
Relationships between Phyllosphere Bacterial Communities and Leaf Functional Traits in a Temperate Forest
 , , ,
, , , 
Abstract
:1. Introduction
2. Results
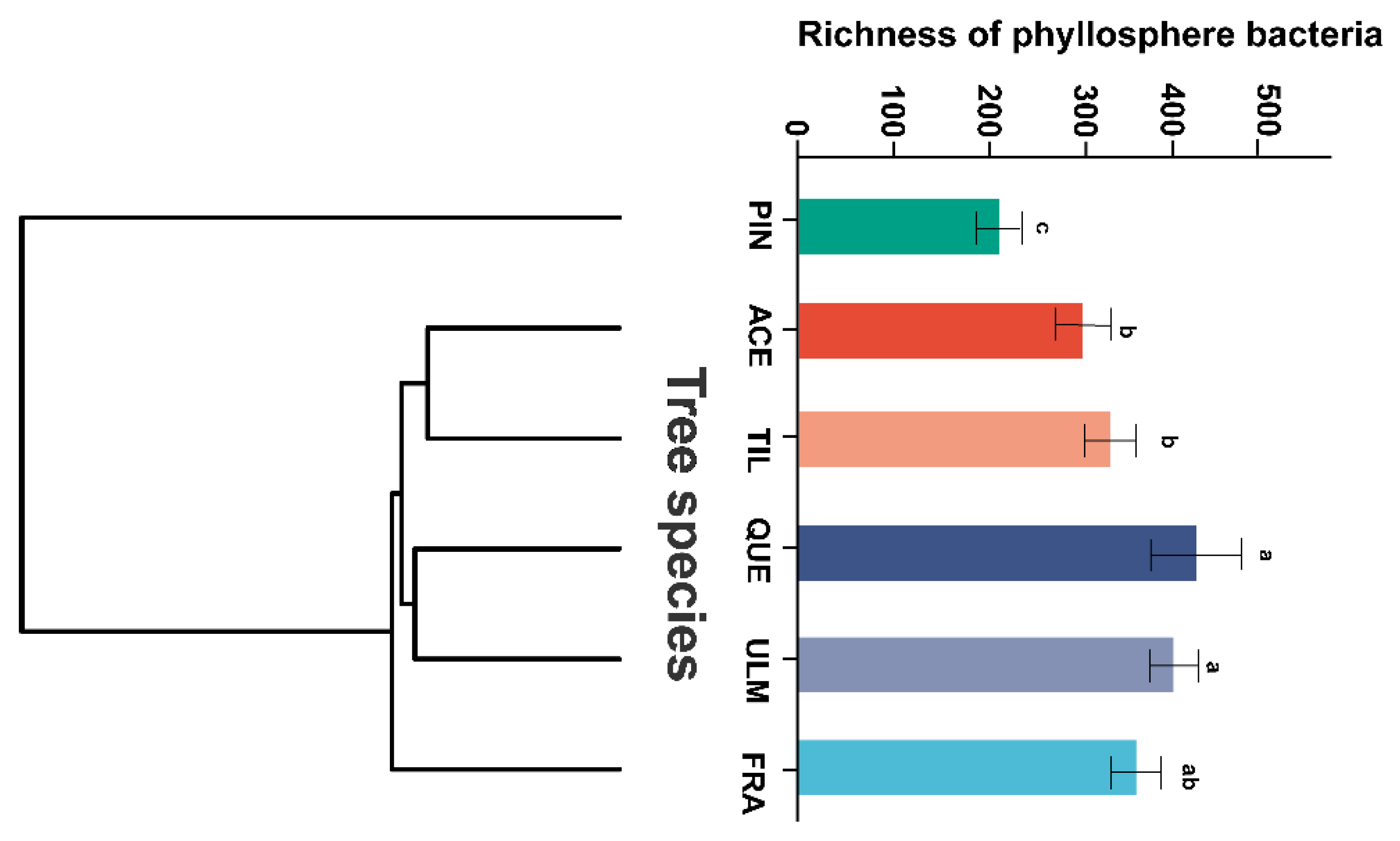
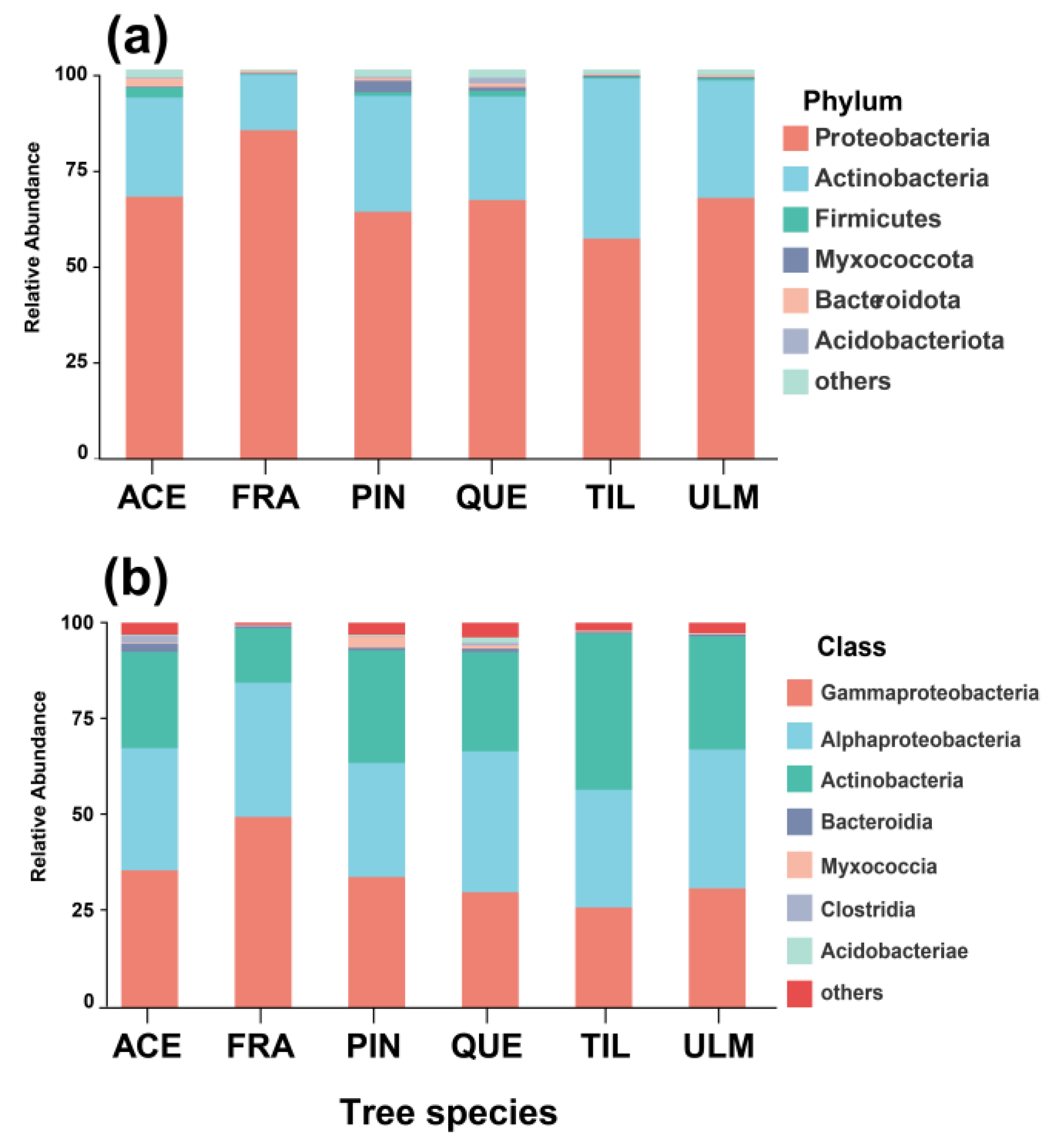
2.1. Phyllosphere Bacterial Diversity and Composition
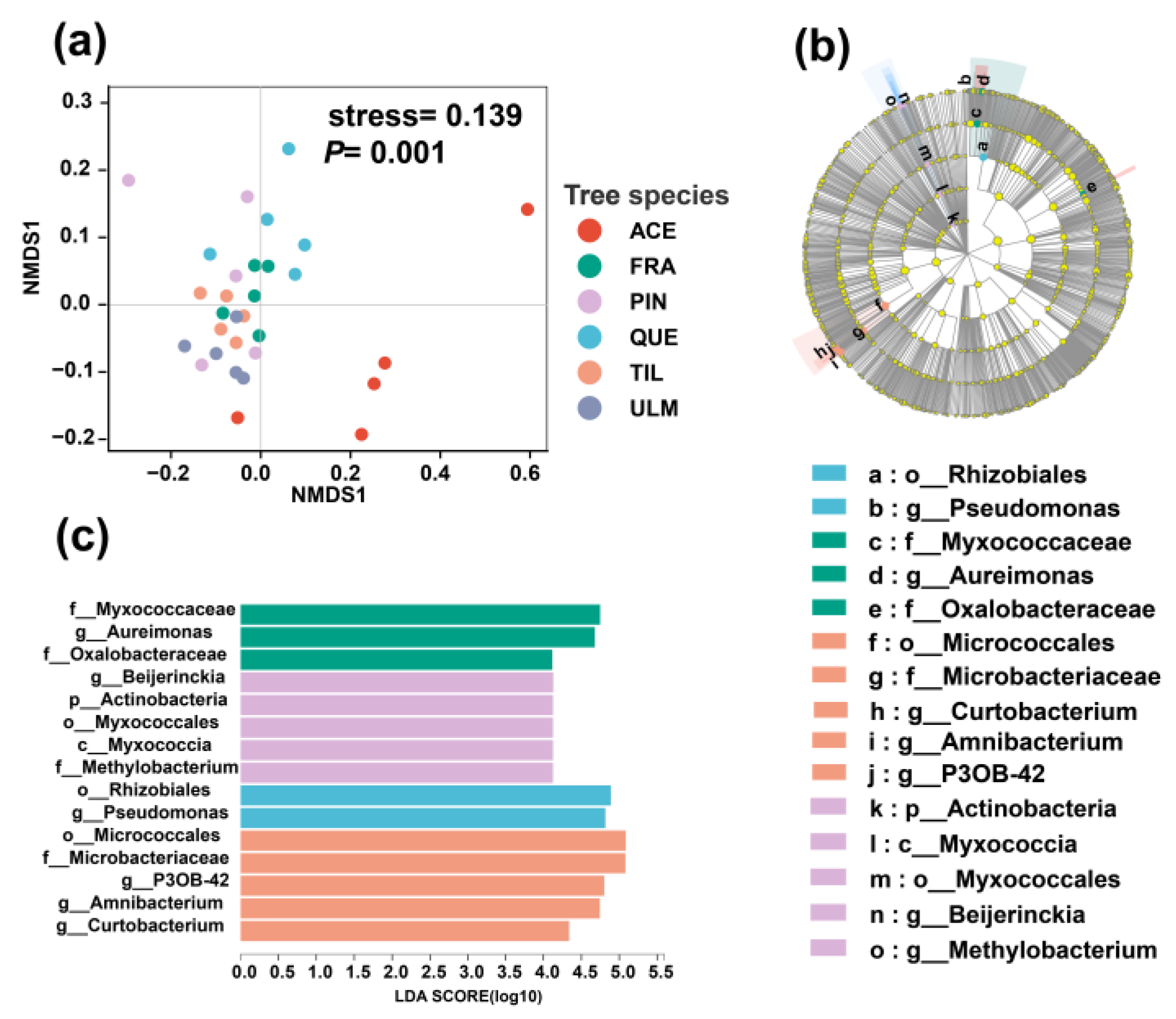
2.2. Differences in Phyllosphere Bacterial Community among Tree Species
2.3. Correlations between Phyllosphere Bacterial Community and Leaf Functional Traits
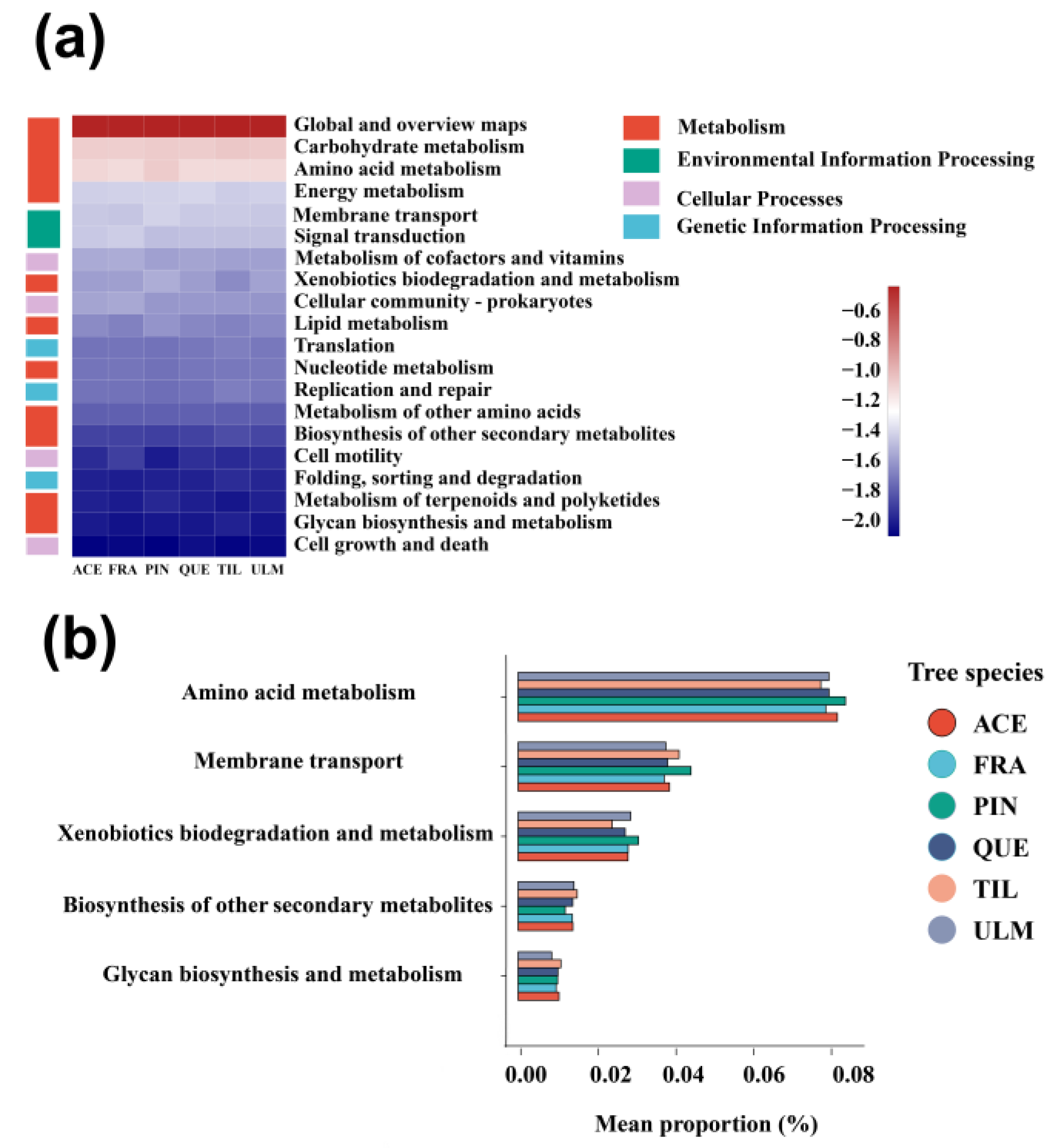
2.4. Functional Genes of Phyllosphere Bacteria
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Leaf Sampling
4.3. Measurements and Calculations of Leaf Functional Traits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Functional Traits | Functions |
|---|---|---|
| LA | Leaf Area | Resource allocation capacity [69] |
| SLA | Specific Leaf Area | Resource allocation capacity [69] |
| LDMC | Leaf Dry Matter Content | Resource allocation capacity [70] |
| LCC13 | Leaf Stable Carbon 13 Content | Water utilization efficiency [70] |
| LNC15 | Leaf Stable Nitrogen 15 Content | Resource utilization efficiency [71] |
| LCC | Leaf Carbon Content | Plant photosynthesis [72] |
| LNC | Leaf Nitrogen Content | Plant photosynthesis [72] |
| LPC | Leaf Phosphorus Content | Plant photosynthesis [72] |
| LKC | Leaf Potassium Content | Plant photosynthesis [72] |
| LCaC | Leaf Calcium Content | Plant metabolism [73] |
| LAlC | Leaf Aluminum Content | Plant metabolism [73] |
| LCuC | Leaf Copper Content | Plant metabolism; resistance to diseases, pests [74] |
| LZnC | Leaf Zinc Content | Plant metabolism; resistance to diseases, pests [74] |
| LSA | Leaf Stomatal Area | Transpiration, photosynthesis [75] |
4.4. Leaf DNA Extraction and Sequencing
4.5. Data Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Last, F. Seasonal incidence of Sporobolomyces on cereal leaves. Trans. Br. Mycol. Soc. 1955, 38, 221–239. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Xiong, C.; Wei, Z.; Chen, Q.L.; Ma, B.; Zhou, S.Y.D.; Tan, J.; Zhang, L.M.; Cui, H.L.; Duan, G.L. Impacts of global change on phyllosphere microbiome. New Phytol. 2021, 234, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Farre-Armengol, G.; Filella, I.; Llusia, J.; Penuelas, J. Bidirectional Interaction between Phyllospheric Microbiotas and Plant Volatile Emissions. Trends Plant Sci. 2016, 21, 854–860. [Google Scholar] [CrossRef]
