | 12–20 | 12–18 | 8–13 | 10–14 | 12–16 | 18–26 | 16–20 | 1–2 | 10–14 | 3–6 | 14–20 |
| Mesotesta layers of cells | 3–5 | – | – | 16–22 | 3–5 | 10–16 | – | – | 6–8 | – | – | – | 6–12 |
| Endotesta layers of cells | 20–32 | 12–20 | 10–18 | 12–20 | 6–12 | 10–16 | 5–8 | 4–6 | 12–18 | 8–10 | 4–6 | 6–14 | 12–18 |
| Nucellus layers | 4–10 | 5–10 | 6–12 | 6–12 | 8–14 | 7–10 | 6–8 | 2–4 | 4–8 | 6–10 | 3–6 | 4–8 | 4–8 |
| Embryo (megagametophyte) shape | straight | straight | straight | straight | straight | straight | straight | straight | straight | straight | straight | straight | straight |
| Embryo size (mm) | 0.8–1.2 × 0.3–0.5 | 0.8–1.7 × 0.3–0.5 | 0.8–1.5 × 0.3–0.4 | 0.65–1.2 × 0.2–0.4 | 0.5–0.9 × 0.2–0.3 | 0.6–0.8 × 0.2–0.3 | 0.5–1.3 × 0.3–0.5 | 0.7–1.2 × 0.3–0.5 | 0.4–0.9 × 0.2–0.4 | 0.25–0.6 × 0.1–0.2 | 0.6–1.0 × 0.2–0.5 | 0.3–0.6 × 0.15–0.25 | 0.3–6 × 0.1–0.2 |
| Bracts | 1–2 | 4–7 | 4–6 | 1–2 | 2 | 2 | 2–4 | 2 | 3–5 | 8–14 | 2–3 | 6–12 | 4–10 |
| Stomata on bracts | present | present | present | present | present | present | present | present | present | present | present | present | present |
| Epimatium | present | present | present | present | present | present | present | present | present | present | present | present | present |
| Epimatium shape | fleshy and fused with outer testa surrounding the whole seed | papery and fused with outer testa surrounding the whole seed | fleshy and fused with outer testa surrounding the whole seed | fleshy and fused with outer testa surrounding the whole seed | papery and fused with outer testa surrounding the whole seed | papery and fused with outer testa surrounding the whole seed | papery and fused with outer testa surrounding the whole seed | papery and fused with outer testa surrounding the whole seed | fleshy and fused with outer testa surrounding the whole seed | fleshy asymmetrical cup-like | papery and fused with outer testa surrounding the whole seed | fleshy asymmetrical cup-like | papery and fused with outer testa surrounding the whole seed |
| Epimatium structure | fleshy | fleshy | fleshy | fleshy | papery | papery | papery | papery | fleshy | fleshy | papery | fleshy | papery |
| Epimatium colour | yellowish | purple | purple | yellowish | purple | olive green | dark purple | purplish black | reddish | brownish black | purplish black | reddish brown | brown |
| Receptaculum | absent | present | absent | absent | present | present | present | present | absent | present | present | present | present |
| Receptaculum colour | - | reddish | - | absent | bright red | yellowish red | blueish black | blueish black | - | reddish | orange–red | reddish | reddish brown |
| Sclereids | present | present | present | present | present | present | present | present | present | present | present | present | present |
| Resin canals | present | present | present | present | present | present | present | present | present | present | present | present | present |
| Dispersal | zoochory [birds, bats (Rousettus aegyptiacus), monkeys (Chlorocebus pygerythrus), woodland dormouse (Graphiurus murinus) and Verreaux’s mouse (Myomyscus verreauxii)] | zoochory | zoochory | zoochory (birds) | zoochory (birds) | zoochory (birds) | zoochory (birds) | zoochory (birds) | zoochory | zoochory [New Zealand bellbird (Anthornis melanura) and Tūī (Prosthemadera novaeseelandiae), Kākāpō (Strigops habroptilus)] | zoochory [New Zealand pigeon (Hemiphaga novaeseelandiae), Tūī (Prosthemadera novaeseelandiae) and New Zealand bellbird (Anthornis melanura), Anthornis melanura, Hemiphaga novaeseelandiae, Prosthemadera novaeseelandiae] | zoochory (birds) | zoochory (birds) |
| Taxon | Number of Prosuspensor Cells | Number of Binucleate Embryonic Cells | Reference |
|---|---|---|---|
| Podocarpoid Clade | |||
| Afrocarpus | 18–23 | 9–11 | Buchholz, 1941 [30] |
| Nageia | 18–23 | 7, 9–11 | # |
| Podocarpus glomeratus | 11–13 | 2–3 | # |
| P. chinensis | 14–15 | 1–2 | # |
| P. lawrencei | 7–10 | 1–2 | # |
| P. nivalis | 7–10 | 1–2 | # |
| P. totara | 7–10 | 1–2 | # |
| P. laetus | 7–10 | 1–2 | # |
| P. urbanii | 11–13 | 2–3 | # |
| Dacrydioid Clade | |||
| Dacrycarpus dacrydioides | 11–14 | 4 or 5 | Buchholz, 1941 [30] |
| Dacrydium | 7–11 | 5–9 | # |
| Other Podocarpaceae | |||
| Phyllocladus | 4–6 | 9–12 | Doyle and Looby, 1939 [31] |
| Saxegothaea | 3–4 | 10–12 | # |
| Pectinopitys ferruginea | 7–9 | 7–9 | Buchholz, 1941 [30] |
| Prumnopitys taxifolia | 6–9 | 9–12 | # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, R.; Hill, R.S.; Dörken, V.M.; Biffin, E. Detailed Seed Cone Morpho-Anatomy Provides New Insights into Seed Cone Origin and Evolution of Podocarpaceae; Podocarpoid and Dacrydioid Clades. Plants 2023, 12, 3903. https://doi.org/10.3390/plants12223903
Khan R, Hill RS, Dörken VM, Biffin E. Detailed Seed Cone Morpho-Anatomy Provides New Insights into Seed Cone Origin and Evolution of Podocarpaceae; Podocarpoid and Dacrydioid Clades. Plants. 2023; 12(22):3903. https://doi.org/10.3390/plants12223903
Chicago/Turabian StyleKhan, Raees, Robert S. Hill, Veit M. Dörken, and Ed Biffin. 2023. "Detailed Seed Cone Morpho-Anatomy Provides New Insights into Seed Cone Origin and Evolution of Podocarpaceae; Podocarpoid and Dacrydioid Clades" Plants 12, no. 22: 3903. https://doi.org/10.3390/plants12223903