
Exogenous Application of 24-Epibrassinolide Confers Saline Stress and Improves Photosynthetic Capacity, Antioxidant Defense, Mineral Uptake, and Yield in Maize

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design Treatments and Growth Conditions
2.1.1. Establishing the Nursery
2.1.2. Transplantation and Growth Conditions
2.1.3. Treatments and Experimental Design
2.2. Measurements
2.2.1. Evaluation of Plant Growth Parameters and Mineral Ions
2.2.2. Photosynthetic and Physiological Traits Assessment
2.2.3. Antioxidant Enzymatic Activity, Proline and Total Phenolic Contents
2.2.4. Yield and Yield Attributes
2.3. Statitical Analysis
3. Results
4. Discussion
4.1. Growth and Morphological Attributes
4.2. Photosynthetic Activity, Gaseous Exchange, and Physiological Performance
4.3. Antioxidants’ Activity
4.4. Mineral Uptake
4.5. Yield and Yield Attributes
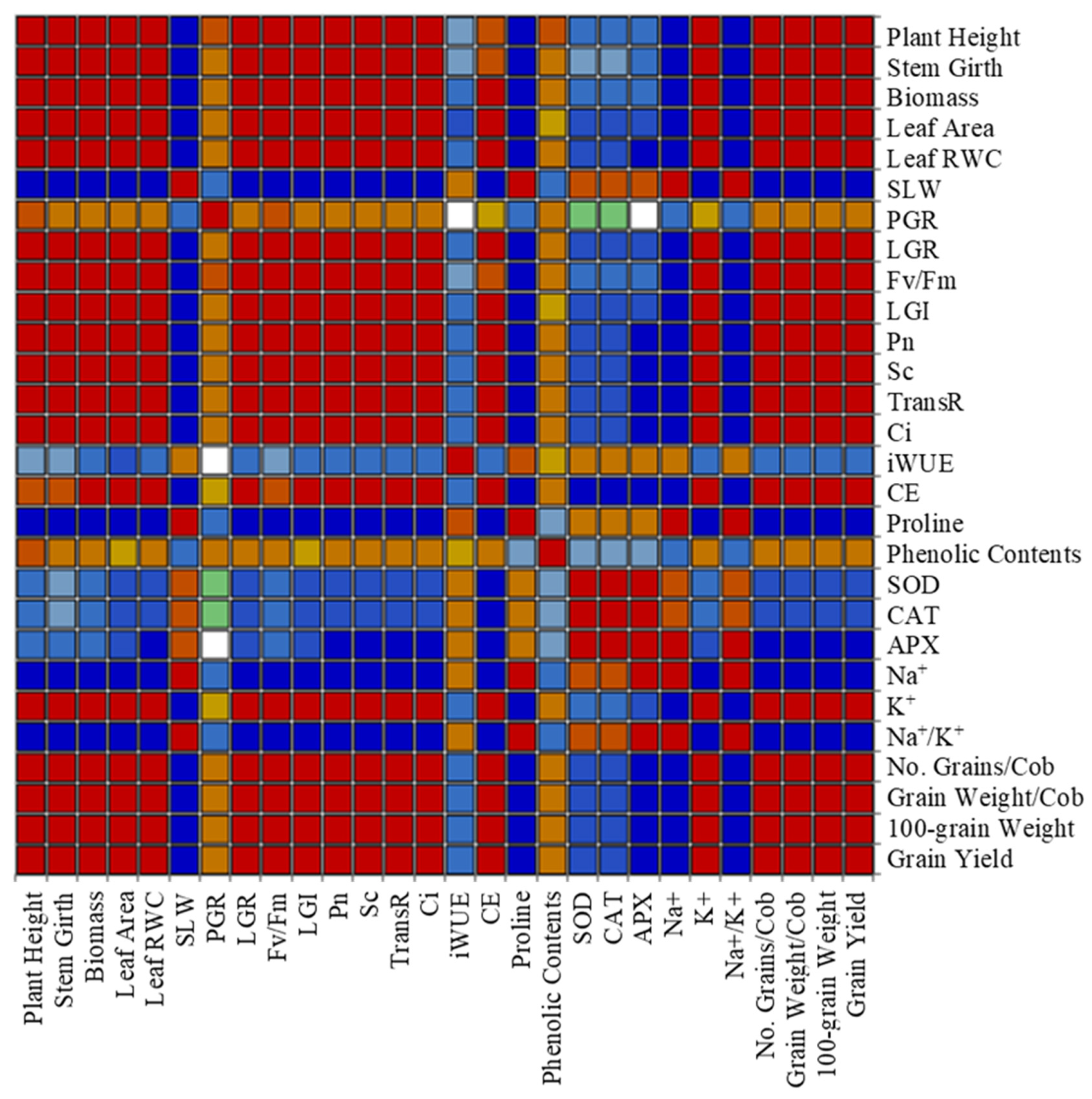
4.6. Correlations and PCA
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Haj-Amor, Z.; Araya, T.; Kim, D.G.; Bouri, S.; Lee, J.; Ghiloufi, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil salinity and its associated effects on soil microorganisms, greenhouse gas emissions, crop yield, biodiversity and desertification: A review. Sci. Total Environ. 2022, 843, 156946. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H. Activated Yeast extract enhances growth, anatomical structure, and productivity of Lupinus termis L. plants under actual salinity conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Dustgeer, Z.; Seleiman, M.F.; Khan, I.; Chattha, M.U.; Ali, E.F.; Alhammad, B.A.; Jalal, R.S.; Refay, Y.; Hassan, M.U. Glycine-betaine induced salinity tolerance in maize by regulating the physiological attributes, antioxidant defense system and ionic homeostasis. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12248. [Google Scholar] [CrossRef]
- Singh, A. Alternative management options for irrigation-induced salinization and waterlogging under different climatic conditions. Ecol. Indic. 2018, 90, 184–192. [Google Scholar] [CrossRef]
- Tian, F.; Hou, M.; Qiu, Y.; Zhang, T.; Yuan, Y. Salinity stress effects on transpiration and plant growth under different salinity soil levels based on thermal infrared remote (TIR) technique. Geoderma 2020, 357, 113961. [Google Scholar] [CrossRef]
