
Genome-Wide Analysis of Lipoxygenase (LOX) Genes in Angiosperms
 ,
,
Abstract
:1. Introduction
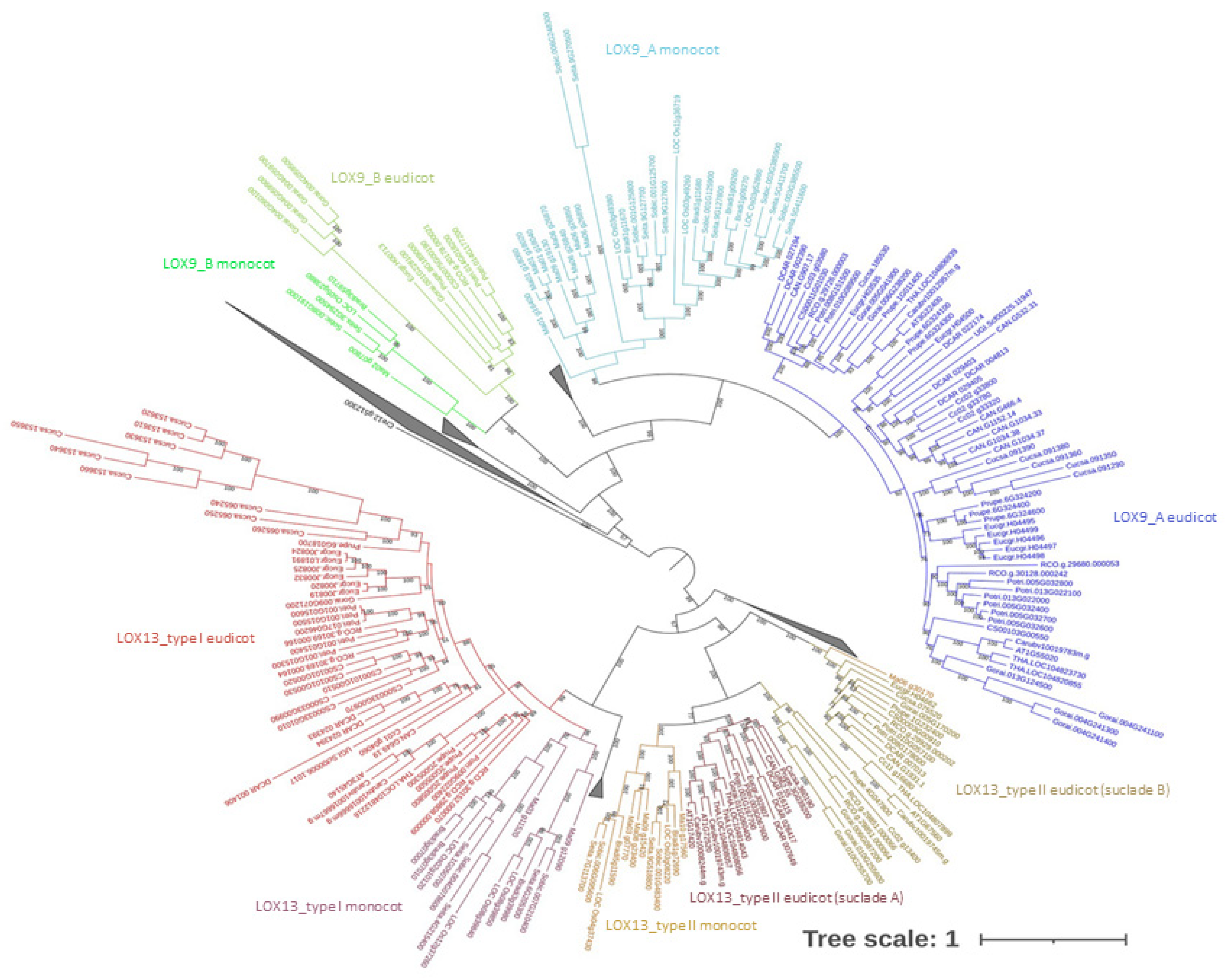
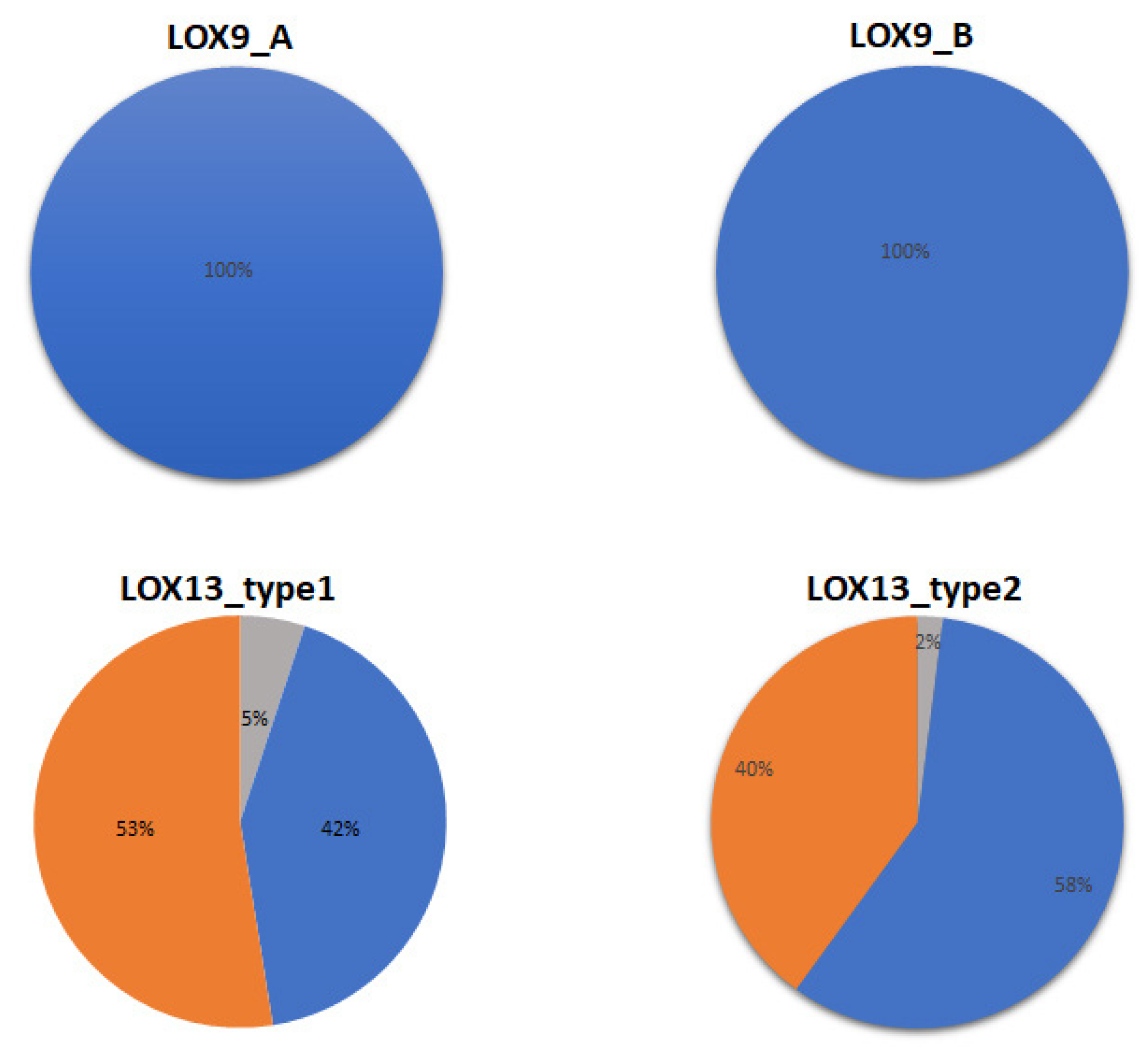
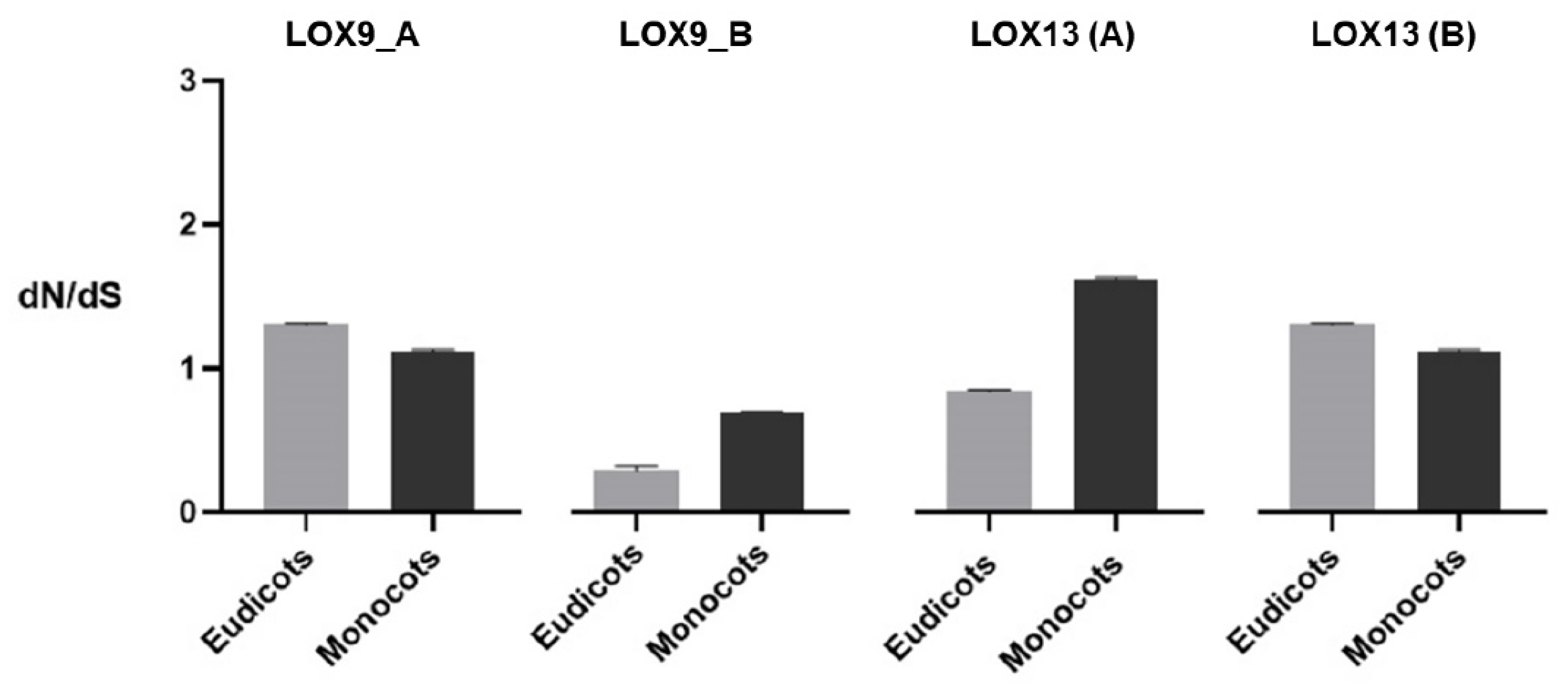
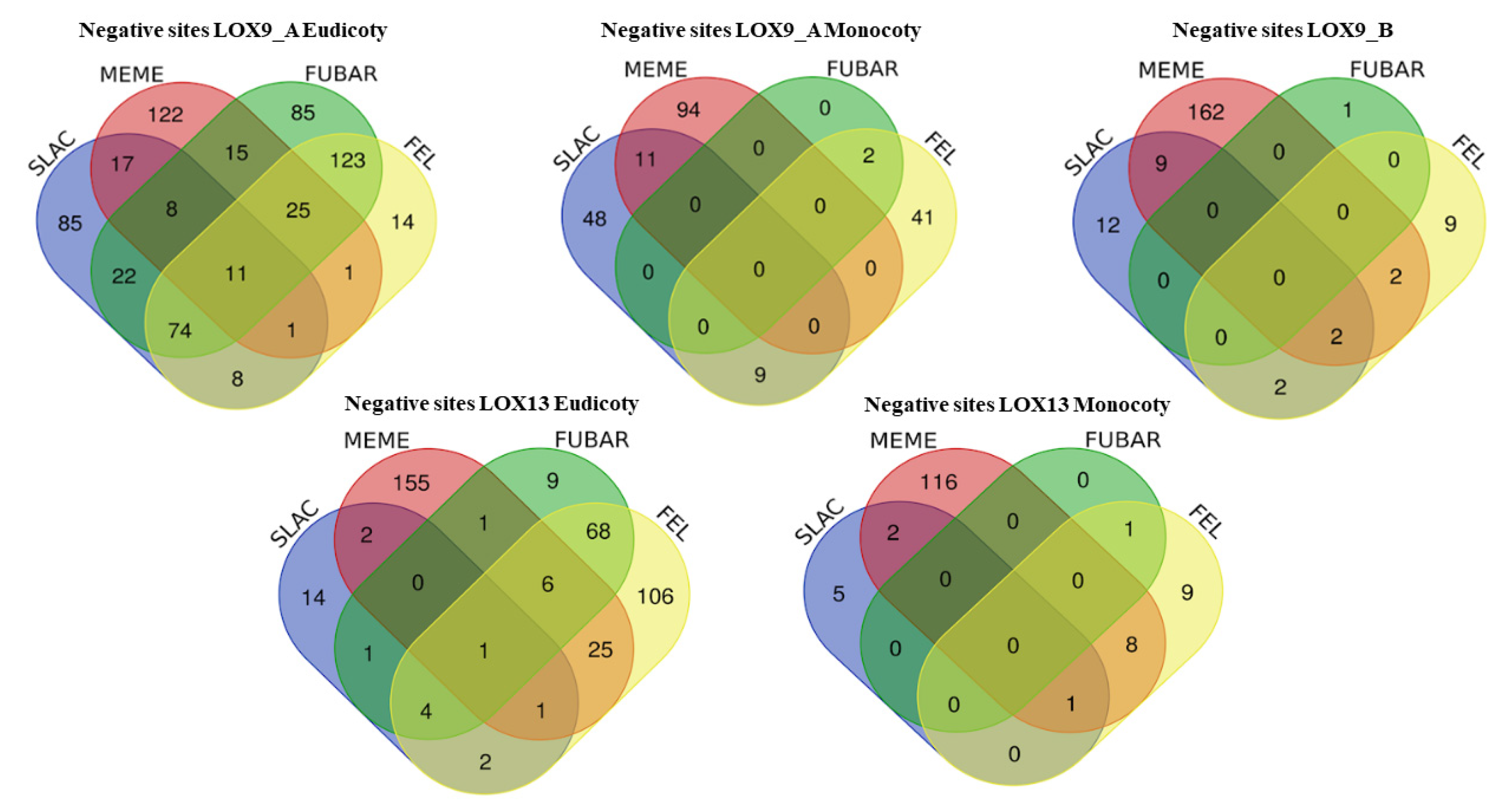
2. Results
3. Discussion
4. Materials and Methods
4.1. Identification and Annotation of LOX Family Genes
4.2. Multiple Sequence Alignment and Phylogenetic Analysis
4.3. Determination of Gene Structures
4.4. Selection Pressure and Evolutionary Analysis
4.5. Analysis of LOX Gene Expression Profiles in Angiosperms
4.6. Investigation of Motif Sequences and Cellular Localization of LOX Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Viswanath, K.K.; Varakumar, P.; Pamuru, R.R.; Basha, S.J.; Rao, A.D. Plant Lipoxygenases and their role in plant physiology. J. Plant Biol. 2020, 63, 83–95. [Google Scholar] [CrossRef]
- Guo, S.; Song, Z.; Ma, R.; Yang, Y.; Yu, M. Genome-wide identification and expression analysis of the lipoxygenase gene family during peach fruit ripening under different postharvest treatments. Acta Physiol. Plant 2017, 39, 111. [Google Scholar] [CrossRef]
- Zheng, K.; Wang, Z.; Pang, L.; Song, Z.; Zhao, H.; Wang, Y.; Wang, B.; Han, S. Systematic identification of methyl jasmonate-responsive long noncoding rnas and their nearby coding genes unveils their potential defence roles in tobacco BY-2 cells. Int. J. Mol. Sci. 2022, 23, 15568. [Google Scholar] [CrossRef]
