
Suspension Cell Culture of Polyscias fruticosa (L.) Harms in Bubble-Type Bioreactors—Growth Characteristics, Triterpene Glycosides Accumulation and Biological Activity
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Suspension Cell Culture Growth in Flasks
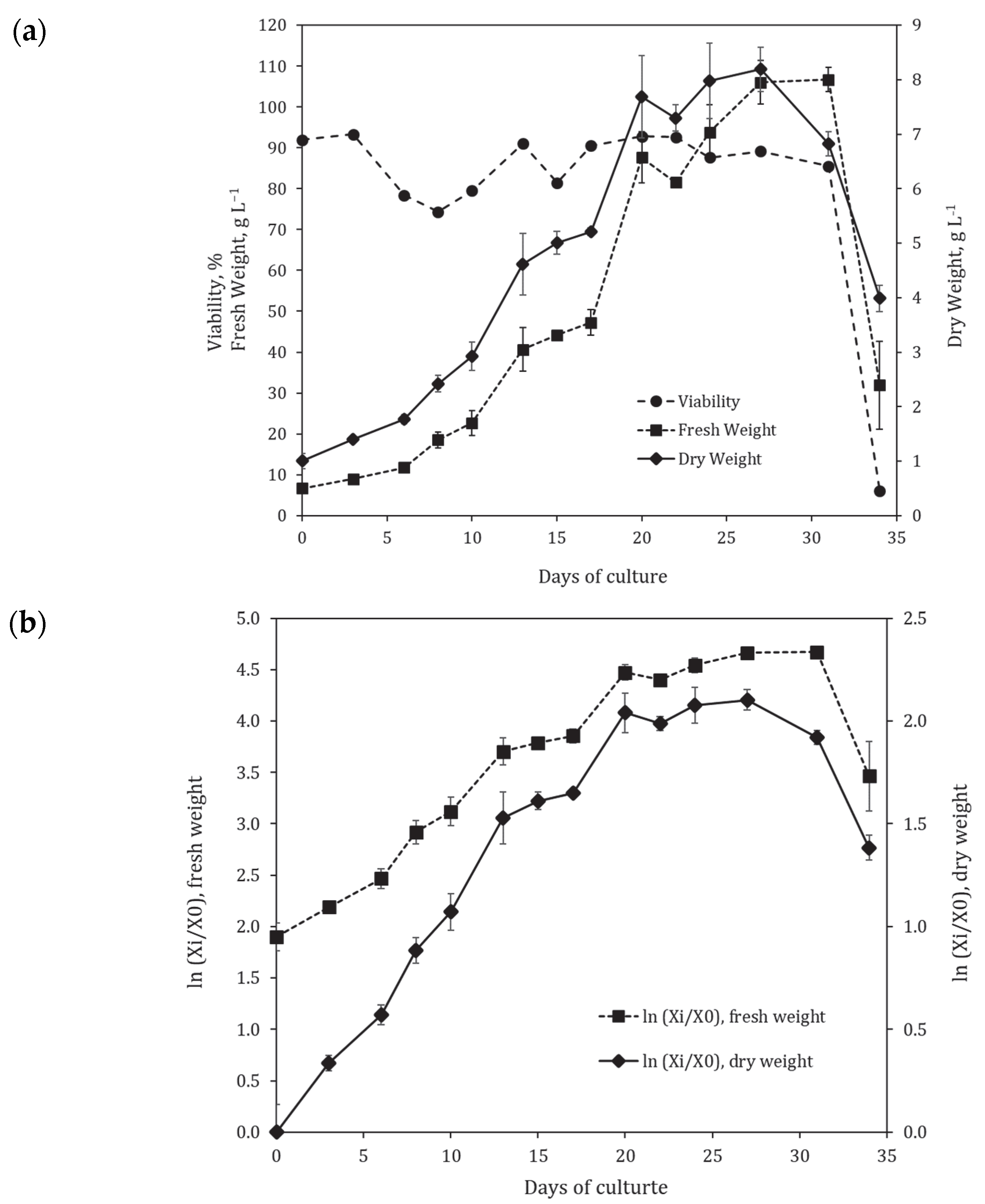
2.2. Suspension Cell Culture Growth in 20 L Bioreactor under a Semi-Continuous Cultivation Regime
2.3. Analysis of the Two Major Secondary Metabolites in the Suspension Cell Culture of Polyscias fruticosa during Cultivation in Flasks and Bioreactors
2.4. Antioxidant and Antimicrobial Activities of the Extracts from Cell Biomass of P. fruticosa Grown in Bioreactors
3. Discussion
4. Materials and Methods
4.1. Cell Suspension Cultivation in Flasks and Bioreactors
4.2. Assessment of Growth and Physiological Characteristics of the Cell Suspension Culture
4.3. Extract Preparation and High-Performance Liquid Chromatography-Electrospray Ionization–Mass Spectrometry (UPLC-ESI-MS) Analysis of Triterpene Glycosides in Cell Biomass
4.4. Test Systems for Rapid Assessment of the Biological Activity of Cell Biomass Extracts: Antioxidant and Antimicrobial Tests
4.4.1. Preparation of Extracts from Plant Leaves and Cell Culture Biomass for Antioxidant and Antimicrobial Activity Tests
4.4.2. Determination of Antioxidant (Antiradical) Activities Using DPPH and TEAC Assays
DPPH Assay
TEAC Assay
Estimation of the Effective Concentration (EC50)
4.4.3. Determination of Antibacterial Activities
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ashmawy, N.S.; Gad, H.A.; Ashour, M.L.; El-Ahmady, S.H.; Singab, A.N.B. The Genus Polyscias (Araliaceae): A Phytochemical and Biological Review. J. Herb. Med. 2020, 23, 100377. [Google Scholar] [CrossRef]
- Thach, B.D.; Le Nguyen Tu Linh, D.T.; Cang, T.T.B.; Giang, T.T.L.; Uyen, N.P.A.; Suong, N.K. Protocol Establishment for Multiplication and Regeneration of Polyscias fruticosa L. Harms. an Important Medicinal Plant in Vietnam. Eur. J. Biotechnol. Genet. Eng. 2016, 3, 31–37. [Google Scholar]
- Pandya, D.; Mankad, A.; Pandya, H. Cost Effective Micropropagation of Polyscias fruicosa (L.) Harm. Int. Assoc. Biol. Comput. Dig. 2022, 1, 58–62. [Google Scholar] [CrossRef]
- Nguyen, T.-T.-D.; Nguyen, Q.-D.; Nguyen, T.-V.-L. Kinetic Study on Chlorophyll and Antioxidant Activity from Polyscias fruticosa (L.) Harms Leaves via Microwave-Assisted Extraction. Molecules 2021, 26, 3761. [Google Scholar] [CrossRef] [PubMed]
- Lutomski, J.; Luận, T.C.; Hoa, T.T. Polyacetylenes in the Araliaceae Family. Part IV. Herba Pol. 1992, 38, 137–140. [Google Scholar]
- Tram, N.T.T.; Tuyet, H.D.; Minh, Q.N. One Unusual Sterol from Polyscias fruticosa (L.) Harms (Araliaceae). Can Tho Univ. J. Sci. 2017, 7, 33–36. [Google Scholar]