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Grattan, S.R.; Rengasamy, P.; Ben-Gal, A.; Assouline, S.; Javaux, M.; Minhas, P.S.; et al. Critical knowledge gaps and research priorities in global soil salinity. Adv. Agron. 2021, 169, 1–191. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Nagpal, S.; Sharma, P. Potential of plant growth-promoting rhizobacteria-plant interactions in mitigating salt stress for sustainable agriculture: A review. Pedosphere 2022, 32, 223–245. [Google Scholar] [CrossRef]
- Lodeyro, A.F.; Carrillo, N. Salt stress in higher plants: Mechanisms of toxicity and defensive responses. In Salt Stress in Higher Plants: Mechanisms of Toxicity and Defensive Responses; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–33. [Google Scholar] [CrossRef]
- Alhammad, B.A.; Ahmad, A.; Seleiman, M.F.; Tola, E. Seed priming with nanoparticles and 24-epibrassinolide improved seed germination and enzymatic performance of Zea mays L. in salt-stressed soil. Plants 2023, 12, 690. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field crop responses and management strategies to mitigate soil salinity in modern agriculture: A review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Semida, W.M.; Rady, M.M.; Mohamed, G.F.; Hemida, K.A.; Alhammad, B.A.; Hassan, M.M.; Shami, A. Sequential application of antioxidants rectifies ion imbalance and strengthens antioxidant systems in salt-stressed cucumber. Plants 2020, 9, 1783. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R. Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Roychoudhury, A. Gene regulation at transcriptional and post-transcriptional levels to combat salt stress in plants. Physiol. Plant. 2021, 173, 1556–1572. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Huang, R. The phytohormonal regulation of Na+/K+ and reactive oxygen species homeostasis in rice salt response. Mol. Breed. 2020, 40, 47. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Wegner, L.H.; Stefano, G.; Shabala, L.; Rossi, M.; Mancuso, S.; Shabala, S. Sequential depolarization of root cortical and stelar cells induced by an acute salt shock–implications for Na+ and K+ transport into xylem vessels. Plant Cell Environ. 2011, 34, 859–869. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Gilliham, M.; Flowers, T.J.; Colmer, T.D. Tissue tolerance: An essential but elusive trait for salt-tolerant crops. Funct. Plant Biol. 2016, 43, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt stress induces physiochemical alterations in rice grain composition and quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Awais, A.; El-Kamil, T.; Thobayet, S.; Alshahrani, S.; Seleiman, M.F. Enhancement of morphological and physiological performance of Zea mays L. under saline stress using ZnO nanoparticles and 24-epibrassinolide seed priming. Agronomy 2023, 13, 771. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Ahmad, A.; Alshahrani, T.S. Integrative effects of zinc nanoparticle and PGRs to mitigate salt stress in maize. Agronomy 2023, 13, 1655. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Athar, H.U.R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Tanveer, M. Role of 24-Epibrassinolide in inducing thermo-tolerance in plants. J. Plant Growth Regul. 2019, 38, 945–955. [Google Scholar] [CrossRef]
- Siddiqui, H.; Ahmed, K.B.M.; Hayat, S. Comparative effect of 28-homobrassinolide and 24-epibrassinolide on the performance of different components influencing the photosynthetic machinery in Brassica juncea L. Plant Physiol. Biochem. 2018, 129, 198–212. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol. Biochem. 2019, 135, 295–303. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Che, Z.; Rehman, A.; Cheema, S.A.; Sharma, A.; Song, H.; ur Rehman, S.; Zhaorong, D. Role of 24-epibrassinolide (EBL) in mediating heavy metal and pesticide induced oxidative stress in plants: A review. Ecotoxicol. Environ. Saf. 2018, 147, 935–944. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Biju, S.; Bhardwaj, R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review. Plant Physiol. Biochem. 2018, 130, 69–79. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Anastassiadou, M.; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Marques, D.C.; et al. Peer review of the pesticide risk assessment of the active substance 24-epibrassinolide. EFSA J. 2020, 18, e06132. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in Boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Reynolds, T.W.; Waddington, S.R.; Anderson, C.L.; Chew, A.; True, Z.; Cullen, A. Environmental impacts and constraints associated with the production of major food crops in Sub-Saharan Africa and South Asia. Food Secur. 2015, 7, 795–822. [Google Scholar] [CrossRef]
- Erenstein, O.; Chamberlin, J.; Sonder, K. Estimating the global number and distribution of maize and wheat farms. Glob. Food Secur. 2021, 30, 100558. [Google Scholar] [CrossRef]
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Wang, J.; Hu, X. Research on corn production efficiency and influencing factors of typical farms: Based on data from 12 corn-producing countries from 2012 to 2019. PLoS ONE 2021, 16, e0254423. [Google Scholar] [CrossRef]
