
Impact of Aerated Drip Irrigation and Nitrogen Application on Soil Properties, Soil Bacterial Communities and Agronomic Traits of Cucumber in a Greenhouse System
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Cucumber Agronomic Traits
2.3. Soil Sampling
2.4. Soil Physicochemical Properties
2.5. Soil DNA Extraction, PCR Amplification, and 16S rRNA Gene Sequencing
2.6. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. Cucumber Root Growth, Biomass, Yield, and IWUE
3.3. Soil Bacterial Community Diversity and Composition
3.4. Composition and Functional Forecast of Soil Bacterial Community
3.5. Co-Occurrence Network Analysis of Bacterial Community
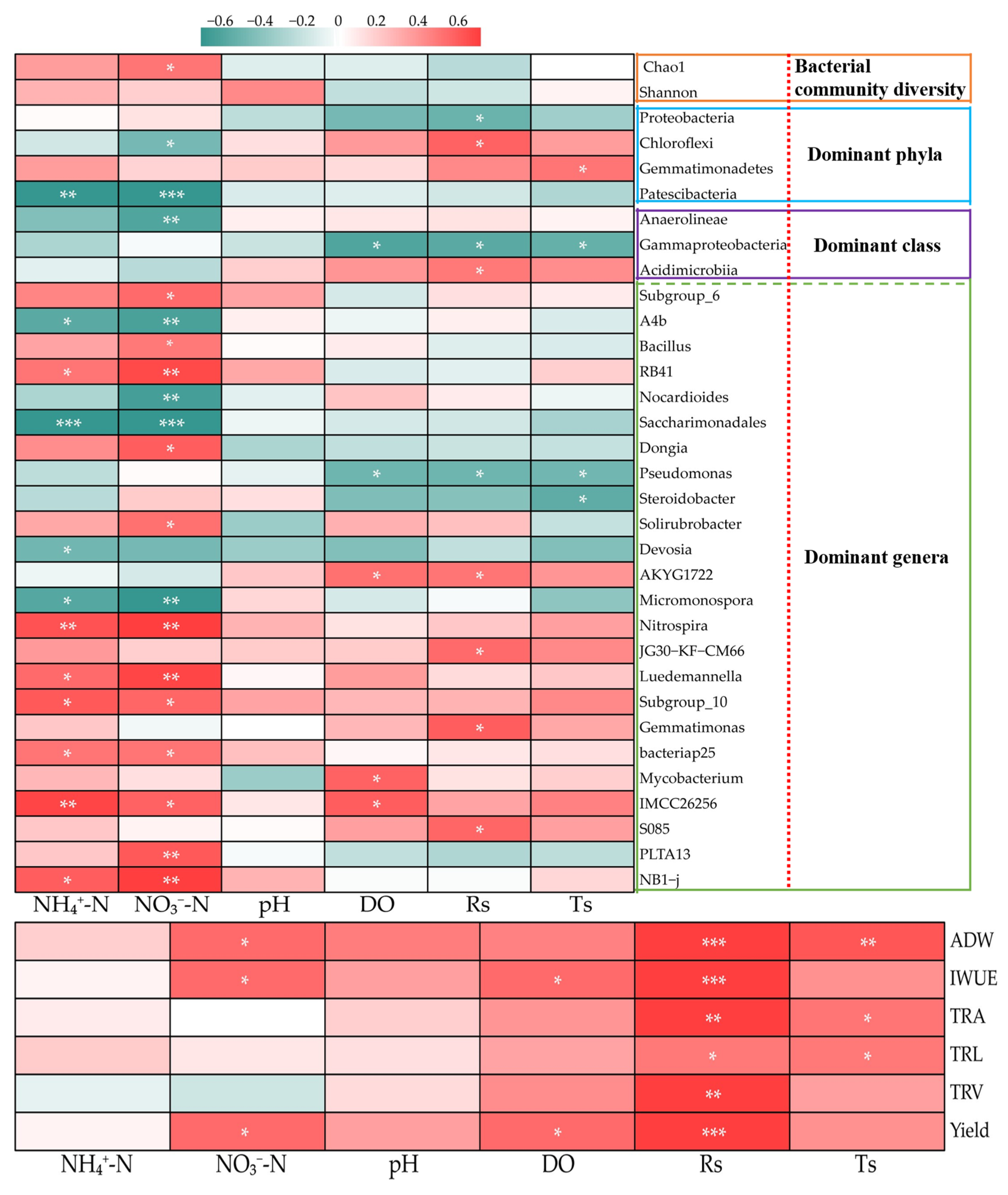
3.6. Relationships between the Bacterial Community, Soil Physicochemical Properties, and Cucumber Agronomic Traits
4. Discussion
4.1. Effects of ADI on Bacterial Community Diversity and Structure
4.2. Effect of ADI on the Soil Microbial Functionality and Co-Occurrence Networks of Bacterial Community
4.3. ADI Improved Soil Properties and Boosted Cucumber Performance by Changing the Bacterial Community
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Liu, Z.H.; Xi, B.D.; He, X.S.; Li, Q.L.; Qi, Y.J.; Jin, M.Y.; Guo, Y. Characteristics of groundwater pollution in a vegetable cultivation area of typical facility agriculture in a developed city. Ecol. Indic. 2019, 105, 709–716. [Google Scholar] [CrossRef]
- Torres Pineda, I.; Lee, Y.D.; Kim, Y.S.; Lee, S.M.; Park, K.S. Review of inventory data in life cycle assessment applied in production of fresh tomato in greenhouse. J. Clean. Prod. 2021, 282, 124395. [Google Scholar] [CrossRef]
- Padalia, K.; Bargali, S.S.; Bargali, K.; Manral, V. Soil microbial biomass phosphorus under different land use systems of Central Himalaya. Trop. Ecol. 2022, 63, 30–48. [Google Scholar] [CrossRef]
- Pant, M.; Negi, G.C.S.; Kumar, P. Macrofauna contributes to organic matter decomposition and soil quality in Himalayan agroecosystems, India. Appl. Soil Ecol. 2017, 120, 20–29. [Google Scholar] [CrossRef]
- Bargali, S.S.; Padalia, K.; Bargali, K. Effects of tree fostering on soil health and microbial biomass under different land use systems in the Central Himalayas. Land. Degrad. Dev. 2019, 30, 1984–1998. [Google Scholar] [CrossRef]
- Karki, H.; Bargali, K.; Bargali, S.S. Spatial and temporal trends in soil N-mineralization rates under the agroforestry systems in Bhabhar belt of Kumaun Himalaya, India. Agroforest. Syst. 2021, 95, 1603–1617. [Google Scholar] [CrossRef]
- Manral, V.; Bargali, K.; Bargali, S.S.; Shahi, C. Changes in soil biochemical properties following replacement of Banj oak forest with Chir pine in Central Himalaya, India. Ecol. Process. 2020, 9, 30. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, P.J.; Wang, L.; Sun, G.Q.; Zhao, J.Y.; Zhang, H.; Du, N. The influence of facility agriculture production on phthalate esters distribution in black soils of northeast China. Sci. Total Environ. 2015, 506–507, 118–125. [Google Scholar] [CrossRef]