- Stone Bram, W.G.; Jackson Colin, R. Biogeographic patterns between bacterial phyllosphere communities of the southern magnolia (Magnolia grandiflora) in a small forest. Microb. Ecol. 2016, 71, 954–961. [Google Scholar] [CrossRef]
- Chen, T.; Nomura, K.; Wang, X.; Sohrabi, R.; Xu, J.; Yao, L.; Paasch, B.C.; Ma, L.; Kremer, J.; Cheng, Y.; et al. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature 2020, 580, 653–657. [Google Scholar] [CrossRef]
- Laforest-Lapointe, I.; Paquette, A.; Messier, C.; Kembel, S.W. Leaf bacterial diversity mediates plant diversity and ecosystem function relationships. Nature 2017, 546, 145–147. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E.; Singh, B. Linking the phyllosphere microbiome to plant health. Trends Plant Sci. 2020, 25, 841–844. [Google Scholar] [CrossRef]
- Lee, D.W.; Hong, J.-S.; Kim, S.-H.; Kim, J.W.; Kim, B.S. First Report of Pseudomonas lurida Causing Bacterial Leaf Spot on Miscanthus sinensis. J. Phytopathol. 2014, 162, 195–200. [Google Scholar] [CrossRef]
- Arun, D.K.; Sabarinathan, K.G.; Gomathy, M.; Kannan, R.; Balachandar, D. Mitigation of drought stress in rice crop with plant growth-promoting abiotic stress-tolerant rice phyllosphere bacteria. J. Basic Microbiol. 2020, 60, 768–786. [Google Scholar] [CrossRef]
- Yao, H.; Sun, X.; He, C.; Maitra, P.; Li, X.-C.; Guo, L.-D. Phyllosphere epiphytic and endophytic fungal community and network structures differ in a tropical mangrove ecosystem. Microbiome 2019, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Sun, X.; He, C.; Li, X.-C.; Guo, L.-D. Host identity is more important in structuring bacterial epiphytes than endophytes in a tropical mangrove forest. FEMS Microbiol. Ecol. 2020, 96, fiaa038. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [PubMed]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. Microbiome 2016, 4, e2367. [Google Scholar] [CrossRef] [PubMed]
- Leveau, J.H.J. A brief from the leaf: Latest research to inform our understanding of the phyllosphere microbiome. Curr. Opin. Microbiol. 2019, 49, 41–49. [Google Scholar] [CrossRef]
- Wang, M.; Sun, B. Unlocking the connection: Aging as a lens to examine the effects of climate warming. Innov. Life 2023, 1, 100003. [Google Scholar] [CrossRef]
- Lajoie, G.; Maglione, R.; Kembel, S.W. Adaptive matching between phyllosphere bacteria and their tree hosts in a neotropical forest. Microbiome 2020, 8, 70. [Google Scholar] [CrossRef]
