
Receptor Plants Alleviated Allelopathic Stress from Invasive Chenopodium ambrosioides L. by Upregulating the Production and Autophagy of Their Root Border Cells
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Material Cultivation
2.3. RBC Removal Assay
2.3.1. Root Length Determination
2.3.2. Root Extracellular Trap Area Determination
2.3.3. Root Cap Pectin Methyl Esterase Activity Assay
2.3.4. RBC Count Determination
2.3.5. Transcriptome Analysis
2.4. Cytotoxicity
2.4.1. ROS Content Measurement
2.4.2. RBC Activity Assay
2.4.3. Ultrastructural Observation of RBCs
2.4.4. Autophagy Detection
2.5. Statistical Analysis
3. Results and Analysis
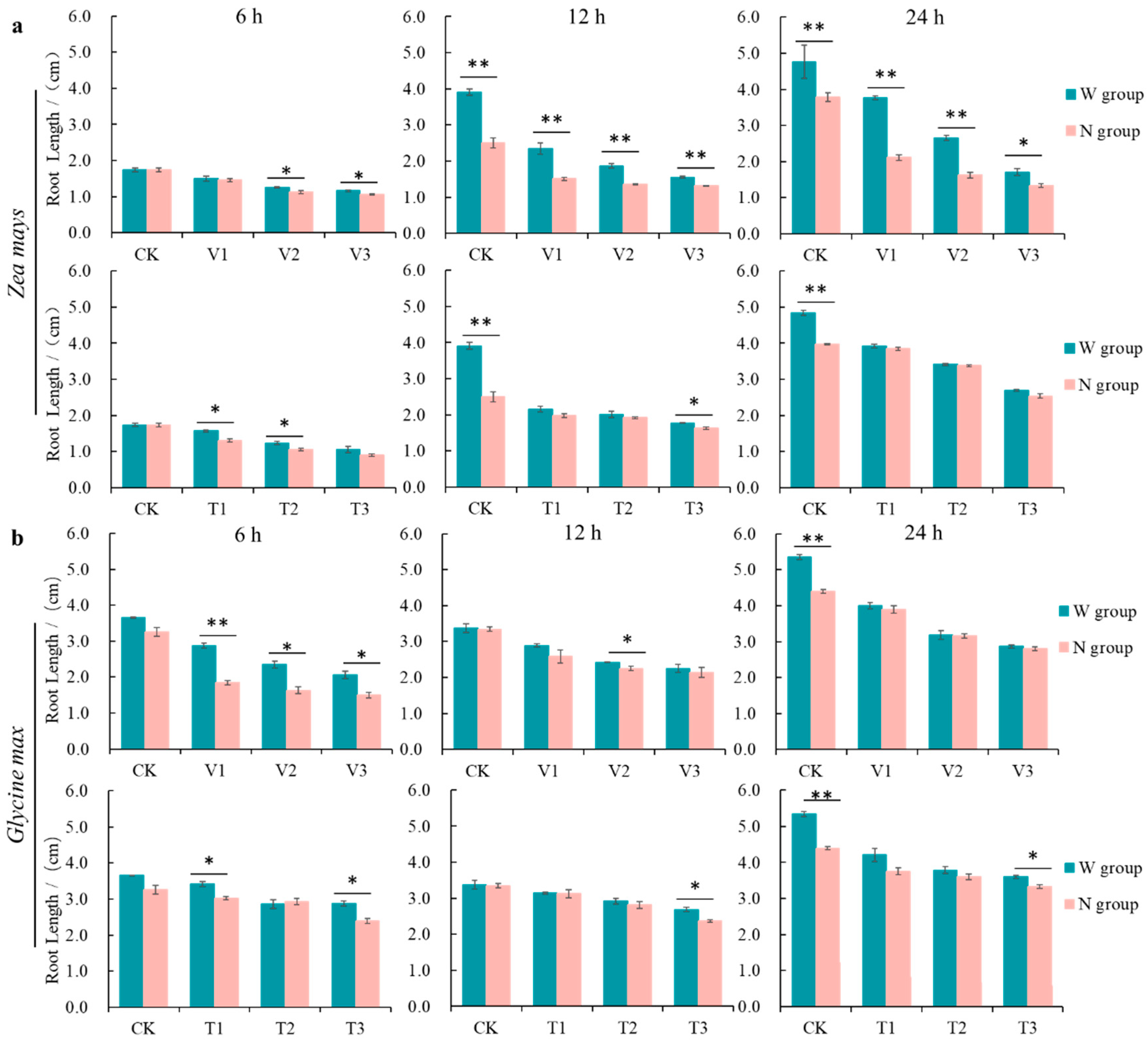
3.1. Effect of RBCs on the Growth of Receptor Roots Induced by the Action of Volatiles from C. ambrosioides
3.2. Changes of Root Extracellular Trap Area in the Receptor Plant by Treatment with C. ambrosioides Volatiles
3.3. Changes of PME Activity in the Receptor Plant by Treatment with C. ambrosioides Volatiles
3.4. Changes in the Number of Receptor RBCs Induced by Allelopathic Stress from C. ambrosioides
3.5. Differentially Expressed Genes Associated with RBC Release
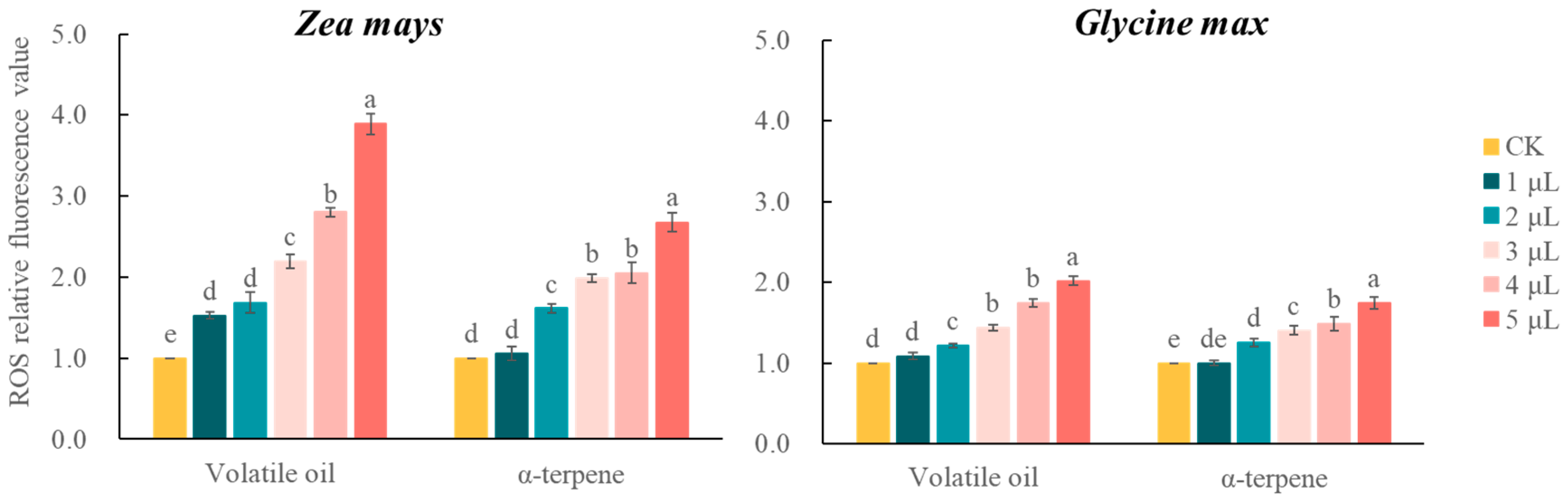
3.6. Effect of C. ambrosioides Volatiles on Cellular Activity and Intracellular ROS Levels in RBCs of Receptor Plant Roots
3.7. Changes in Ultrastructure and Autophagy of Receptor RBCs under Treatment with C. ambrosioides Volatiles
3.7.1. Changes in Ultrastructure
3.7.2. Changes in Autophagy
4. Discussion
4.1. Volatile Allelopathic Stress from C. ambrosioides Alters the Production and Release of Receptor Plant RBCs
4.2. Autophagy Occurs in RBCs under Volatile Stress from C. ambrosioides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, F.; Cheng, Z. Corrigendum: Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2016, 7, 1697. [Google Scholar] [CrossRef] [PubMed]
- Lara-Nuñez, A.; Romero-Romero, T.; Ventura, J.L.; Blancas, V.; Cruz-Ortega, R. Allelochemical stress causes inhibition of growth and oxidative damage in Lycopersicon esculentum Mill. Plant Cell Environ. 2006, 29, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Kumbhar, B.A.; Patel, D.D. Allelopathic effects of different weed species on crop. J. Pharm. Sci. Biosci. Res 2016, 6, 801–805. [Google Scholar]
- Singh, A.A.; Rajeswari, G.; Nirmal, L.A.; Jacob, S. Synthesis and extraction routes of allelochemicals from plants and microbes: A review. Rev. Anal. Chem. 2021, 40, 293–311. [Google Scholar] [CrossRef]