- Hu, W.Y.; Zhang, Y.X.; Huang, B.; Teng, Y. Soil environmental quality in greenhouse vegetable production systems in eastern China: Current status and management strategies. Chemosphere 2017, 170, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.Q.; Jia, Z.X.; Zhang, X.; Shao, H.B. Effects of Soil Rhizosphere Aeration on the Root Growth and Water Absorption of Tomato. CLEAN-Soil Air Water 2012, 40, 1364–1371. [Google Scholar] [CrossRef]
- Zhou, Y.P.; Zhou, B.; Xu, F.P.; Muhammad, T.; Li, Y.K. Appropriate dissolved oxygen concentration and application stage of micro-nano bubble water oxygation in greenhouse crop plantation. Agr. Water Manag. 2019, 223, 105713. [Google Scholar] [CrossRef]
- Pedersen, O.; Sauter, M.; Colmer, T.D.; Nakazono, M. Regulation of root adaptive anatomical and morphological traits during low soil oxygen. New. Phytol. 2021, 229, 42–49. [Google Scholar] [CrossRef]
- Niu, W.Q.; Guo, Q.; Zhou, X.B.; Helmers, M.J. Effect of Aeration and Soil Water Redistribution on the Air Permeability under Subsurface Drip Irrigation. Soil. Sci. Soc. Am. J. 2012, 76, 815–820. [Google Scholar] [CrossRef]
- Zhang, Q.; Du, Y.D.; Cui, B.J.; Sun, J.; Wang, J.; Wu, M.L.; Niu, W.Q. Aerated irrigation offsets the negative effects of nitrogen reduction on crop growth and water-nitrogen utilization. J. Clean. Prod. 2021, 313, 127917. [Google Scholar] [CrossRef]
- Du, Y.D.; Zhang, Q.; Cui, B.J.; Sun, J.; Wang, Z.; Ma, L.H.; Niu, W.Q. Aerated irrigation improves tomato yield and nitrogen use efficiency while reducing nitrogen application rate. Agr. Water Manag. 2020, 235, 106152. [Google Scholar] [CrossRef]
- Niu, W.Q.; Zhang, E.X.; Lv, C.; Sun, J.; Dong, A.H.; Wu, M.L. Clogging characteristics of muddy drip irrigation dripper under different aeration and particle size conditions. Trans. Chin. Soc. Agric. Mach. 2021, 52, 144–152. (In Chinese) [Google Scholar]
- Zhang, Q.; Niu, W.Q.; Du, Y.D.; Sun, J.; Cui, B.J.; Zhang, E.X.; Wang, Y.B.; Siddique, K.H.M. Effect of aerated drip irrigation and nitrogen doses on N2O emissions, microbial activity, and yield of tomato and muskmelon under greenhouse conditions. Agr. Water Manag. 2023, 283, 108321. [Google Scholar] [CrossRef]
- Zhou, Y.P.; Bastida, F.; Zhou, B.; Sun, Y.F.; Gu, T.; Li, S.Q.; Li, Y.K. Soil fertility and crop production are fostered by micro-nano bubble irrigation with associated changes in soil bacterial community. Soil Biol. Biochem. 2020, 141, 107663. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Zhu, J.J.; Niu, W.Q.; Zhang, Z.H.; Siddique, K.H.M.; Dan, S.; Yang, R.Y. Distinct roles for soil bacterial and fungal communities associated with the availability of carbon and phosphorus under aerated drip irrigation. Agr. Water Manag. 2022, 274, 107925. [Google Scholar] [CrossRef]