- Hunter, P.J.; Hand, P.; Pink, D.; Whipps, J.M.; Bending, G.D. Both leaf properties and microbe-microbe interactions influence within-species variation in bacterial population diversity and structure in the lettuce (Lactuca species) phyllosphere. Appl. Environ. Microbiol. 2010, 76, 8117–8125. [Google Scholar] [CrossRef]
- Liu, L.; Lu, L.; Li, H.; Meng, Z.; Dong, T.; Peng, C.; Xu, X. Divergence of phyllosphere microbial communities between females and males of the Dioecious Populus cathayana. Mol. Plant-Microbe Interact. 2021, 34, 351–361. [Google Scholar] [CrossRef]
- Remus-Emsermann, M.N.; Schlechter, R.O. Phyllosphere microbiology: At the interface between microbial individuals and the plant host. New Phytol. 2018, 218, 1327–1333. [Google Scholar] [CrossRef]
- Kembel Steven, W.; Mueller Rebecca, C. Plant traits and taxonomy drive host associations in tropical phyllosphere fungal communities. Botany 2014, 92, 303–311. [Google Scholar] [CrossRef]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.M.; Freiberg, M.; Hewitt, C.N.; Itioka, T.; Koh, L.P.; Ma, K.; et al. Forests and their canopies: Achievements and horizons in canopy science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Wilkes, P.; Jones, S.D.; Suarez, L.; Mellor, A.; Woodgate, W.; Soto-Berelov, M.; Haywood, A.; Skidmore, A.K. Mapping forest canopy height across large areas by upscaling ALS estimates with freely available satellite data. Remote Sens. 2015, 7, 12563–12587. [Google Scholar] [CrossRef]
- Ozanne, C.M.; Anhuf, D.; Boulter, S.L.; Keller, M.; Kitching, R.L.; Körner, C.; Meinzer, F.C.; Mitchell, A.; Nakashizuka, T.; Dias, P.S. Biodiversity meets the atmosphere: A global view of forest canopies. Science 2003, 301, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.F.; Scheffers, B.R. Vertical stratification influences global patterns of biodiversity. Ecography 2019, 42, 249. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph Wright, S.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L. CTFS-Forest GEO: A worldwide network monitoring forests in an era of global change. Glob. Chang. Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef]
- Bai, F.; Sang, W.; Axmacher, J.C. Forest vegetation responses to climate and environmental change: A case study from Changbai Mountain, NE China. For. Ecol. Manag. 2011, 262, 2052–2060. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Z.; Liu, X.; Zhang, X.-H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef]