- Cebrián, G.; Segura, M.; Martínez, J.; Iglesias-Moya, J.; Martínez, C.; Garrido, D.; Jamilena, M. Jasmonate-deficient mutant lox3a reveals crosstalk between JA and ET in the differential regulation of male and female flower opening and early fruit development in Cucurbita pepo. J. Exp. Bot. 2022, erac468. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhi, F.; Min, Y.; Ma, R.; Ge, A.; Wang, S.; Wang, J.; Liu, Z.; Guo, Y.; Chen, M. The MYB59 Transcription factor negatively regulates salicylic acid- and jasmonic acid-mediated leaf senescence. Plant Physiol. 2022, kiac589. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.; Oliveira, M.G.A.; Lanna, A.C.; Pires, C.V.; Piovesan, N.D.; José, I.C.; Bárbara, R.; Batista, E.G.B.; Moreira, M.A. Characterization of lipoxygenase pathway of soybean plants resistant and susceptible to diaphorte phaseolorum f.sp. meridionalis, pathogen responsible for stem canker. Rev. Bras. Fisiol. Vegl. 2001, 13, 316–328. [Google Scholar] [CrossRef]
- Shaban, M.; Ahmed, M.M.; Sun, H.; Ullah, A.; Zhu, L. Genome-wide identification of lipoxygenase gene family in cotton and functional characterization in response to abiotic stresses. BMC Genom. 2018, 19, 599. [Google Scholar] [CrossRef]
- Umate, P. Genome-wide analysis of lipoxygenase gene family in Arabidopsis and rice. Plant Signal Behav. 2011, 6, 335–338. [Google Scholar] [CrossRef] [Green Version]