- Huan, V.D.; Yamamura, S.; Ohtani, K.; Kasai, R.; Yamasaki, K.; Nham, N.T.; Chau, H.M. Oleanane Saponins from Polyscias fruticosa. Phytochemistry 1998, 47, 451–457. [Google Scholar] [CrossRef]
- Tran, V.T.; Tran, T.H.H.; Nguyen, T.D. Validated High Performance Liquid Chromatography Method for Quantification of a Major Saponin in Polyscias fruticosa. J. Multidiscip. Eng. Sci. Technol. 2016, 3, 4880–4882. [Google Scholar]
- Do, V.M.; Tran, C.L.; Nguyen, T.P. Polysciosides J and K, Two New Oleanane-Type Triterpenoid Saponins from the Leaves of Polyscias fruticosa (L.) Harms. Cultivating in an Giang Province, Viet Nam. Nat. Prod. Res. 2020, 34, 1250–1255. [Google Scholar] [CrossRef]
- Koffuor, G.A.; Boye, A.; Ofori-Amoah, J.; Kyei, S.; Abokyi, S.; Nyarko, R.A.; Naalukyem, B.R. Anti-Inflammatory and Safety Assessment of Polyscias fruticosa (L.) Harms (Araliaceae) Leaf Extract in Ovalbumin-Induced Asthma. J. Phytopharm. 2014, 3, 337–342. [Google Scholar] [CrossRef]
- Divakar, M.C.; Bensita, M.B. Screening of Various Leaf Extracts of Polyscias fruticosa Harms for Their Antidiabetic Activity. Indian J. Nat. Prod. 1998, 14, 24–28. [Google Scholar]
- Varadharajan, R.; Rajalingam, D. Diuretic Activity of Polyscias fruticosa (L.) Harms. Int. J. Innov. Drug Discov. 2011, 1, 15–18. [Google Scholar]
- Mawea, F.; Indrayati, A.; Rahmawati, I. Review: Antibacterial Activity of Polyscias fruticosa and Molecular Mechanism of Active Compounds. Int. J. Health Med. Curr. Res. 2021, 6, 2062–2087. [Google Scholar] [CrossRef]
- Bensita, M.B.; Nilani, P.; Sandhya, S.M. Studies on the Adaptogenic and Antibacterial Properties of Polyscias fructicosa (L) Harms. Anc. Sci. Life 1999, 18, 231–246. [Google Scholar] [PubMed]
- Luyen, N.T.; Dang, N.H.; Binh, P.T.X.; Hai, N.T.; Dat, N.T. Hypoglycemic Property of Triterpenoid Saponin PFS Isolated from Polyscias fruticosa Leaves. An. Acad. Bras. Ciências 2018, 90, 2881–2886. [Google Scholar] [CrossRef]
- Tran, T.H.H.; Nguyen, H.D.; Nguyen, T.D. α-Amylase and α -Glucosidase Inhibitory Saponins from Polyscias fruticosa Leaves. J. Chem. 2016, 2016, 2082946. [Google Scholar] [CrossRef]
- Le, Q.-U. A Science Opinion on Polyscias fruticosa and Morus alba L. Combination: Better Anti-Diabetic and Late Complication Inhibitory Properties? Curr. Res. Diabetes Obes. J. 2019, 10, 555798. [Google Scholar] [CrossRef]
- Asumeng Koffuor, G.; Boye, A.; Kyei, S.; Ofori-Amoah, J.; Akomanin Asiamah, E.; Barku, A.; Acheampong, J.; Amegashie, E.; Kumi Awuku, A. Anti-Asthmatic Property and Possible Mode of Activity of an Ethanol Leaf Extract of Polyscias fruticosa. Pharm. Biol. 2016, 54, 1354–1363. [Google Scholar] [CrossRef]
- Yen, T.T. Improvement of Learning Ability in Mice and Rats with the Root Extract of Dinh Lang (Policias fruticosum L.). Acta Physiol. Hung. 1990, 75, 69–76. [Google Scholar]
- Yen, T.T. Stimulation of Sexual Performance in Male Rats with the Root Extract of Dinh Lang (Policias fruticosum L.). Acta Physiol. Hung. 1990, 75, 61–67. [Google Scholar]