- Sun, X.; Deharveng, L.; Bedos, A.; Chang, L.; Scheu, S.; Wu, D. Changes in diversity and body size of Onychiurinae (Collembola: Onychiuridae) along an altitudinal gradient in Changbai Mountain, China. Soil. Ecol. Lett. 2020, 2, 230–239. [Google Scholar] [CrossRef]
- Liu, N.; Hu, H.; Ma, W.; Deng, Y.; Dimitrov, D.; Wang, Q.; Shrestha, N.; Su, X.; Feng, K.; Liu, Y.; et al. Relationships between soil microbial diversities across an aridity gradient in temperate grasslands. Microb. Ecol. 2022, 85, 1013–1027. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef]
- Redford Amanda, J.; Bowers Robert, M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar] [CrossRef]
- Espenshade, J.; Thijs, S.; Gawronski, S.; Boye, H.; Weyens, N.; Vangronsveld, J. Influence of urbanization on epiphytic bacterial communities of the platanus × hispanica tree leaves in a biennial study. Front. Microbiol. 2019, 10, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, I.; Canedoli, C.; Imperato, V.; Tagliaferri, I.; Gkorezis, P.; Vangronsveld, J.; Padoa Schioppa, E.; Papacchini, M.; Bestetti, G.; Franzetti, A. Diversity and hydrocarbon-degrading potential of epiphytic microbial communities on Platanus × acerifolia leaves in an urban area. Environ. Pollut. 2017, 220, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Smets, W.; Wuyts, K.; Oerlemans, E.; Wuyts, S.; Denys, S.; Samson, R.; Lebeer, S. Impact of urban land use on the bacterial phyllosphere of ivy (Hedera sp.). Atmos. Environ. 2016, 147, 376–383. [Google Scholar] [CrossRef]
- Lymperopoulou, D.S.; Adams, R.I.; Lindow, S.E. Contribution of vegetation to the microbial composition of nearby outdoor air. Appl. Environ. Microbiol. 2016, 82, 3822–3833. [Google Scholar] [CrossRef]
- Leff, J.W.; Del Tredici, P.; Friedman, W.E.; Fierer, N. Spatial structuring of bacterial communities within individual Ginkgo biloba trees. Environ. Microbiol. 2015, 17, 2352–2361. [Google Scholar] [CrossRef]
- Watanabe, K.; Kohzu, A.; Suda, W.; Yamamura, S.; Takamatsu, T.; Takenaka, A.; Koshikawa, M.K.; Hayashi, S.; Watanabe, M. Microbial nitrification in throughfall of a Japanese cedar associated with archaea from the tree canopy. Springerplus 2016, 5, 1596. [Google Scholar] [CrossRef]