- Walz, J.; Knoblauch, C.; Böhme, L.; Pfeiffer, E.M. Regulation of soil organic matter decomposition in permafrost-affected Siberian tundra soils—Impact of oxygen availability, freezing and thawing, temperature, and labile organic matter. Soil Biol. Biochem. 2017, 110, 34–43. [Google Scholar] [CrossRef]
- Zhu, Y.; Shen, R.; He, J.; Wang, Y.; Han, X.; Jia, Z. China Soil Microbiome Initiative: Progress and Perspective. Bull. Chin. Acad. Sci. (Chin. Version) 2017, 32, 554–565. [Google Scholar]
- Manral, V.; Bargali, K.; Bargali, S.S.; Jhariya, M.K.; Padalia, K. Relationships between soil and microbial biomass properties and annual flux of nutrients in Central Himalaya forests, India. Land. Degrad. Dev. 2022, 33, 2014–2025. [Google Scholar] [CrossRef]
- Zeglin, L.H.; Dahm, C.N.; Barrett, J.E.; Gooseff, M.N.; Fitpatrick, S.K.; Takacs-Vesbach, C.D. Bacterial Community Structure Along Moisture Gradients in the Parafluvial Sediments of Two Ephemeral Desert Streams. Microb. Ecol. 2011, 61, 543–556. [Google Scholar] [CrossRef]
- Truu, M.; Ostonen, I.; Preem, J.K.; Lõhmus, K.; Nõlvak, H.; Ligi, T.; Rosenvald, K.; Parts, K.; Kupper, P.; Truu, J. Elevated Air Humidity Changes Soil Bacterial Community Structure in the Silver Birch Stand. Front. Microbiol. 2017, 8, 557. [Google Scholar] [CrossRef]
- Bastida, F.; Torres, I.F.; Abadía, J.; Romero-Trigueros, C.; Ruiz-Navarro, A.; Alarcón, J.J.; García, C.; Nicolás, E. Comparing the impacts of drip irrigation by freshwater and reclaimed wastewater on the soil microbial community of two citrus species. Agr. Water Manag. 2018, 203, 53–62. [Google Scholar] [CrossRef]
- Zolti, A.; Green, S.J.; Ben Mordechay, E.; Hadar, Y.; Minz, D. Root microbiome response to treated wastewater irrigation. Sci. Total Environ. 2019, 655, 899–907. [Google Scholar] [CrossRef]
- Chen, Z.; Maltz, M.R.; Cao, J.X.; Yu, H.; Shang, H.; Aronson, E. Elevated O3 alters soil bacterial and fungal communities and the dynamics of carbon and nitrogen. Sci. Total Environ. 2019, 677, 272–280. [Google Scholar] [CrossRef]
- Ye, G.P.; Lin, Y.X.; Kuzyakov, Y.; Liu, D.Y.; Luo, J.; Lindsey, S.; Wang, W.J.; Fan, J.B.; Ding, W.X. Manure over crop residues increases soil organic matter but decreases microbial necromass relative contribution in upland Ultisols: Results of a 27-year field experiment. Soil Biol. Biochem. 2019, 134, 15–24. [Google Scholar] [CrossRef]
- Georgiou, K.; Abramoff, R.Z.; Harte, J.; Riley, W.J.; Torn, M.S. Microbial community-level regulation explains soil carbon responses to long-term litter manipulations. Nat. Commun. 2017, 8, 1223. [Google Scholar] [CrossRef]