- Boye, A.; OseiOwusu, A.; Koffuor, G.; Barku, V.; Asiamah, E.; Asante, E. Assessment of Polyscias fruticosa (L.) Harm (Araliaceae) Leaf Extract on Male Fertility in Male Wistar Rats. J. Intercult. Ethnopharmacol. 2018, 7, 45–56. [Google Scholar] [CrossRef]
- Yen, T.T.; Knoll, J. Extension of Lifespan in Mice Treated with Dinh Lang (Policias fruticosum L.) and (−)Deprenyl. Acta Physiol. Hung. 1992, 79, 119–124. [Google Scholar] [PubMed]
- Selvaraj, B.; Le, T.T.; Kim, D.W.; Jung, B.H.; Yoo, K.-Y.; Ahn, H.R.; Thuong, P.T.; Tran, T.T.T.; Pae, A.N.; Jung, S.H.; et al. Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells. Int. J. Mol. Sci. 2023, 24, 3969. [Google Scholar] [CrossRef]
- Ly, H.T.; Nguyen, T.T.H.; Le, V.M.; Lam, B.T.; Mai, T.T.T.; Dang, T.P.T. Therapeutic Potential of Polyscias fruticosa (L.) Harms Leaf Extract for Parkinson’s Disease Treatment by Drosophila Melanogaster Model. Oxidative Med. Cell. Longev. 2022, 2022, 5262677. [Google Scholar] [CrossRef] [PubMed]
- Nhu-Trang, T.-T.; Phan, T.-K.-T.; Lam, T.-M.-T.; Nguyen, P.-B.-D.; Nguyen, T.-V.-L. Drying Kinetics and Energy Consumption in Hot-Air and Microwave Drying of Polyscias fruticosa (L.) Harms Leaves. AIP Conf. Proc. 2022, 2610, 070004. [Google Scholar]
- MinhThu, N.; Son, H.L. Effect of Storage Temperature and Preservatives on the Stability and Quality of Polyscias fruticosa (L.) Harms Herbal Health Drinks. J. Pharm. Res. Int. 2019, 26, 1–7. [Google Scholar] [CrossRef]
- Tran, C.H.; Nguyen, H.A.; Nguyen, T.N.T.; Ha, T.T.N.; Le, T.H.A. Effects of Storage Conditions on Polyphenol and Triterpenoid Saponin Content and the Antioxidant Capacity of Ethanolic Extract from Leaves of Polyscias fruticosa (L.) Harms. J. Sci. Technol. Food 2020, 3, 47–53. [Google Scholar]
- Nguyen, M.P. Impact of Roasting to Total Phenolic, Flavonoid and Antioxidant Activities in Root, Bark and Leaf of Polyscias fruticosa. J. Pharm. Res. Int. 2020, 32, 13–17. [Google Scholar] [CrossRef]
- Sakr, S.S.; Melad, S.S.; El-Shamy, M.A.; Elhafez, A.E. In Vitro Propagation of Polyscias fruticosa Plant. Int. J. Plant Soil Sci. 2014, 3, 1254–1265. [Google Scholar] [CrossRef]
- Vinh, D.T.; Phuong Hoa, M.T.; Nhu Thao, L.T.; Trang Nha, N.H.; Minh, T. Van Saponin Production by Cell Culture Techniques of Polyscias fruticosa L. Harms. Acad. J. Biol. 2015, 37, 135–141. [Google Scholar] [CrossRef]
- Kim, P.T.A.; An, L.T.B.; Chung, N.T.; Truong, N.T.; Thu, L.T.A.; Huan, L.V.T.; Loc, N.H. Growth and Oleanolic Acid Accumulation of Polyscias fruticosa Cell Suspension Cultures. Curr. Pharm. Biotechnol. 2021, 22, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Phuong, T.T.B.; Trung, V.P.; An, N.H.; Tuan, N.D.; Nguyen, P.T.T. The Effects of 2,4-Dichlorophenoxyacetic Acid and α-Naphthaleneacetic Acid on Biomass Increment, Rhizogenesis and Somatic Embryogenesis of Suspension-Cultured Dinh Lang Cells [Polyscias fruticosa (L.) Harms]. Indian J. Agric. Res. 2021, 56, 70–75. [Google Scholar] [CrossRef]