- Atamna-Ismaeel, N.; Finkel, O.; Glaser, F.; von Mering, C.; Vorholt, J.A.; Koblizek, M.; Belkin, S.; Beja, O. Bacterial anoxygenic photosynthesis on plant leaf surfaces. Environ. Microbiol. Rep. 2012, 4, 209–216. [Google Scholar] [CrossRef]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Impact of plant genotype and plant habitat in shaping bacterial pathobiome: A comparative study in olive tree. Sci. Rep. 2020, 10, 3475. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Epiphytic and endophytic bacteria on olive tree phyllosphere: Exploring tissue and cultivar effect. Microb. Ecol. 2020, 80, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, C.; Gueidan, C.; Selbmann, L.; Gorbushina, A.A.; Crous, P.W.; Groenewald, J.Z.; Muggia, L.; Grube, M.; Isola, D.; Schoch, C.L.; et al. Phylogeny of rock-inhabiting fungi related to Dothideomycetes. Stud. Mycol. 2009, 61, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-L.; Cai, L.; Wang, H.-C.; Cai, L.-T.; Goodwin, P.; Ma, J.; Wang, F.; Li, Z. Fungal composition and diversity of the tobacco leaf phyllosphere during curing of leaves. Front. Microbiol. 2019, 11, 554051. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Nakanishi, S.; Choe, D. Phytosociological study on natural summer-green forests in Korea. Ecol. Res. 1994, 9, 21–32. [Google Scholar] [CrossRef]
- Mills, G. An urban canopy-layer climate model. Theor. Appl. Climatol. 1997, 57, 229–244. [Google Scholar] [CrossRef]
- Rico, L.; Ogaya, R.; Terradas, J.; Penuelas, J. Community structures of N-2-fixing bacteria associated with the phyllosphere of a Holm oak forest and their response to drought. Plant Biol. 2014, 16, 586–593. [Google Scholar] [CrossRef]
- Abanda-Nkpwatt, D.; Muesch, M.; Tschiersch, J.; Boettner, M.; Schwab, W. Molecular interaction between Methylobacterium extorquens and seedlings: Growth promotion, methanol consumption, and localization of the methanol emission site. J. Exp. Bot. 2006, 57, 4025–4032. [Google Scholar] [CrossRef]
- Kang, S.-M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.-J.; Park, J.-M.; Kim, B.-R.; Shin, D.-H.; Lee, I.-J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- He, F.; Yang, B.; Wang, H.; Yan, Q.; Cao, Y.; He, X. Changes in composition and diversity of fungal communities along Quercus mongolica forests developments in Northeast China. Appl. Soil. Ecol. 2016, 100, 162–171. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Tariq, H.; Latif, S.; Yasmin, H.; Mehmood, A.; Shahid, M.A. Water conservation and plant survival strategies of rhizobacteria under drought stress. Agronomy 2020, 10, 1683. [Google Scholar] [CrossRef]