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Banerjee, S.; Thrall, P.H.; Bissett, A.; Van der Heijden, M.G.A.; Richardson, A.E. Linking microbial co-occurrences to soil ecological processes across a woodland-grassland ecotone. Ecol. Evol. 2018, 8, 8217–8230. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.Z.; Zhuang, S.Y.; Gao, J.S.; Tang, L.Z.; Harindintwali, J.D.; Wang, F. Aeration increases soil bacterial diversity and nutrient transformation under mulching-induced hypoxic conditions. Sci. Total Environ. 2022, 817, 153017. [Google Scholar] [CrossRef]
- Wang, J.W.; Niu, W.Q.; Zhang, M.Z.; Li, Y. Effect of alternate partial root-zone drip irrigation on soil bacterial communities and tomato yield. Appl. Soil Ecol. 2017, 119, 250–259. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef]
- Wu, B.H.; Luo, H.Y.; Wang, X.T.; Liu, H.; Peng, H.; Sheng, M.P.; Xu, F.; Xu, H. Effects of environmental factors on soil bacterial community structure and diversity in different contaminated districts of Southwest China mine tailings. Sci. Total Environ. 2022, 802, 149899. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Niu, W.Q.; Zhang, M.Z.; Wang, J.W.; Zhang, Z.X. Artificial soil aeration increases soil bacterial diversity and tomato root performance under greenhouse conditions. Land. Degrad. Dev. 2020, 31, 1443–1461. [Google Scholar] [CrossRef]
- Zhou, Y.P.; Bastida, F.; Liu, Y.Z.; He, J.; Chen, W.; Wang, X.Y.; Xiao, Y.; Song, P.; Li, Y.K. Impacts and mechanisms of nanobubbles level in drip irrigation system on soil fertility, water use efficiency and crop production: The perspective of soil microbial community. J. Clean. Prod. 2022, 333, 130050. [Google Scholar] [CrossRef]
- Liu, X.Y.; Hu, S.H.; Sun, R.; Wu, Y.G.; Qiao, Z.X.; Wang, S.C.; Zhang, Z.H.; Cui, C.W. Dissolved oxygen disturbs nitrate transformation by modifying microbial community, co-occurrence networks, and functional genes during aerobic-anoxic transition. Sci. Total Environ. 2021, 790, 148245. [Google Scholar] [CrossRef]
- Baram, S.; Evans, J.F.; Berezkin, A.; Ben-Hur, M. Irrigation with treated wastewater containing nanobubbles to aerate soils and reduce nitrous oxide emissions. J. Clean. Prod. 2021, 280, 124509. [Google Scholar] [CrossRef]
- Liu, D.T.; Song, C.C.; Xin, Z.H.; Fang, C.; Liu, Z.H.; Xu, Y.P. Agricultural management strategies for balancing yield increase, carbon sequestration, and emission reduction after straw return for three major grain crops in China: A meta-analysis. J. Environ. Manag. 2023, 340, 117965. [Google Scholar] [CrossRef]
- Zhu, J.J.; Xu, N.; Siddique, K.H.M.; Zhang, Z.H.; Niu, W. Aerated drip irrigation improves water and nitrogen uptake efficiencies of tomato roots with associated changes in the antioxidant system. Sci. Hortic. 2022, 306, 111471. [Google Scholar] [CrossRef]