- Hau, N.T.; Nhu Thao, L.T.; Minh, T. Van Cultivation of Leaf-Tissue of Polyscias fruticosa (L.) Harms for Quantity of Saponin Accumulation. Acad. J. Biol. 2015, 37, 184–189. [Google Scholar] [CrossRef]
- Lộc, P.V. A Study on Adventitious Root Formation of Ming Aralia (Polyscias fruticosa L. Harms) by in Vitro Culture. Univ. Danang J. Sci. Technol. 2014, 3, 106–108. [Google Scholar]
- Titova, M.V.; Popova, E.V.; Konstantinova, S.V.; Kochkin, D.V.; Ivanov, I.M.; Klyushin, A.G.; Titova, E.G.; Nebera, E.A.; Vasilevskaya, E.R.; Tolmacheva, G.S.; et al. Suspension Cell Culture of Dioscorea deltoidea—A Renewable Source of Biomass and Furostanol Glycosides for Food and Pharmaceutical Industry. Agronomy 2021, 11, 394. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Weber, J. Bioreactors for Plant Cells: Hardware Configuration and Internal Environment Optimization as Tools for Wider Commercialization. Biotechnol. Lett. 2014, 36, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Valdiani, A.; Hansen, O.K.; Nielsen, U.B.; Johannsen, V.K.; Shariat, M.; Georgiev, M.I.; Omidvar, V.; Ebrahimi, M.; Tavakoli Dinanai, E.; Abiri, R. Bioreactor-Based Advances in Plant Tissue and Cell Culture: Challenges and Prospects. Crit. Rev. Biotechnol. 2019, 39, 20–34. [Google Scholar] [CrossRef]
- Thanh, N.T.; Murthy, H.N.; Paek, K.Y. Optimization of Ginseng Cell Culture in Airlift Bioreactors and Developing the Large-Scale Production System. Ind. Crops Prod. 2014, 60, 343–348. [Google Scholar] [CrossRef]
- Kubica, P.; Szopa, A.; Kokotkiewicz, A.; Miceli, N.; Taviano, M.F.; Maugeri, A.; Cirmi, S.; Synowiec, A.; Gniewosz, M.; Elansary, H.O.; et al. Production of Verbascoside, Isoverbascoside and Phenolic Acids in Callus, Suspension, and Bioreactor Cultures of Verbena officinalis and Biological Properties of Biomass Extracts. Molecules 2020, 25, 5609. [Google Scholar] [CrossRef]
- Kochkin, D.V.; Sukhanova, E.S.; Nosov, A.M. The Accumulation of Triterpene Glycosides in the Growh Cycle of Cell Suspension Cultures of Polyscias Fruticosa. Vestn. Volga State Univ. Technol. Ser For. Ecol. Nat. Manag. 2014, 4, 67–73. [Google Scholar]
- Kochkin, D.V.; Sukhanova, E.S.; Nosov, A.M. Triterpene Glycosides in Suspension-Cell Culture of Polyscias fruticosa (L.) Harms. In Proceedings of the International Conference with Elements of Scientific School for Young Scientists “Perspectives of Phytobiotechnology to Improving the Quality of Life in the North”, October 10–16, 2010; North-Eastern Federal University in Yakutsk: Yakutsk, Russia, 2020; pp. 201–204. [Google Scholar]