- Vogt, J.; Schiller, D.; Ulrich, D.; Schwab, W.; Dunemann, F. Identification of lipoxygenase (LOX) genes putatively involved in fruit flavour formation in apple (Malus × domestica). Tree Genet. Genomes 2013, 9, 1493–1511. [Google Scholar] [CrossRef]
- Sarde, S.J.; Kumar, A.; Remme, R.N.; Dicke, M. Genome-wide identification, classification and expression of lipoxygenase gene family in pepper. Plant Mol. Biol. 2018, 98, 375–387. [Google Scholar] [CrossRef]
- Feng, B.; Dong, Z.; Xu, Z.; An, X.; Quin, H.; Wu, N.; Wang, D.; Wang, T. Molecular analysis of lipoxygenase (LOX) genes in common wheat and phylogenetic investigation of LOX proteins from model and crop plants. J. Cereal Sci. 2010, 52, 387–394. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, X.; Guo, L.; Xu, Q.; Zhao, S.; Li, F.; Yan, X.; Liu, S.; Wei, C. Characterization and alternative splicing profiles of the lipoxygenase. Gene Family in Tea Plant (Camellia sinensis). Plant Cell Physiol. 2018, 59, 1765–1781. [Google Scholar] [CrossRef] [Green Version]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Li, L.; Dunwell, J.M.; Qiao, X.; Liu, X.; Zhang, S. Characterization of the lipoxygenase (LOX) gene family in the Chinese white pear (Pyrusbretschneideri) and comparison with other members of the Rosaceae. BMC Genom. 2014, 15, 444. [Google Scholar] [CrossRef] [Green Version]
- Van Bel, M.; Diels, T.; Vancaester, E.; Kreft, L.; Botzki, A.; Van de Peer, Y.; Coppens, F.; Vandepoele, K. PLAZA 4.0: An integrative resource for functional, evolutionary and comparative plant genomics. Nucleic Acids Res 2018, 46, D1190–D1196. [Google Scholar] [CrossRef]