- Bull, C.T.; Shetty, K.G.; Subbarao, K.V. Interactions between myxobacteria, plant pathogenic fungi, and biocontrol agents. Plant Dis. 2002, 86, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Meyer Katrin, M.; Leveau Johan, H.J. Microbiology of the phyllosphere: A playground for testing ecological concepts. Oecologia 2012, 168, 621–629. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef]
- Kaczmarczyk, A.; Hochstrasser, R.; Vorholt, J.A.; Francez-Charlot, A. Complex two-component signaling regulates the general stress response in Alphaproteobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, E5196–E5204. [Google Scholar] [CrossRef]
- Ryffel, F.; Helfrich, E.J.N.; Kiefer, P.; Peyriga, L.; Portais, J.-C.; Piel, J.; Vorholt, J.A. Metabolic footprint of epiphytic bacteria on Arabidopsis thaliana leaves. ISME J. 2016, 10, 632–643. [Google Scholar] [CrossRef]
- Mueller, D.B.; Schubert, O.T.; Roest, H.; Aebersold, R.; Vorholt, J.A. Systems-level Proteomics of Two Ubiquitous Leaf Commensals Reveals Complementary Adaptive Traits for Phyllosphere Colonization. Mol. Cell. Proteom. 2016, 15, 3256–3269. [Google Scholar] [CrossRef]
- Yadav, R.K.P.; Papatheodorou, E.M.; Karamanoli, K.; Constantinidou, H.-I.A.; Vokou, D. Abundance and diversity of the phyllosphere bacterial communities of Mediterranean perennial plants that differ in leaf chemistry. Chemoecology 2008, 18, 217–226. [Google Scholar] [CrossRef]
- Nehls, U.; Mikolajewski, S.; Magel, E.; Hampp, R. Carbohydrate metabolism in ectomycorrhizas: Gene expression, monosaccharide transport and metabolic control. New Phytol. 2001, 150, 533–541. [Google Scholar] [CrossRef]
- Neis, E.P.; Dejong, C.H.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Delmotte, N.; Knief, C.; Chaffron, S.; Innerebner, G.; Roschitzki, B.; Schlapbach, R.; von Mering, C.; Vorholt Julia, A. Community proteogenomics reveals insights into the physiology of phyllosphere bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 16428–16433. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil. Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Yuan, Z.; Ali, A.; Ruiz-Benito, P.; Jucker, T.; Mori, A.; Wang, S.; Zhang, X.; Li, H.; Hao, Z.; Wang, X.; et al. Above- and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. J. Ecol. 2020, 108, 2012–2024. [Google Scholar] [CrossRef]
- Shao, G.; Schall, P.; Weishampel, J.F. Dynamic simulations of mixed broadleaved-Pinus koraiensis forests in the Changbaishan Biosphere Reserve of China. For. Ecol. Manag. 1994, 70, 169–181. [Google Scholar] [CrossRef]