- Kochkin, D.V.; Sukhanova, E.S.; Sergeev, R.V.; Nosov, A.M. Triterpene Glycosides of Polyscias spp. Cell Cultures. Vestn. Volga State Univ. Technol. Ser For. Ecol. Nat. Manag. 2014, 1, 69–76. [Google Scholar]
- Sukhanova, E.S.; Chernyak, N.D.; Nosov, A.M. Obtaining and Description of Polyscias filicifolia and Polyscias fruticosa Calli and Suspension Cell Cultures. Biotekhnologiia 2010, 4, 44–50. [Google Scholar]
- Sukhanova, E.S.; Kochkin, D.V.; Titova, M.V.; Nosov, A.M. Growth and Biosynthetic Characteristics of Different Polyscias Plant Cell Culture Strains. Vestn. Volga State Univ. Technol. Ser For. Ecol. Nat. Manag. 2012, 2, 57–66. [Google Scholar]
- Yang, W.; Ye, M.; Qiao, X.; Liu, C.; Miao, W.; Bo, T.; Tao, H.; Guo, D. A Strategy for Efficient Discovery of New Natural Compounds by Integrating Orthogonal Column Chromatography and Liquid Chromatography/Mass Spectrometry Analysis: Its Application in Panax Ginseng, Panax Quinquefolium and Panax Notoginseng to Characterize 4. Anal. Chim. Acta 2012, 739, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.-T.; Liang, X.-L.; Zhu, H.-Y.; Peng, X.-L.; Guo, X.-J.; Zhao, L.-S. Rapid Separation and Identification of 31 Major Saponins in Shizhu Ginseng by Ultra-High Performance Liquid Chromatography–Electron Spray Ionization–MS/MS. J. Ginseng Res. 2016, 40, 220–228. [Google Scholar] [CrossRef]
- Huang, T.-K.; McDonald, K.A. Bioreactor Engineering for Recombinant Protein Production in Plant Cell Suspension Cultures. Biochem. Eng. J. 2009, 45, 168–184. [Google Scholar] [CrossRef]
- Motolinía-Alcántara, E.A.; Castillo-Araiza, C.O.; Rodríguez-Monroy, M.; Román-Guerrero, A.; Cruz-Sosa, F. Engineering Considerations to Produce Bioactive Compounds from Plant Cell Suspension Culture in Bioreactors. Plants 2021, 10, 2762. [Google Scholar] [CrossRef]
- Su, R.; Sujarani, M.; Shalini, P.; Prabhu, N. A Review on Bioreactor Technology Assisted Plant Suspension Culture. Asian J. Biotechnol. Bioresour. Technol. 2019, 5, 1–13. [Google Scholar] [CrossRef]
- Esperança, M.N.; Mendes, C.E.; Rodriguez, G.Y.; Cerri, M.O.; Béttega, R.; Badino, A.C. Sparger Design as Key Parameter to Define Shear Conditions in Pneumatic Bioreactors. Biochem. Eng. J. 2020, 157, 107529. [Google Scholar] [CrossRef]
- Seidel, S.; Maschke, R.W.; Werner, S.; Jossen, V.; Eibl, D. Oxygen Mass Transfer in Biopharmaceutical Processes: Numerical and Experimental Approaches. Chem. Ing. Tech. 2021, 93, 42–61. [Google Scholar] [CrossRef]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor Scale-up and Oxygen Transfer Rate in Microbial Processes: An Overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef]
- Garcia-Ochoa, F.; Gomez, E.; Santos, V.E. Fluid Dynamic Conditions and Oxygen Availability Effects on Microbial Cultures in STBR: An Overview. Biochem. Eng. J. 2020, 164, 107803. [Google Scholar] [CrossRef]