- Basit, F.; Chen, M.; Ahmed, T.; Shahid, M.; Noman, M.; Liu, J.; An, J.; Hashem, A.; Fahad Al-Arjani, A.B.; Alqarawi, A.A.; et al. Seed priming with brassinosteroids alleviates chromium stress in rice cultivars via improving ROS metabolism and antioxidant defense response at biochemical and molecular levels. Antioxidants 2021, 10, 1089. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B. The comprehensive system of leaf analysis and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 3, 1035–1059. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2017, 104, 1372–1378. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef]
- Talat, N. Alleviation of soil salinization and the management of saline soils, climate change, and soil interactions. In Climate Change and Soil Interactions; Elsevier: Amsterdam, The Netherlands, 2020; pp. 305–329. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Egamberdieva, D.; Alam, P.; Alyemeni, M.N.; Ashraf, M. Modification of osmolytes and antioxidant enzymes by 24-epibrassinolide in chickpea seedlings under mercury (Hg) toxicity. J. Plant Growth Regul. 2018, 37, 309–322. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Mansour, E.; Ali, M.M.; Yasin, M.A.; Abdul-Hamid, M.I.; Rady, M.M.; Ali, E.F. Exogenously used 24-epibrassinolide promotes drought tolerance in maize hybrids by improving plant and water productivity in an arid environment. Plants 2021, 10, 354. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, R.; Harlina, P.W.; Ewas, M.; Zhenyuan, P.; Nie, X.; Gallego, P.P.; Ullah Khan, S.; Nishawy, E.; Khan, A.H.; Jia, H. Foliar applied 24-epibrassinolide alleviates salt stress in rice (Oryza sativa L.) by suppression of ABA levels and upregulation of secondary metabolites. J. Plant Interact. 2021, 16, 533–549. [Google Scholar] [CrossRef]
- Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings. Forests 2018, 10, 4. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to salt stress in lettuce: Changes in chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Yun, P.; Xu, L.; Wang, S.S.; Shabala, L.; Shabala, S.; Zhang, W.Y. Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoot. Plant Growth Regul. 2018, 86, 323–331. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef]
- Babitha, K.C.; Vemanna, R.S.; Nataraja, K.N.; Udayakumar, M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress. PLoS ONE 2015, 10, e0137098. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef]