- Perez-Harguindeguy, N.; Diaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Zhao, W.L.; Chen, Y.J.; Brodribb, T.J.; Cao, K.F. Weak co-ordination between vein and stomatal densities in 105 angiosperm tree species along altitudinal gradients in Southwest China. Funct. Plant Biol. 2016, 43, 1126–1133. [Google Scholar] [CrossRef]
- Wright, I.; Reich, P.; Westoby, M.; Ackerly, D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Cornelissen, J.; Diemer, M.; Flexas, J.; et al. The world-wide leaf economics spectrum. Nature 2004, 21, 821–827. [Google Scholar] [CrossRef]
- Ripullone, F.; Lauteri, M.; Grassi, G.; Amato, M.; Borghetti, M. Variation in nitrogen supply changes water-use efficiency of Pseudotsuga menziesii and Populus x euroamericana; a comparison of three approaches to determine water-use efficiency. Tree Physiol. 2004, 24, 671–679. [Google Scholar] [CrossRef]
- Saitoh, T.; Seiwa, K.; Nishiwaki, A. Effects of resource heterogeneity on nitrogen translocation within clonal fragments of Sasa palmata: An isotopic (15N) assessment. Ann. Bot. 2006, 98, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Longstreth, D.J.; Nobel, P.S. Nutrient Influences on Leaf Photosynthesis: Effects of nitrogen, phosphorus, and potassium for Gossypium hirsutum L. Plant Physiol. 1980, 65, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Bollard, E.; Butler, G. Mineral nutrition of plants. Annu. Rev. Plant Physiol. 1966, 17, 77–112. [Google Scholar] [CrossRef]
- Pollard, A.J.; Powell, K.D.; Harper, F.A.; Smith, J.A.C. The genetic basis of metal hyperaccumulation in plants. Crit. Rev. Plant Sci. 2002, 21, 539–566. [Google Scholar] [CrossRef]
- Sharkey, T.D. Transpiration-induced changes in the photosynthetic capacity of leaves. Planta 1984, 160, 143–150. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Dixon, P. Vegan, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]





| Comparison among Tree Species | R | p |
|---|---|---|
| Global | 0.300 | <0.001 |
| ULM vs. ACE | 0.216 | 0.025 |
| ULM vs. TIL | 0.228 | 0.023 |
| ULM vs. PIN | 0.504 | 0.010 |
| ULM vs. FRA | 0.636 | 0.008 |
| ULM vs. QUE | 0.796 | 0.008 |
| ACE vs. TIL | 0.260 | 0.016 |
| ACE vs. PIN | 0.480 | 0.020 |
| ACE vs. FRA | 0.356 | 0.016 |
| ACE vs. QUE | 0.312 | 0.025 |
| TIL vs. PIN | 0.524 | 0.009 |
| TIL vs. FRA | 0.792 | 0.011 |
| TIL vs. QUE | 0.696 | 0.010 |
| PIN vs. FRA | 0.540 | 0.011 |
| PIN vs. QUE | 0.488 | 0.007 |
| FRA vs. QUE | 0.660 | 0.011 |