- Kieran, P.; MacLoughlin, P.; Malone, D. Plant Cell Suspension Cultures: Some Engineering Considerations. J. Biotechnol. 1997, 59, 39–52. [Google Scholar] [CrossRef]
- Lee, C.W.; Shuler, M.L. The Effect of Inoculum Density and Conditioned Medium on the Production of Ajmalicine and Catharanthine from Immobilized Catharanthus roseus Cells. Biotechnol. Bioeng. 2000, 67, 61–71. [Google Scholar] [CrossRef]
- Sakurai, M.; Mori, T.; Seki, M.; Furusaki, S. Changes of Anthocyanin Composition by Conditioned Medium and Cell Inoculum Size Using Strawberry Suspension Culture. Biotechnol. Lett. 1996, 18, 1149–1154. [Google Scholar] [CrossRef]
- Thanh, N.T.; Murthy, H.N.; Yu, K.W.; Jeong, C.S.; Hahn, E.J.; Paek, K.Y. Effect of Inoculum Size on Biomass Accumulation and Ginsenoside Production by Large-Scale Cell Suspension Cultures of Panax Ginseng. J. Plant Biotechnol. 2004, 6, 265–268. [Google Scholar]
- Tan, S.H.; Maziah, M.; Ariff, A. Synergism Effect between Inoculum Size and Aggregate Size on Flavonoid Production in Centella asiatica (L.) Urban (Pegaga) Cell Suspension Cultures. Int. J. Res. Eng. Technol. 2013, 2, 244–253. [Google Scholar]
- Georgiev, M.I.; Eibl, R.; Zhong, J.-J. Hosting the Plant Cells in Vitro: Recent Trends in Bioreactors. Appl. Microbiol. Biotechnol. 2013, 97, 3787–3800. [Google Scholar] [CrossRef]
- Titova, M.V.; Popova, E.V.; Shumilo, N.A.; Kulichenko, I.E.; Chernyak, N.D.; Ivanov, I.M.; Klushin, A.G.; Nosov, A.M. Stability of Cryopreserved Polyscias filicifolia Suspension Cell Culture during Cultivation in Laboratory and Industrial Bioreactors. Plant Cell Tissue Organ Cult. 2021, 145, 591–600. [Google Scholar] [CrossRef]
- Demidova, E.; Globa, E.; Klushin, A.; Kochkin, D.; Nosov, A. Effect of Methyl Jasmonate on the Growth and Biosynthesis of C13- and C14-Hydroxylated Taxoids in the Cell Culture of Yew (Taxus wallichiana Zucc.) of Different Ages. Biomolecules 2023, 13, 969. [Google Scholar] [CrossRef]
- Titova, M.V.; Reshetnyak, O.V.; Osipova, E.A.; Osip’yants, A.I.; Shumilo, N.A.; Oreshnikov, A.V.; Nosov, A.M. Submerged Cultivation of Stephania glabra (Roxb.) Miers Cells in Different Systems: Specific Features of Growth and Accumulation of Alkaloid Stepharine. Appl. Biochem. Microbiol. 2012, 48, 645–649. [Google Scholar] [CrossRef]
- Nguyen, N.Q.; Nguyen, M.T.; Nguyen, V.T.; Le, V.M.; Trieu, L.H.; Le, X.T.; Khang, T.V.; Giang, N.T.; Thach, N.Q.; Hung, T.T. The Effects of Different Extraction Conditions on the Polyphenol, Flavonoids Components and Antioxidant Activity of Polyscias fruticosa Roots. IOP Conf. Ser. Mater. Sci. Eng. 2020, 736, 022067. [Google Scholar] [CrossRef]
- Nguyen, T.T.H. Studies on Antioxidant Activity of Polyscias fruticosa Harms. (Araliaceae). J. Med. Mater. Hanoi. 2003, 142–146. [Google Scholar]