- Hickman, D.T.; Rasmussen, A.; Ritz, K.; Birkett, M.A.; Neve, P. Review: Allelochemicals as mult-kingdom plant defence compounds: Towards an integrated approach. Pest Manag. Sci. 2021, 77, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva, A.A.; Golosova, E.V.; Shelepova, O.V. Allelopathic activity of Acer negundo L. leaf litter as a vector of invasion species into plant communities. BIO Web Conf. EDP Sci. 2021, 38, 00088. [Google Scholar] [CrossRef]
- Limaverde, P.W.; Campina, F.F.; da Cunha, F.A.B.; Crispim, F.D.; Figueredo, F.G.; Lima, L.F.; Oliveira-Tintino, C.D.d.M.; de Matos, Y.M.L.S.; Morais-Braga, M.F.B.; Menezes, I.R.A.; et al. Inhibition of the TetK efflux-pump by the essential oil of Chenopodium ambrosioides L. and α-terpinene against Staphylococcus aureus IS-58. Food Chem. Toxicol. 2017, 109, 957–961. [Google Scholar] [CrossRef]
- Hu, W.; Ma, D.; Wang, Y.; Zhang, H.; Li, Q. Allelopathicpotential of volatile oil from Chenopodium ambrosioides L. on root tip cells of Vicia faba. Shengtai Xuebao/Acta Ecol. Sin. 2011, 31, 3684–3690. [Google Scholar]
- Li, J.; He, Y.; Ma, D.; He, B.; Wang, Y.; Chen, B. Volatile allelochemicals of Chenopodium ambrosioides L. induced mitochondrion-mediated Ca2+-dependent and Caspase-dependent apoptosis signaling pathways in receptor plant cells. Plant Soil 2018, 425, 297–308. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Zhu, X.; Zhang, Z.; Huang, X. Identify potential allelochemicals from Humulus scandens (Lour.) Merr. root extracts that induce allelopathy on Alternanthera philoxeroides (Mart.) Griseb. Sci. Rep. 2021, 11, 1–8. [Google Scholar]
- Lu, Y.; Wang, Y.; Wu, B.; Wang, S.; Wei, M.; Du, D.; Wang, C. Allelopathy of three Compositae invasive alien species on indigenous Lactuca sativa L. enhanced under Cu and Pb pollution. Sci. Hortic. 2020, 267, 109323. [Google Scholar] [CrossRef]
- Wang, Y.N.; Hu, Z.L.; Hao, J.M.; Qian, R.H.; Ren, Q.R.; Ma, D.W. Allelopathic effects of volatile oil and its two main components from Chenopodium ambrosioides L. on maize (Zea mays) root border cells. Allelopath. J. 2019, 46, 71–84. [Google Scholar] [CrossRef]
- Limaverde, P.W.; Figueredo, F.G.; Campina, F.F.; da Cunha, F.A.B.; da Costa, R.H.S.; Pereira, P.S.; Lima, L.F.; de Matos, Y.M.L.S.; Coutinho, H.D.M.; Siqueira-Júnior, J.P.; et al. Inhibition of the essential oil from Chenopodium ambrosioides L. and α-terpinene on the NorA efflux-pump of Staphylococcus aureus. Food Chem. 2018, 262, 72–77. [Google Scholar]
- Ogunleye, G.S.; Fagbohun, O.F.; Babalola, O.O. Chenopodium ambrosioides var. ambrosioides leaf extracts possess regenerative and ameliorative effects against mercury-induced hepatotoxicity and nephrotoxicity. Ind. Crops Prod. 2020, 154, 112723. [Google Scholar] [CrossRef]
- Hawes, M.C.; Gunawardena, U.; Miyasaka, S.; Zhao, X. The role of root border cells in plant defense. Trends Plant Sci. 2000, 5, 128–133. [Google Scholar] [CrossRef]