- Liu, S.Q.; Liu, X.H.; Jiang, L.W. Genome-wide identification, phylogeny and expression analysis of the lipoxygenase gene family in cucumber. Genet. Mol. Res. 2011, 10, 2613–2636. [Google Scholar] [CrossRef]
- Wang, J.; Hu, T.; Wang, W.; Hu, H.; Wei, Q.; Wei, X.; Bao, C. Bioinformatics analysis of the lipoxygenase gene family in radish (Raphanussativus) and functional characterization in response to abiotic and biotic stresses. Int. J. Mol. Sci. 2019, 20, 6095. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Wang, P.; Li, C.; Han, S.; Lopez-Baltazar, J.; Zhang, X.; Wang, X. Identification of lipoxygenase (LOX) genes from legumes and their responses in wild type and cultivated peanut upon Aspergillus flavus infection. Sci. Rep. 2016, 12, 35245. [Google Scholar] [CrossRef] [Green Version]
- Edger, P.P.; Hall, J.C.; Harkess, A.; Tang, M.; Coombs, J.; Mohammadin, S.; Schranz, M.E.; Xiong, Z.; Leebens-Mack, J.; Meyers, B.C.; et al. Brassicales phylogeny inferred from 72 plastid genes: A reanalysis of the phylogenetic localization of two paleopolyploid events and origin of novel chemical defenses. Am. J. Bot. 2018, 105, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anterola, A.; Göbel, C.; Hornung, E.; Sellhorn, G.; Feussner, I.; Grimes, H. Physcomitrella patens has lipoxygenases for both eicosanoid and octadecanoid pathways. Phytochemistry 2009, 70, 