- Nguyen, M.P. Changes of Phytochemical, Antioxidant Characteristics of Polyscias fruticosa Rhizomes during Convective and Freeze Drying. Biosci. Res. 2020, 17, 323–326. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Titova, M.V.; Shumilo, N.A.; Kulichenko, I.E.; Ivanov, I.M.; Sukhanova, E.S.; Nosov, A.M. Features of Respiration and Formation of Steroidal Glycosides in Dioscorea deltoidea Cell Suspension Culture Grown in Flasks and Bioreactors. Russ. J. Plant Physiol. 2015, 62, 557–563. [Google Scholar] [CrossRef]
- Dixon, R.A.; Gonzales, R.A. (Eds.) Plant Cell Culture: A Practical Approach, 2nd ed.; Practical Approach Series, 145; Oxford University Press: New York, NY, USA, 1995. [Google Scholar]
- Pirt, S.J. Principles of Microbe and Cell Cultivation; Blackwell Scientific: Oxford, UK, 1975; ISBN 0632081503. [Google Scholar]
- Kochkin, D.V.; Galishev, B.A.; Glagoleva, E.S.; Titova, M.V.; Nosov, A.M. Rare Triterpene Glycoside of Ginseng (Ginsenoside Malonyl-Rg1) Detected in Plant Cell Suspension Culture of Panax japonicus Var. Repens. Russ. J. Plant Physiol. 2017, 64, 649–656. [Google Scholar] [CrossRef]
- Masci, A.; Mattioli, R.; Costantino, P.; Baima, S.; Morelli, G.; Punzi, P.; Giordano, C.; Pinto, A.; Donini, L.M.; D’Erme, M.; et al. Neuroprotective Effect of Brassica Oleracea Sprouts Crude Juice in a Cellular Model of Alzheimer’s Disease. Oxid. Med. Cell. Longev. 2015, 2015, 781938. [Google Scholar] [CrossRef]
- Orlova, A.; Kysil, E.; Tsvetkova, E.; Meshalkina, D.; Whaley, A.; Whaley, A.O.; Laub, A.; Francioso, A.; Babich, O.; Wessjohann, L.A.; et al. Phytochemical Characterization of Water Avens (Geum rivale L.) Extracts: Structure Assignment and Biological Activity of the Major Phenolic Constituents. Plants 2022, 11, 2859. [Google Scholar] [CrossRef]
- CLSI Standard M07; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. 11th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Cultivation System | ||||
|---|---|---|---|---|---|
| 250 mL Flasks | 20 L Bioreactors * | ||||
| MC 1 | MC 2 | MC 3 | Average of MC 1–3 | ||
| X0, g L−1 | 1.10 ± 0.45 | 2.40 ± 0.37 | 1.97 ± 0.10 | 1.83 ± 0.27 | 2.07 ± 0.30 |
| ∆tmax, days | 22.5 ± 3.5 a | 15.0 ± 2.5 b | 16.5 ± 4.0 ab | 18.5 ± 4.5 ab | 16.7 ± 2.0 |
| Xmax, g L−1 | 8.19 ± 0.81 a | 7.31 ± 1.43 a | 6.31 ± 0.34 a | 6.76 ± 0.81 a | 6.79 ± 0.50 |
| μ, day−1 | 0.21 ± 0.03 a | 0.10 ± 0.02 b | 0.13 ± 0.03 b | 0.11 ± 0.02 b | 0.11 ± 0.03 |
| τ, day | 3.3 ± 0.4 b | 6.9 ± 1.2 a | 5.3 ± 1.6 ab | 6.3 ± 1.3 a | 6.3 ± 1.4 |
| Y | 0.27 ± 0.05 a | 0.16 ± 0.05 b | 0.14 ± 0.01 b | 0.16 ± 0.03 b | 0.16 ± 0.01 |
| Pmax, g (L · day)−1 | 0.45 ± 0.12 a | 0.33 ± 0.06 ab | 0.30 ± 0.05 b | 0.35 ± 0.15 ab | 0.32 ± 0.13 |
| Day of the Growth Cycle | Number of Cells in the Aggregate | |||||
|---|---|---|---|---|---|---|
| 1–5 | 6–10 | 11–20 | 21–30 | 31–50 | >50 | |
| % of Aggregates Consisted of a Given Number of Cells | ||||||
| 250 mL flasks | ||||||
| Day 10 | 4.3 ± 2.1 a | 12.0 ± 2.0 a | 17.0 ± 1.0 a | 15.0 ± 6.2 a | 24.3 ± 3.2 a | 27.6 ± 5.5 b |