- Ropitaux, M.; Bernard, S.; Follet-Gueye, M.L.; Vicré, M.; Boulogne, I.; Driouich, A. Xyloglucan and cellulose form molecular cross-bridges connecting root border cells in pea (Pisum sativum). Plant Physiol. Biochem. 2019, 139, 191–196. [Google Scholar] [CrossRef]
- Ropitaux, M.; Bernard, S.; Schapman, D.; Follet-Gueye, M.L.; Vicré, M.; Boulogne, I.; Driouich, A. Root border cells and mucilage secretions of soybean, Glycine max (Merr) L.: Characterization and role in interactions with the oomycete Phytophthora parasitica. Cells 2020, 9, 2215. [Google Scholar] [CrossRef]
- Wen, F.; Curlango-Rivera, G.; Huskey, D.A.; Xiong, Z.; Hawes, M.C. Visualization of extracellular DNA released during border cell separation from the root cap. Am. J. Bot. 2017, 104, 970–978. [Google Scholar] [CrossRef]
- Tran, T.M.; MacIntyre, A.; Hawes, M.; Allen, C. Escaping Underground Nets: Extracellular DNases Degrade Plant Extracellular Traps and Contribute to Virulence of the Plant Pathogenic Bacterium Ralstonia solanacearum. PLoS Pathog. 2016, 12, e1005686. [Google Scholar] [CrossRef]
- Hawes, M.; Allen, C.; Turgeon, B.G.; Curlango-Rivera, G.; Minh Tran, T.; Huskey, D.A.; Xiong, Z. Root border cells and their role in plant defense. Annu. Rev. Phytopathol. 2016, 54, 143–161. [Google Scholar] [CrossRef]
- Brigham, L.A.; Woo, H.H.; Hawes, M.C. Chapter 26 Root Border Cells as Tools in Plant Cell Studies. Methods Cell Biol. 1995, 49, 377–387. [Google Scholar]
- Llugany, M.; Lombini, A.; Poschenrieder, C.; Dinelli, E.; Barceló, J. Different mechanisms account for enhanced copper resistance in Silene armeria ecotypes from mine spoil and serpentine sites. Plant Soil 2003, 251, 55–63. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Y.; Xu, G.; Song, J.; Wu, T.; Mei, X.; Liu, P. Effects of iron toxicity on the morphological and biological characteristics of rice root border cells. J. Plant Nutr. 2017, 40, 332–343. [Google Scholar] [CrossRef]
- Cannesan, M.A.; Durand, C.; Burel, C.; Gangneux, C.; Lerouge, P.; Ishii, T.; Laval, K.; Follet-Gueye, M.L.; Driouich, A.; Vicré-Gibouin, M. Effect of arabinogalactan proteins from the root caps of pea and Brassica napus on Aphanomyces euteiches zoospore chemotaxis and germination. Plant Physiol. 2012, 159, 1658–1670. [Google Scholar] [CrossRef]
- Huskey, D.A.; Curlango-Rivera, G.; Root, R.A.; Wen, F.; Amistadi, M.K.; Chorover, J.; Hawes, M.C. Trapping of lead (Pb) by corn and pea root border cells. Plant Soil 2018, 45430, 205–217. [Google Scholar] [CrossRef]