- Sharma, I.; Pati, P.K.; Bhardwaj, R. Effect of 24-epibrassinolide on oxidative stress markers induced by nickel-ion in Raphanus sativus L. Acta Physiol. Plant. 2011, 33, 1723–1735. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Agami, R.A. Alleviating the adverse effects of NaCl stress in maize seedlings by pretreating seeds with salicylic acid and 24-epibrassinolide. S. Afr. J. Bot. 2013, 88, 171–177. [Google Scholar] [CrossRef]
- Goda, H.; Shimada, Y.; Asami, T.; Fujioka, S.; Yoshida, S. Microarray analysis of brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2002, 130, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Shu, S.; Sun, J.; Guo, S.; Tezuka, T. Effects of 24-epibrassinolide on the photosynthetic characteristics, antioxidant system, and chloroplast ultrastructure in Cucumis sativus L. under Ca(NO3)2 stress. Photosynth. Res. 2012, 112, 205–214. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, W.; Hu, G.; Chen, W.; Zhuge, Y.; Wang, Z.; He, M.R. Role of exogenous 24-epibrassinolide in enhancing the salt tolerance of wheat seedlings. J. Soil Sci. Plant Nutr. 2017, 17, 554–569. [Google Scholar] [CrossRef]
- Azhar, N.; Su, N.; Shabala, L.; Shabala, S. Exogenously applied 24-epibrassinolide (EBL) ameliorates detrimental effects of salinity by reducing K+ efflux via depolarization-activated K+ channels. Plant Cell Physiol. 2017, 58, 802–810. [Google Scholar] [CrossRef]
- Das, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Understanding salinity responses and adopting ‘omics-based’ approaches to generate salinity tolerant cultivars of rice. Front. Plant Sci. 2015, 6, 712. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Shabala, S.; Cuin, T.A.; Pang, J.; Percey, W.; Chen, Z.; Conn, S.; Eing, C.; Wegner, L.H. Xylem ionic relations and salinity tolerance in barley. Plant J. 2010, 61, 839–853. [Google Scholar] [CrossRef]
- Shahid, M.; Naeem-Ullah, U.; Khan, W.; Saeed, D.S.; Razzaq, K. Application of nanotechnology for insect pests management: A review. J. Innov. Sci. 2021, 7, 28–39. [Google Scholar] [CrossRef]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogués, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Hayat, S.; Ahmad, A. 28-Homobrassinolide ameliorates the saline stress in chickpea (Cicer arietinum L.). Environ. Exp. Bot. 2007, 59, 217–223. [Google Scholar] [CrossRef]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy. The Principles and Practice of Numerical Classification; W H Freeman & Co.: New York, NY, USA, 1973. [Google Scholar]