| Day 20 | 2.3 ± 0.6 a | 5.7 ± 1.5 b | 13.0 ± 1.7 ab | 17.7 ± 8.4 a | 29.0 ± 4.3 a | 33.0 ± 6.9 ab |
| 20 L bioreactors | ||||||
| Day 10 | 4.0 ± 1.7 a | 7.7 ± 1.5 b | 10.0 ± 3.6 b | 15.4 ± 7.4 a | 25.3 ± 7.6 a | 39.3 ± 2.9 a |
| tR, min * | [M-H]−, m/z ** | Identification Results |
|---|---|---|
| 7.906 | 955.6 | PFS *** |
| 13.440 | 793.6 | Ladyginoside A (LadA) |
| Cultivation System | Variant | Triterpene Glycoside Content, mg gDW−1 | |
|---|---|---|---|
| PFS | Ladyginoside A | ||
| Flasks | Day 20 | 0.50 ± 0.05 bc | 0.22 ± 0.01 b |
| Day 27 | 0.34 ± 0.12 c | 0.20 ± 0.01 b | |
| Bioreactors * | MC no. 1 | 0.78 ± 0.01 a | 0.66 ± 0.01 a |
| MC no. 2 | 1.03 ± 0.37 a | 0.75 ± 0.08 a | |
| MC no. 3 | 0.92 ± 0.15 ab | 0.79 ± 0.21 a | |
| Average of MC 1-3 | 0.91 ± 0.12 | 0.73 ± 0.07 | |
| Extracts | MICs, µg mL−1 | |||
|---|---|---|---|---|
| Escherichia coli ATCC 25922 | Staphylococcus aureus ATCC 25923 | MRSA ATCC 33591 | Pseudomonas aeruginosa ATCC 27853 | |
| Cell biomass | 250 | 500 | 2000 | 500 |
| Plant leaves | 4000 | 4000 | 4000 | 4000 |
| Extracts | Assay * | |
|---|---|---|
| DPPH EC50, mg mL−1 | TEAC EC50, mg mL−1 | |
| Cell biomass | 17.85 ± 0.44 a | 11.10 ± 0.98 a |
| Plant leaves | 10.69 ± 0.09 b | 8.81 ± 0.13 a |
| Positive control | 0.150 ± 0.002 | 0.036 ± 0.001 |
| Cultivation System | 250 mL Flasks on an Orbital Shaker | 20 L Bioreactors (Bubble-Type) |
|---|---|---|
| Mode of operation | Batch | Semi-continuous |
| Working volume | 35 mL medium | 15 L |
| Aeration device | No | Sparger * nh = 1 dh = 6.0 mm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titova, M.V.; Kochkin, D.V.; Sukhanova, E.S.; Gorshkova, E.N.; Tyurina, T.M.; Ivanov, I.M.; Lunkova, M.K.; Tsvetkova, E.V.; Orlova, A.; Popova, E.V.; et al. Suspension Cell Culture of Polyscias fruticosa (L.) Harms in Bubble-Type Bioreactors—Growth Characteristics, Triterpene Glycosides Accumulation and Biological Activity. Plants 2023, 12, 3641. https://doi.org/10.3390/plants12203641
Titova MV, Kochkin DV, Sukhanova ES, Gorshkova EN, Tyurina TM, Ivanov IM, Lunkova MK, Tsvetkova EV, Orlova A, Popova EV, et al. Suspension Cell Culture of Polyscias fruticosa (L.) Harms in Bubble-Type Bioreactors—Growth Characteristics, Triterpene Glycosides Accumulation and Biological Activity. Plants. 2023; 12(20):3641. https://doi.org/10.3390/plants12203641
Chicago/Turabian StyleTitova, Maria V., Dmitry V. Kochkin, Elena S. Sukhanova, Elena N. Gorshkova, Tatiana M. Tyurina, Igor M. Ivanov, Maria K. Lunkova, Elena V. Tsvetkova, Anastasia Orlova, Elena V. Popova, and et al. 2023. "Suspension Cell Culture of Polyscias fruticosa (L.) Harms in Bubble-Type Bioreactors—Growth Characteristics, Triterpene Glycosides Accumulation and Biological Activity" Plants 12, no. 20: 3641. https://doi.org/10.3390/plants12203641