| ACE (Mean ± SE) | FRA (Mean ± SE) | PIN (Mean ± SE) | QUE (Mean ± SE) | TIL (Mean ± SE) | ULM (Mean ± SE) | |
|---|---|---|---|---|---|---|
| LA (cm2) *** | 24.52 ± 1.69 | 36.08 ± 2.45 | 0.73 ± 0.035 | 65.36 ± 10.88 | 38.26 ± 3.86 | 17.02 ± 0.53 |
| SLA (cm2/g) ** | 190.05 ± 30.52 | 260.03 ± 36.44 | 84.84 ± 11.57 | 310.555 ± 14.59 | 315.12 ± 20.76 | 76.56 ± 12.01 |
| LDMC (g/g) ** | 0.092 ± 0.014 | 0.15 ± 0.04 | 0.02 ± 0.00 | 0.30 ± 0.04 | 0.17 ± 0.02 | 0.10 ± 0.01 |
| LCC13 (d13C/12C) ** | −30.58 ± 0.30 | −27.78 ± 0.53 | −30.51 ± 0.478 | −29.47 ± 0.24 | −29.68 ± 0.52 | −29.32 ± 0.30 |
| LNC15 (%) * | −2.39 ± 0.77 | 0.18 ± 0.67 | 0.48 ± 0.40 | −0.20 ± 0.19 | 1.01 ± 0.49 | −1.55 ± 0.44 |
| LCC (%) | 41.73 ± 0.09 | 42.34 ± 0.38 | 46.53 ± 0.70 | 43.04 ± 0.15 | 43.87 ± 0.38 | 39.30 ± 0.28 |
| LNC (%) | 1.96 ± 0.08 | 1.90 ± 0.26 | 1.604 ± 0.12 | 2.32 ± 0.08 | 2.09 ± 0.16 | 1.92 ± 0.11 |
| LPC (g/kg) | 1.87 ± 0.15 | 1.94 ± 0.49 | 2.10 ± 0.10 | 2.68 ± 0.26 | 3.03 ± 0.30 | 2.11 ± 0.11 |
| LKC (g/kg) | 0.07 ± 0.01 | 0.08 ± 0.00 | 0.04 ± 0.01 | 0.01 ± 0.00 | 0.08 ± 0.01 | 0.07 ± 0.01 |
| LCaC (g/100 g) ** | 0.24 ± 0.01 | 0.25 ± 0.03 | 0.07 ± 0.02 | 0.139 ± 0.01 | 0.17 ± 0.02 | 0.30 ± 0.02 |
| LAlC (g/kg) *** | 0.63 ± 0.07 | 0.46 ± 0.07 | 1.63 ± 0.31 | 0.65 ± 0.14 | 0.45 ± 0.08 | 0.45 ± 0.06 |
| LCuC (g/kg) ** | 4.90 ± 0.62 | 4.12 ± 0.52 | 4.77 ± 0.14 | 4.90 ± 0.62 | 3.44 ± 0.158 | 5.11 ± 0.50 |
| LZnC (mg/kg) *** | 63.99 ± 7.39 | 25.56 ± 1.15 | 64.49 ± 5.77 | 41.42 ± 8.66 | 30.34 ± 9.19 | 33.21 ± 5.33 |
| LSA (10−9 m2) *** | 7.37 ± 0.02 | 17.06 ± 0.05 | 12.96 ± 0.10 | 24.24 ± 0.07 | 7.90 ± 0.07 | 18.74 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Ye, J.; Lin, F.; Wang, X.; Yang, T.; Bi, B.; Mao, Z.; Fang, S.; Wang, X.; Hao, Z.; et al. Relationships between Phyllosphere Bacterial Communities and Leaf Functional Traits in a Temperate Forest. Plants 2023, 12, 3854. https://doi.org/10.3390/plants12223854
Yuan Z, Ye J, Lin F, Wang X, Yang T, Bi B, Mao Z, Fang S, Wang X, Hao Z, et al. Relationships between Phyllosphere Bacterial Communities and Leaf Functional Traits in a Temperate Forest. Plants. 2023; 12(22):3854. https://doi.org/10.3390/plants12223854
Chicago/Turabian StyleYuan, Zuoqiang, Ji Ye, Fei Lin, Xing Wang, Teng Yang, Boyuan Bi, Zikun Mao, Shuai Fang, Xugao Wang, Zhanqing Hao, and et al. 2023. "Relationships between Phyllosphere Bacterial Communities and Leaf Functional Traits in a Temperate Forest" Plants 12, no. 22: 3854. https://doi.org/10.3390/plants12223854