- Carreras, A.; Bernard, S.; Durambur, G.; Gügi, B.; Loutelier, C.; Pawlak, B.; Boulogne, I.; Vicré, M.; Driouich, A.; Goffner, D.; et al. In vitro characterization of root extracellular trap and exudates of three Sahelian woody plant species. Planta 2020, 251, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gunawardena, U.; Hawes, M.C. Tissue specific localization of root infection by fungal pathogens: Role of root border cells. Mol. Plant-Microbe Interact. 2002, 15, 1128–1136. [Google Scholar] [CrossRef]
- Mravec, J.; Guo, X.; Hansen, A.R.; Schückel, J.; Kračun, S.K.; Mikkelsen, M.D.; Mouille, G.; Johansen, I.E.; Ulvskov, P.; Domozych, D.S.; et al. Pea border cell maturation and release involve complex cell wall structural dynamics. Plant Physiol. 2017, 174, 1051–1066. [Google Scholar] [CrossRef]
- Ma, J.; Feng, X.; Yang, X.; Cao, Y.; Zhao, W.; Sun, L. The leaf extract of crofton weed (Eupatorium adenophorum) inhibits primary root growth by inducing cell death in maize root border cells. Plant Divers. 2020, 42, 174–180. [Google Scholar] [CrossRef]
- Hu, Z.L.; Wang, Y.N.; Ma, D.W.; Chen, B.; He, Y.Q.; Zhou, J. The alleviate effect of extracellular DNA and protein in maize root border cells on the allelochemical stress from Chenopodium ambrosioides L. Sci. Agric. Sin. 2015, 48, 1962–1970. [Google Scholar]
- Chen, B.; Zhou, J.; Gou, X.; Ma, D.W.; Wang, Y.N.; Hu, Z.L.; He, Y.Q. Volatiles from Chenopodium ambrosioides L. induce the oxidative damage in maize (Zea mays L.) radicles. Allelopath. J. 2016, 38, 171–181. [Google Scholar]
- He, S.L.; Gu, C.; Ma, D.W.; Zhang, H. The regulation of signal molecules reactive oxygen species and NO in the induction of root edge cell death in Silkworm beans under allelopathic stress induced by soil mustard. J. Ecol. 2022, 42, 10. [Google Scholar]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The master of bulk and selective recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Wang, P.; Mugume, Y.; Bassham, D.C. New advances in autophagy in plants: Regulation, selectivity and function. Semin. Cell Dev. Biol. 2018, 80, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, J.; Peng, C.; Xu, C.; Yuan, X.; Shi, J. Mechanistic study of programmed cell death of root border cells of cucumber (Cucumber sativus L.) induced by copper. Plant Physiol. Biochem. 2015, 97, 412–419. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, L.; Xu, H. Lysosome calcium in ROS regulation of autophagy. Autophagy 2016, 12, 1954–1955. [Google Scholar] [CrossRef]
- Yang, X.; Bassham, D.C. New insight into the mechanism and function of autophagy in plant cells. Int. Rev. Cell Mol. Biol. 2015, 320, 1–40. [Google Scholar] [PubMed]
- Radwan, M.N.; Morad, M.M.; Ali, M.M.; Wasfy, K.I. Extraction of peppermint volatile oil using a simple constructed steam distillation system. Plant Arch. 2020, 20, 1487–1491. [Google Scholar]