- Saed-Moucheshi, A.; Pessarakli, M.; Heidari, B. Comparing relationships among yield and its related traits in mycorrhizal and nonmycorrhizal inoculated wheat cultivars under different water regimes using multivariate statistics. Int. J. Agron. 2013, 2013, 682781. [Google Scholar] [CrossRef]
- Afzal, M.; Alghamdi, S.S.; Migdadi, H.H.; El-Harty, E.; Al-Faifi, S.A. Agronomical and physiological responses of faba bean genotypes to salt stress. Agriculture 2022, 12, 235. [Google Scholar] [CrossRef]
- Otie, V.; Ibrahim, A.; Udo, I.; Kashiwagi, J.; Matsuura, A.; Shao, Y.; Itam, M.; An, P.; Eneji, A.E. Foliarly applied 24-Epibrassinolide modulates the electrical conductivity of the saturated rhizospheric soil extracts of soybean under salinity stress. Plants 2022, 11, 2330. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Significance | ||
|---|---|---|---|
| Salinity | EBL | Salinity × EBL | |
| Plant Height | ** | ** | NS |
| Stem Diameter | ** | ** | * |
| Biomass per Plant | ** | ** | NS |
| Leaf Area per Plant | ** | ** | ** |
| Leaf Relative Water Contents (RWC) | ** | ** | * |
| Specific Leaf Weight (SLW) | ** | ** | ** |
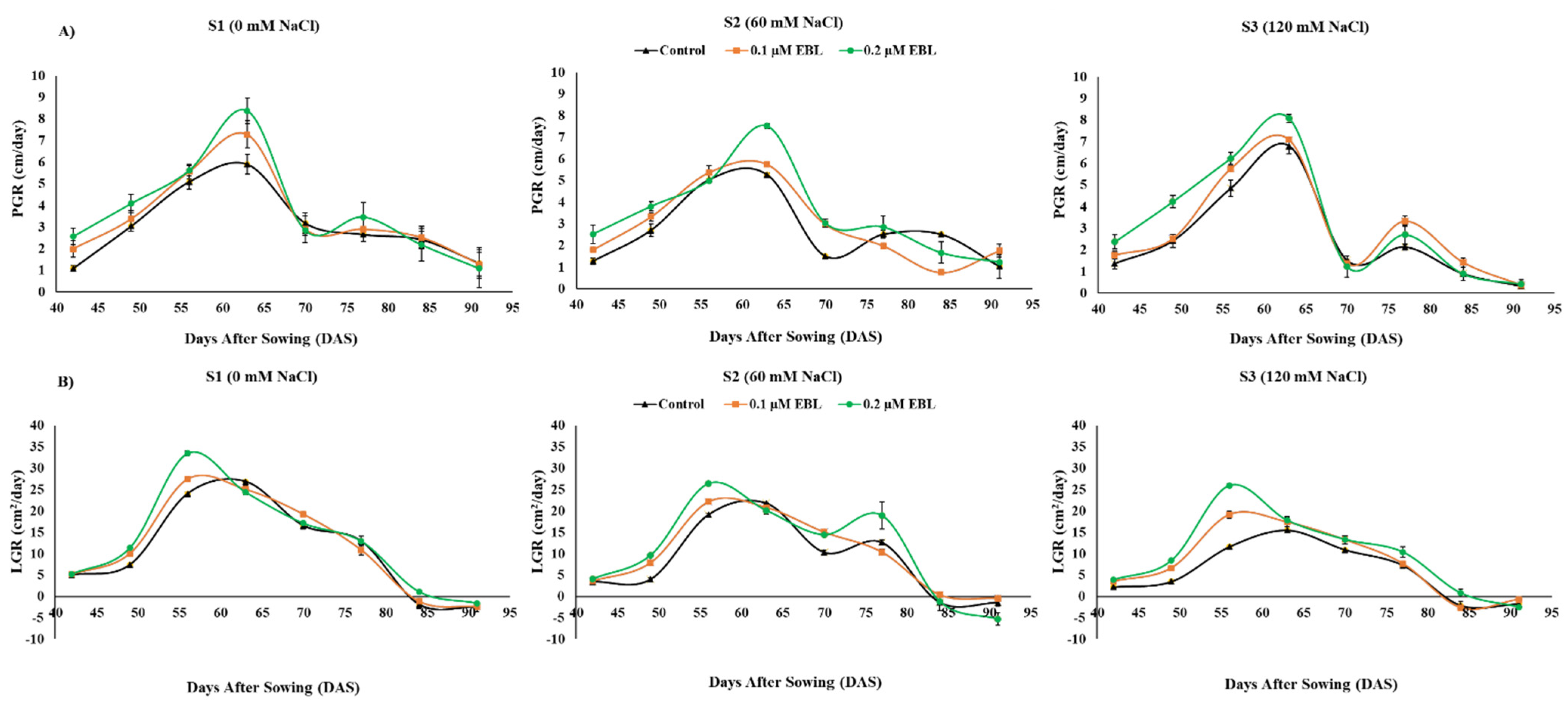
| Plant Growth Rate (PGR) | ** | ** | NS |
| Leaf Growth Rate (LGR) | ** | ** | ** |
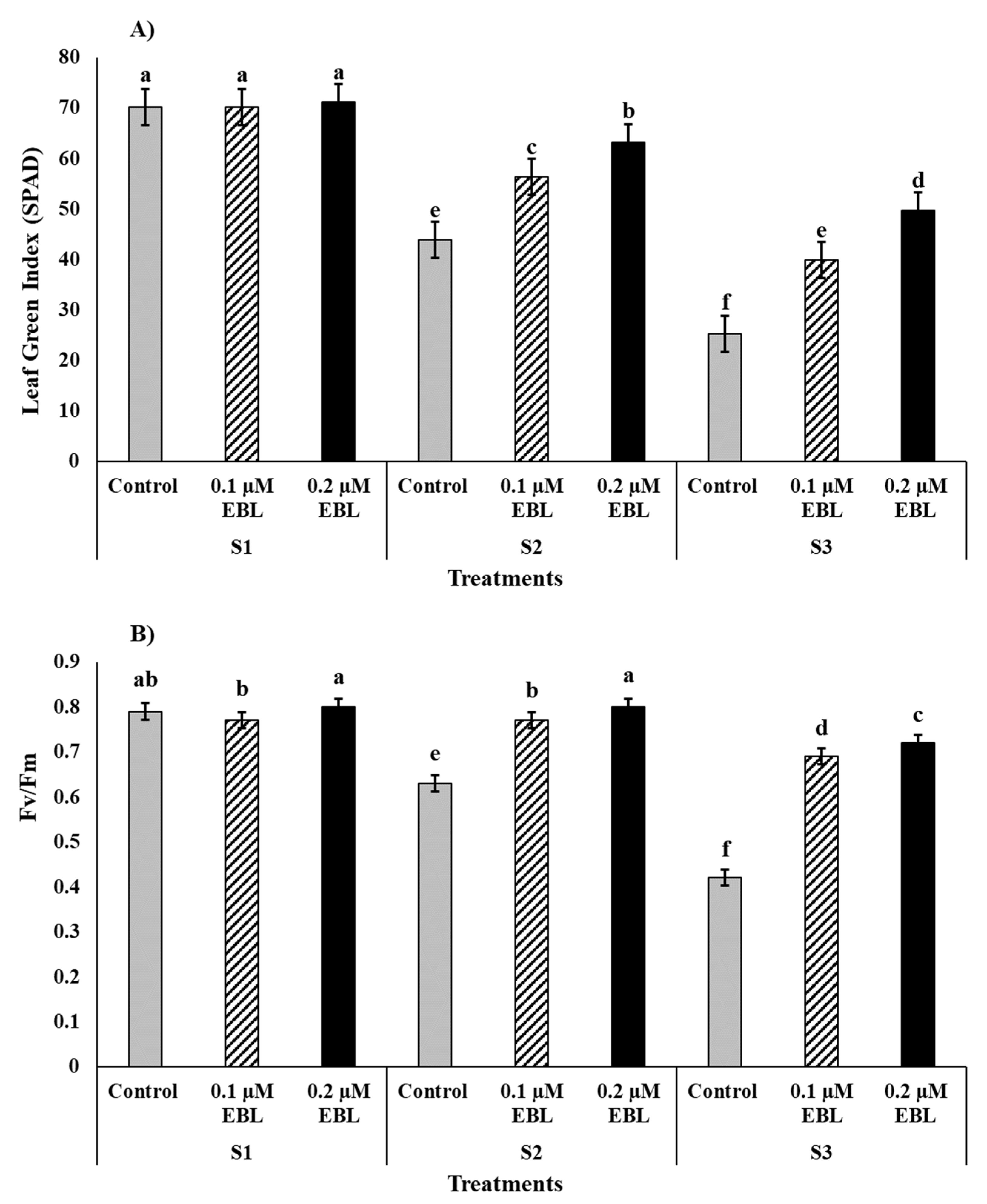
| Photochemical efficiency of PSII (Fv/Fm) | ** | ** | ** |
| Leaf Green Index (LGI) (SPAD-Reading) | ** | ** | ** |
| Rate of Photosynthesis (Pn) | ** | ** | ** |
| Stomatal Conductance (Sc) | ** | ** | * |
| Transpiration Rate (TransR) | ** | ** | ** |
| Intercellular CO2 Concentration (Ci) | ** | ** | NS |
| Intrinsic Water Use Efficiency (iWUE) | * | NS | NS |
| Carboxylation Efficiency (CE) | ** | ** | ** |
| Proline | ** | ** | ** |
| Total Phenolic Contents | ** | ** | * |
| Superoxide dismutase (SOD) | ** | ** | ** |
| Catalase (CAT) | ** | ** | ** |
| Ascorbate peroxidase (APX) | ** | ** | * |
| Na+ Concentration | ** | ** | ** |
| K+ Concentration | ** | ** | ** |
| Na+/K+ | ** | ** | ** |
| Number of Grains per Ear | ** | ** | NS |
| Grain Weight per Ear | ** | ** | NS |
| 100-grain Weight | ** | ** | ** |
| Grain Yield per Plant | ** | ** | ** |
| Treatments/Parameters | Plant Height (cm) | Stem Diameter (mm) | Biomass (g/plant) | Leaf Area (cm2/plant) | Leaf RWC (%) | SLW (g/m2) | |
|---|---|---|---|---|---|---|---|