- Liu, J.A.; Shu, A.P.; Song, W.F.; Shi, W.C.; Li, M.; Zhang, W.X.; Li, Z.Z.; Liu, G.R.; Yuan, F.S.; Zhang, S.X.; et al. Long-term organic fertilizer substitution increases rice yield by improving soil properties and regulating soil bacteria. Geoderma 2021, 404, 115287. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Wang, J.M.; Feng, Y. The effects of biochar addition on soil physicochemical properties: A review. Catena 2021, 202, 105284. [Google Scholar] [CrossRef]
- Lei, H.J.; Jin, C.C.; Xiao, Z.Y.; Chen, J.; Leghari, S.J.; Pan, H.W. Relationship between pepper (Capsicum annuum L.) root morphology, inter-root soil bacterial community structure and diversity under water–air intercropping conditions. Planta 2023, 257, 98. [Google Scholar] [CrossRef] [PubMed]
- Šibanc, N.; Dumbrell, A.J.; Mandić-Mulec, I.; Maček, I. Impacts of naturally elevated soil CO2 concentrations on communities of soil archaea and bacteria. Soil Biol. Biochem. 2014, 68, 348–356. [Google Scholar] [CrossRef]
- Krause, S.M.B.; Dohrmann, A.B.; Gillor, O.; Christensen, B.T.; Merbach, I.; Tebbe, C.C. Soil properties and habitats determine the response of bacterial communities to agricultural wastewater irrigation. Pedosphere 2020, 30, 146–158. [Google Scholar] [CrossRef]
- Tian, J.Q.; Liu, L.F.; Chen, H.; Zhong, L.; Zhou, X.; Jiang, L.; Zhan, W.; Wang, Y.F. Aerobic environments in combination with substrate additions to soil significantly reshape depth-dependent microbial distribution patterns in Zoige peatlands, China. Appl. Soil Ecol. 2022, 170, 104252. [Google Scholar] [CrossRef]
- Wu, D.M.; Dai, Q.P.; Liu, X.Z.; Fan, Y.P.; Wang, J.X. Comparison of bacterial community structure and potential functions in hypoxic and non-hypoxic zones of the Changjiang Estuary. PLoS ONE 2019, 14, e0217431. [Google Scholar] [CrossRef]
- Qiao, Z.X.; Sun, R.; Wu, Y.G.; Hu, S.H.; Liu, X.Y.; Chan, J.W.; Mi, X.H. Characteristics and metabolic pathway of the bacteria for heterotrophic nitrification and aerobic denitrification in aquatic ecosystems. Environ. Res. 2020, 191, 110069. [Google Scholar] [CrossRef]
- Gao, G.F.; Li, P.F.; Zhong, J.X.; Shen, Z.-J.; Chen, J.; Li, Y.T.; Isabwe, A.; Zhu, X.Y.; Ding, Q.S.; Zhang, S.; et al. Spartina alterniflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrification in mangrove wetland. Sci. Total Environ. 2019, 653, 231–240. [Google Scholar] [CrossRef]
- Chowdhury, M.A.; de Neergaard, A.; Jensen, L.S. Potential of aeration flow rate and bio-char addition to reduce greenhouse gas and ammonia emissions during manure composting. Chemosphere 2014, 97, 16–25. [Google Scholar] [CrossRef]