- Mohammed, Y.H.; Ghaidaa, J.M.; Imad, H.H. Analysis of bioactive chemical compounds of Nigella sativa using gas chromatography-mass spectrometry. J. Pharmacogn. Phytother. 2016, 8, 8–24. [Google Scholar] [CrossRef]
- Xie, Y.Y.; Yao, L.X.; Wei, S.; He, S.H.; Chen, L.; Ma, D.W. Allelopathic effects of weed spp on the root border cells of Fagopyrum tataricum (L.) Gaertn. Allelopath. J. 2021, 52, 85–102. [Google Scholar] [CrossRef]
- Xiao, Z.X.; Liang, Y.C. Silicon prevents aluminum from entering root tip by promoting formation of root border cells in rice. Plant Physiol. Biochem. 2022, 175, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Richard, L.; Qin, L.X.; Gadal, P.; Goldberg, R. Molecular cloning and characterisation of a putative pectin methylesterase cDNA in Arabidopsis thaliana (L.). Febs Lett. 1994, 355, 135–139. [Google Scholar] [CrossRef]
- Rasool, F.; Khan, M.R.; Schneider, M.; Uzair, M.; Aqeel, M.; Ajmal, W.; Léon, J.; Naz, A.A. Transcriptome unveiled the gene expression patterns of root architecture in drought-tolerant and sensitive wheat genotypes. Plant Physiol. Biochem. 2022, 178, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.N.; Dong, L.N.; Lü, W. Transcriptome analysis of maize seedling roots in response to nitrogen-, phosphorus-, and potassium deficiency. Plant Soil 2020, 447, 637–658. [Google Scholar] [CrossRef]
- Liu, X.; Liang, W.; Li, Y.X.; Li, C.Y. Transcriptome analysis reveals the effects of alkali stress on root system architecture and endogenous hormones in apple rootstocks. J. Integr. Agric. 2019, 18, 2264–2271. [Google Scholar] [CrossRef]
- Huskey, D.A.; Curlango-Rivera, G.; Hawes, M.C. Use of rhodizonic acid for rapid detection of root border cell trapping of lead and reversal of trapping with DNase. Appl. Plant Sci. 2019, 7, e01240. [Google Scholar] [CrossRef]
- Liu, Q.; Li, K.; Guo, X.; Ma, L.; Guo, Y.; Liu, Z. Developmental characteristics of grapevine seedlings root border cells and their response to ρ-hydroxybenzoic acid. Plant Soil 2019, 443, 199–218. [Google Scholar] [CrossRef]
- Hawes, M.C.; McLain, J.; Ramirez-Andreotta, M.; Curlango-Rivera, G.; Flores-Lara, Y.; Brigham, L.A. Extracellular Trapping of Soil contaminants by root border cells: New insights into plant defense. Agronomy 2016, 6, 5. [Google Scholar] [CrossRef]
- Hawes, M.C.; Curlango-Rivera, G.; Xiong, Z.; Kessler, J.O. Roles of root border cells in plant defense and regulation of rhizosphere microbial populations by extracellular DNA ‘trapping’. Plant Soil 2012, 355, 1–16. [Google Scholar] [CrossRef]