| S1 | 228.56 A | 18.32 A | 114.94 A | 6903 A | 82.16 A | 50.51 C | |
| S2 | 213.67 B | 17.13 B | 99.60 B | 5490 B | 73.44 B | 53.23 B | |
| S3 | 187.11 C | 16.10 C | 86.98 C | 4596 C | 65.78 C | 58.38 A | |
| LSD0.05 | 4.77 | 0.36 | 2.65 | 27.08 | 2.21 | 1.04 | |
| EBL | |||||||
| Control | 191.78 C | 15.79 C | 92.96 C | 4794 C | 72.23 C | 56.57 A | |
| 0.1 µM EBL | 211.67 B | 17.42 B | 97.86 B | 5725 B | 73.58 B | 53.61 B | |
| 0.2 µM EBL | 225.89 A | 18.34 A | 110.71 A | 6469 A | 75.54 A | 51.94 C | |
| LSD0.05 | 3.88 | 0.39 | 3.56 | 51.51 | 1.04 | 0.81 | |
| Salinity × EBL | |||||||
| Salinity | EBL | ||||||
| S1 | Control | 209.67 | 16.37 de | 108.20 | 6304 c | 81.77 a | 51.77 de |
| 0.1 µM EBL | 230.67 | 18.93 b | 114.83 | 6889 b | 81.93 a | 50.63 e | |
| 0.2 µM EBL | 245.33 | 19.67 a | 121.80 | 7514 a | 82.77 a | 49.13 f | |
| S2 | Control | 197.33 | 15.93 e | 93.43 | 4726 g | 71.73 c | 55.27 c |
| 0.1 µM EBL | 212.67 | 17.03 c | 95.43 | 5587 f | 73.13 c | 52.97 d | |
| 0.2 µM EBL | 231.00 | 18.43 b | 109.93 | 6158 d | 75.47 b | 51.47 e | |
| S3 | Control | 168.33 | 15.07 f | 77.23 | 3353 h | 63.27 f | 62.67 a |
| 0.1 µM EBL | 191.67 | 16.30 de | 93.30 | 4700 g | 65.67 e | 57.23 b | |
| 0.2 µM EBL | 201.33 | 16.93 cd | 100.40 | 5734 e | 68.40 d | 55.23 c | |
| LSD0.05 | NS | 0.68 | NS | 89.22 | 1.80 | 1.40 | |
| Treatments/Parameters | Proline (mg/gFW) | Phenolic (mgGAE/gFM) | SOD (U/gFW) | CAT (U/gFW/min) | APX (umol/gFW/min) | |
|---|---|---|---|---|---|---|
| Salinity | ||||||
| S1 | 0.48 C | 45.07 B | 96.33 C | 30.11 C | 19.14 C | |
| S2 | 1.91 B | 56.08 A | 134.44 B | 47.23 B | 28.11 B | |
| S3 | 2.68 A | 31.30 C | 174.56 A | 62.11 A | 35.56 A | |
| LSD0.05 | 0.13 | 1.49 | 2.80 | 2.54 | 1.10 | |
| EBL | ||||||
| Control | 2.38 A | 41.06 C | 126.56 C | 42.56 C | 25.33 C | |
| 0.1 µM EBL | 1.60 B | 44.43 B | 133.89 B | 46.56 B | 27.11 B | |
| 0.2 µM EBL | 1.09 C | 47.00 A | 144.89 A | 50.22 A | 30.33 A | |
| LSD0.05 | 0.53 | 1.73 | 2.26 | 0.97 | 1.41 | |
| Salinity × EBL | ||||||
| Salinity | EBL | |||||
| S1 | Control | 0.49 f | 44.32 d | 93.33 h | 29.33 g | 18.67 e |
| 0.1 µM EBL | 0.49 f | 45.76 d | 95.67 h | 30.00 g | 18.33 e | |
| 0.2 µM EBL | 0.48 f | 45.14 d | 100.00 g | 31.00 g | 20.33 e | |
| S2 | Control | 2.83 b | 52.06 c | 126.33 f | 41.67 f | 25.67 d |
| 0.1 µM EBL | 1.86 d | 56.06 b | 134.33 e | 47.33 e | 27.67 d | |
| 0.2 µM EBL | 1.05 e | 60.12 a | 142.67 d | 52.33 d | 31.00 c | |