- Fu, X.H.; Song, Q.L.; Li, S.Q.; Shen, Y.F.; Yue, S.C. Dynamic changes in bacterial community structure are associated with distinct priming effect patterns. Soil Biol. Biochem. 2022, 169, 108671. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Q.H.; Wang, J.M.; Ma, Y.X.; Cao, B. Responses of heavy metals mobility and resistant bacteria to adding time of activated carbon during chicken manure composting*. Environ. Pollut. 2021, 290, 118070. [Google Scholar] [CrossRef] [PubMed]
- Che, R.X.; Wang, S.P.; Wang, Y.F.; Xu, Z.H.; Wang, W.; Rui, Y.C.; Wang, F.; Hu, J.M.; Tao, J.; Cui, X.Y. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil Biol. Biochem. 2019, 139, 107611. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, L.; Xue, S.; Liu, B.J.; Jin, G. Artificial root exudates excite bacterial nitrogen fixation in the subsurface of mine soils. Appl. Soil Ecol. 2021, 157, 103774. [Google Scholar] [CrossRef]
- Bello, A.; Wang, B.; Zhao, Y.; Yang, W.; Ogundeji, A.; Deng, L.T.; Egbeagu, U.U.; Yu, S.; Zhao, L.Y.; Li, D.T.; et al. Composted biochar affects structural dynamics, function and co-occurrence network patterns of fungi community. Sci. Total Environ. 2021, 775, 145672. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Chai, X.T.; Tariq, A.; Zeng, F.J.; Li, X.Y.; Graciano, C. Intercropping Systems Modify Desert Plant-Associated Microbial Communities and Weaken Host Effects in a Hyper-Arid Desert. Front. Microbiol. 2021, 12, 754453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Juneau, P.; Huang, R.L.; He, Z.; Sun, B.; Zhou, J.Z.; Liang, Y.T. Coexistence between antibiotic resistance genes and metal resistance genes in manure-fertilized soils. Geoderma 2021, 382, 114760. [Google Scholar] [CrossRef]
- Price, G.W.; Langille, M.G.I.; Yurgel, S.N. Microbial co-occurrence network analysis of soils receiving short- and long-term applications of alkaline treated biosolids. Sci. Total Environ. 2021, 751, 141687. [Google Scholar] [CrossRef]
- Wang, R.Q.; Xiao, Y.P.; Lv, F.J.; Hu, L.Y.; Wei, L.G.; Yuan, Z.Q.; Lin, H.X. Bacterial community structure and functional potential of rhizosphere soils as influenced by nitrogen addition and bacterial wilt disease under continuous sesame cropping. Appl. Soil Ecol. 2018, 125, 117–127. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Bei, S.K.; Li, B.S.; Zhang, J.L.; Christie, P.; Li, X.L. Organic fertilizer, but not heavy liming, enhances banana biomass, increases soil organic carbon and modifies soil microbiota. Appl. Soil Ecol. 2019, 136, 67–79. [Google Scholar] [CrossRef]
- Chen, L.F.; He, Z.B.; Wu, X.R.; Du, J.; Zhu, X.; Lin, P.F.; Tian, Q.Y.; Kong, J.Q. Linkages between soil respiration and microbial communities following afforestation of alpine grasslands in the northeastern Tibetan Plateau. Appl. Soil Ecol. 2021, 161, 103882. [Google Scholar] [CrossRef]