- Park, H.J.; Wang, W.; Curlango-Rivera, G.; Xiong, Z.; Lin, Z.; Huskey, D.A.; Hawes, M.C.; VanEtten, H.D.; Turgeon, B.G. A DNase from a Fungal Phytopathogen Is a Virulence Factor Likely Deployed as Counter Defense against Host-Secreted Extracellular DNA. mBio 2019, 10, e02805-18. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Celoy, R.M.; Nguyen, T.; Zeng, W.; Keegstra, K.; Immerzeel, P.; Pauly, M.; Hawes, M.C. Inducible expression of Pisum sativum xyloglucan fucosyltransferase in the pea root cap meristem, and effects of antisense mRNA expression on root cap cell wall structural integrity. Plant Cell Rep. 2008, 27, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Karve, R.; Suárez-Román, F.; Iyer-Pascuzzi, A.S. The transcription factor NIN-LIKE PROTEIN7 controls border-like cell release. Plant Physiol. 2016, 171, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.C.; Terneus, K.; Hall, Q.; Tan, L.; Wang, Y.; Wegenhart, B.L.; Chen, L.; Lamport, D.T.; Chen, Y.; Kieliszewski, M.J. Self-assembly of the plant cell wall requires an extensin scaffold. Proc. Natl. Acad. Sci. USA 2008, 105, 2226–2231. [Google Scholar] [CrossRef] [PubMed]
- Do, B.C.; Dang, T.T.; Berrin, J.G.; Haltrich, D.; To, K.A.; Sigoillot, J.C.; Yamabhai, M. Cloning, expression in Pichia pastoris, and characterization of a thermostable GH5 mannan endo-1, 4-β-mannosidase from Aspergillus niger BK01. Microb. Cell Factories 2009, 8, 1–12. [Google Scholar]
- Wang, H.; Jin, Y.; Wang, C.; Li, B.; Jiang, C.; Sun, Z.; Zhang, Z.; Kong, F.; Zhang, H. Fasciclin-like arabinogalactan proteins, PtFLAs, play important roles in GA-mediated tension wood formation in Populus. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Du, J.; Anderson, C.T.; Xiao, C. Dynamics of pectic homogalacturonan in cellular morphogenesis and adhesion, wall integrity sensing and plant development. Nat. Plants 2022, 8, 332–340. [Google Scholar] [CrossRef]
- Durand, C.; Vicré-Gibouin, M.; Follet-Gueye, M.L.; Duponchel, L.; Moreau, M.; Lerouge, P.; Driouich, A. The organization pattern of root border-like cells of Arabidopsis is dependent on cell wall homogalacturonan. Plant Physiol. 2009, 150, 1411–1421. [Google Scholar] [CrossRef]
- Mravec, J.; Kračun, S.K.; Rydahl, M.G.; Westereng, B.; Miart, F.; Clausen, M.H.; Fangel, J.U.; Daugaard, M.; Van Cutsem, P.; De Fine Licht, H.H.; et al. Tracking developmentally regulated post-synthetic processing of homogalacturonan and chitin using reciprocal oligosaccharide probes. Development 2014, 141, 4841–4850. [Google Scholar] [CrossRef]