| S3 | Control | 3.83 a | 26.79 g | 160.00 c | 56.67 c | 31.67 c |
| 0.1 µM EBL | 2.45 c | 31.47 f | 171.67 b | 62.33 b | 35.33 b | |
| 0.2 µM EBL | 1.76 d | 35.63 e | 192.00 a | 67.33 a | 39.67 a | |
| LSD0.05 | 0.09 | 3.00 | 3.94 | 1.68 | 2.44 | |
| Treatments/Parameters | Number of Grains per Ear | Grain Weight per Ear (g) | 100-grain Weight (g) | Grain Yield (g/plant) | |
|---|---|---|---|---|---|
| Salinity | |||||
| S1 | 210.19 A | 58.27 A | 27.78 A | 175.99 A | |
| S2 | 161.02 B | 38.51 B | 23.98 B | 101.00 B | |
| S3 | 97.89 C | 18.23 C | 18.22 C | 48.10 C | |
| LSD0.05 | 4.78 | 1.67 | 0.64 | 6.98 | |
| EBL | |||||
| Control | 136.67 C | 32.02 C | 22.07 C | 88.16 C | |
| 0.1 µM EBL | 153.44 B | 37.29 B | 23.33 B | 107.67 B | |
| 0.2 µM EBL | 178.78 A | 45.70 A | 24.58 A | 129.24 A | |
| LSD0.05 | 5.81 | 1.89 | 0.27 | 4.14 | |
| Salinity × EBL | |||||
| Salinity | EBL | ||||
| S1 | Control | 190.00 | 51.10 | 27.20 b | 153.70 c |
| 0.1 µM EBL | 204.05 | 56.33 | 27.60 b | 170.30 b | |
| 0.2 µM EBL | 236.00 | 67.38 | 28.53 a | 203.97 a | |
| S2 | Control | 143.00 | 32.17 | 22.37 e | 93.70 f |
| 0.1 µM EBL | 163.03 | 38.43 | 24.20 d | 101.47 e | |
| 0.2 µM EBL | 177.00 | 44.93 | 25.37 c | 117.83 d | |
| S3 | Control | 77.00 | 12.80 | 16.63 h | 27.07 i |
| 0.1 µM EBL | 93.33 | 17.10 | 18.20 g | 51.23 h | |
| 0.2 µM EBL | 123.33 | 24.80 | 19.83 f | 66.00 g | |
| LSD0.05 | NS | NS | 0.47 | 7.17 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seleiman, M.F.; Ahmad, A.; Tola, E.; Alhammad, B.A.; Almutairi, K.F.; Madugundu, R.; Al-Gaadi, K.A. Exogenous Application of 24-Epibrassinolide Confers Saline Stress and Improves Photosynthetic Capacity, Antioxidant Defense, Mineral Uptake, and Yield in Maize. Plants 2023, 12, 3559. https://doi.org/10.3390/plants12203559
Seleiman MF, Ahmad A, Tola E, Alhammad BA, Almutairi KF, Madugundu R, Al-Gaadi KA. Exogenous Application of 24-Epibrassinolide Confers Saline Stress and Improves Photosynthetic Capacity, Antioxidant Defense, Mineral Uptake, and Yield in Maize. Plants. 2023; 12(20):3559. https://doi.org/10.3390/plants12203559
Chicago/Turabian StyleSeleiman, Mahmoud F., Awais Ahmad, ElKamil Tola, Bushra Ahmed Alhammad, Khalid F. Almutairi, Rangaswamy Madugundu, and Khalid A. Al-Gaadi. 2023. "Exogenous Application of 24-Epibrassinolide Confers Saline Stress and Improves Photosynthetic Capacity, Antioxidant Defense, Mineral Uptake, and Yield in Maize" Plants 12, no. 20: 3559. https://doi.org/10.3390/plants12203559