- Wang, Y.; Hong, Y.; Tian, Y.; Tian, G.Q.; Zhang, J.H.; Wu, H.; Bai, Y.; Qian, J. Changes in bacterial community composition and soil properties altered the response of soil respiration to rain addition in desert biological soil crusts. Geoderma 2022, 409, 115635. [Google Scholar] [CrossRef]
- Chen, J.H.; He, Y.; Wang, J.H.; Huang, M.S.; Guo, C.X. Dynamics of nitrogen transformation and bacterial community with different aeration depths in malodorous river. World. J. Microb. Biot. 2019, 35, 196. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | N Fertilizer Application Rate (kg ha−1) | Irrigation Water Dissolved Oxygen (mg L−1) |
|---|---|---|
| O1N1 | 240 | 5 |
| O2N1 | 240 | 10 |
| O3N1 | 240 | 20 |
| O1N2 | 360 | 5 |
| O2N2 | 360 | 10 |
| O3N2 | 360 | 20 |
| Group | R-Value | p-Value |
|---|---|---|
| O1N1 vs. O2N1 | 0.6295 | 0.026 |
| O1N1 vs. O3N1 | 0.4424 | 0.105 |
| O2N1 vs. O3N1 | 0.0740 | 0.776 |
| O1N2 vs. O2N2 | 0.3704 | 0.112 |
| O1N2 vs. O3N2 | 0.5828 | 0.041 |
| O2N2 vs. O3N2 | 0.2222 | 0.280 |
| O1N1 vs. O1N2 | 0.2963 | 0.084 |
| O2N1 vs. O2N2 | 0.4526 | 0.103 |
| O3N1 vs. O3N2 | 0.6034 | 0.032 |
| O1 | O2 | O3 | N1 | N2 | |
|---|---|---|---|---|---|
| Nodes | 121 | 140 | 134 | 135 | 131 |
| Edges | 897 | 1041 | 790 | 928 | 905 |
| Average path length | 3.16 | 2.94 | 3.14 | 2.93 | 2.77 |
| Clustering coefficient | 0.623 | 0.606 | 0.621 | 0.587 | 0.602 |
| Average degree | 12.81 | 15.54 | 13.06 | 13.75 | 13.82 |
| Modularity | 0.512 | 0.581 | 0.575 | 0.476 | 0.436 |
| Image density | 0.117 | 0.092 | 0.109 | 0.103 | 0.106 |
| Positive links/Negative Links (P/N) | 0.82 | 1.10 | 0.91 | 0.94 | 1.21 |
| Environmental Factor | R-Value | p-Value |
|---|---|---|
| pH | 0.1099 | 0.266 |
| DO | 0.3105 | 0.007 ** |
| Rs | 0.3520 | 0.002 ** |
| TS | 0.1748 | 0.089 |
| NO3−-N | 0.2767 | 0.019 * |
| NH4+-N | −0.0225 | 0.503 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Z.; Lei, H.; Lian, Y.; Zhang, Z.; Pan, H.; Yin, C.; Dong, Y. Impact of Aerated Drip Irrigation and Nitrogen Application on Soil Properties, Soil Bacterial Communities and Agronomic Traits of Cucumber in a Greenhouse System. Plants 2023, 12, 3834. https://doi.org/10.3390/plants12223834
Xiao Z, Lei H, Lian Y, Zhang Z, Pan H, Yin C, Dong Y. Impact of Aerated Drip Irrigation and Nitrogen Application on Soil Properties, Soil Bacterial Communities and Agronomic Traits of Cucumber in a Greenhouse System. Plants. 2023; 12(22):3834. https://doi.org/10.3390/plants12223834
Chicago/Turabian StyleXiao, Zheyuan, Hongjun Lei, Yingji Lian, Zhenhua Zhang, Hongwei Pan, Chen Yin, and Yecheng Dong. 2023. "Impact of Aerated Drip Irrigation and Nitrogen Application on Soil Properties, Soil Bacterial Communities and Agronomic Traits of Cucumber in a Greenhouse System" Plants 12, no. 22: 3834. https://doi.org/10.3390/plants12223834