- Araniti, F.; Costas-Gil, A.; Cabeiras-Freijanes, L.; Lupini, A.; Sunseri, F.; Reigosa, M.J.; Abenavoli, M.R.; Sánchez-Moreiras, A.M. Rosmarinic acid induces programmed cell death in Arabidopsis seedlings through reactive oxygen species and mitochondrial dysfunction. PLoS ONE 2018, 13, e0208802. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Mahdavikia, F.; Saharkhiz, M.J.; Karami, A. Defensive response of radish seedlings to the oxidative stress arising from phenolic compounds in the extract of peppermint (Mentha× piperita L.). Sci. Hortic. 2017, 214, 133–140. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Madkour, L.H. Oxidative stress and oxidative damage-induced cell death. In Reactive Oxygen Species (ROS), Nanoparticles, and Endoplasmic Reticulum (ER) Stress-Induced Cell Death Mechanisms; Academic Press: Cambridge, MA, USA, 2020; Volume 8, pp. 175–197. [Google Scholar]
- Jiang, X.S.; Chen, X.M.; Wan, J.M.; Gui, H.B.; Ruan, X.Z.; Du, X.G. Autophagy protects against palmitic acid-induced apoptosis in podocytes in vitro. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Xia, F.N.; Xie, L.J.; Yu, L.J.; Chen, Q.F.; Zhuang, X.H.; Wang, Q.; Li, F.; Jiang, L.; Xie, Q.; et al. TRAF family proteins regulate autophagy dynamics by modulating AUTOPHAGY PROTEIN6 stability in Arabidopsis. Plant Cell 2017, 29, 890–911. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Jikumaru, Y.; Kamiya, Y.; Kusano, M.; Consonni, C.; Panstruga, R.; Ohsumi, Y.; Shirasu, K. Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in Arabidopsis. Plant Cell 2009, 21, 2914–2927. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor Plants | Treatment | Comparisons | ||
|---|---|---|---|---|
| Control | Volatile Oil | α-Terpinene | ||
| Zea mays | ZCKN ZCKW | ZVN | ZTN | ZCKN-vs-ZCKW |
| ZCKN-vs-ZTN | ||||
| ZCKN-vs-ZVN | ||||
| Glycine max | GCKN GCKW | GVN | GTN | GCKN-vs-GCKW |
| GCKN-vs-GTN | ||||
| GCKN-vs-GVN | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhou, X.; He, S.; Wang, W.; Ma, D.; Wang, Y.; Zhang, H. Receptor Plants Alleviated Allelopathic Stress from Invasive Chenopodium ambrosioides L. by Upregulating the Production and Autophagy of Their Root Border Cells. Plants 2023, 12, 3810. https://doi.org/10.3390/plants12223810
Wang Q, Zhou X, He S, Wang W, Ma D, Wang Y, Zhang H. Receptor Plants Alleviated Allelopathic Stress from Invasive Chenopodium ambrosioides L. by Upregulating the Production and Autophagy of Their Root Border Cells. Plants. 2023; 12(22):3810. https://doi.org/10.3390/plants12223810
Chicago/Turabian StyleWang, Qiang, Xijie Zhou, Shengli He, Wenguo Wang, Danwei Ma, Yu Wang, and Hong Zhang. 2023. "Receptor Plants Alleviated Allelopathic Stress from Invasive Chenopodium ambrosioides L. by Upregulating the Production and Autophagy of Their Root Border Cells" Plants 12, no. 22: 3810. https://doi.org/10.3390/plants12223810