40–52. [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Palmieri-Thiers, C.; Canaan, S.; Brunini, V.; Lorenzi, V.; Tomi, F.; Desseyn, J.L.; Garscha, U.; Oliw, E.H.; Berti, L.; Maury, J. A lipoxygenase with dual positional specificity is expressed in olives (Olea europaea L.) during ripening. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1791, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Shen, W.; Liu, L.; Jiang, L.; Liu, Y.; Su, N.; Wan, J. A novel lipoxygenase gene from developing rice seeds confers dual position specificity and responds to wounding and insect attack. Plant Mol. Biol. 2008, 66, 401–414. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Bio. Evo. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Dong, Q.; Magwanga, R.O.; Cai, X.; Lu, P.; Nyangasi Kirungu, J.; Zhou, Z.; Wang, X.; Wang, X.; Xu, Y.; Hou, Y.; et al. RNA-Sequencing, Physiological and RNAi Analyses Provide Insights into the Response Mechanism of the ABC-Mediated Resistance to Verticilliumdahliae Infection in Cotton. Genes 2019, 10, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ksouri, N.; Jiménez, S.; Wells, C.E.; Contreras-Moreira, B.; Gogorcena, Y. Transcriptional Responses in Root and Leaf of Prunus persica under Drought Stress Using RNA Sequencing. Front. Plant Sci. 2016, 7, 1715. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Xue, J.; Wang, L.; Huang, J.; Fu, C.; Yan, X. Transcriptomic analysis for different sex type of ricinus communis L. during development from apical buds to inflorescences by digital gene expression profiling. Front. Plant Sci. 2016, 12, 1208. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Contreras, I.K.; Zamora-Hernández, T.; Huerta-Heredia, A.A.; Capataz-Tafur, J.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Transcriptomic analysis of submergence-tolerant and sensitive Brachypodium distachyon ecotypes reveals oxidative stress as a major tolerance factor. Sci. Rep. 2016, 6, 27686. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.S.; Goodrich, L.V.; Brown, P.J.; James, B.T.; Moose, S.P.; Lambert, K.N.; Riechers, D.E. Transcriptome profiling and genome-wide association studies reveal gsts and other defense genes involved in multiple signaling pathways induced by herbicide safener in grain sorghum. Front. Plant Sci. 2019, 10, 192. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res 2011, 39, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res 2016, 44, 242–245. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.; Shank, S.D.; Spielman, S.J.; Li, M.; Muse, S.V.; Pond, S.L.K. Datamonkey 2.0: A Modern Web Application for Characterizing Selective and Other Evolutionary Processes. Mol. Biol. Evol. 2018, 35, 773–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Chen, Y.; Lu, C.; Hwang, J. Prediction of protein subcellular localization. Proteins: Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; Heijne von, G.; Elofsson, A.; Henrik, N. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
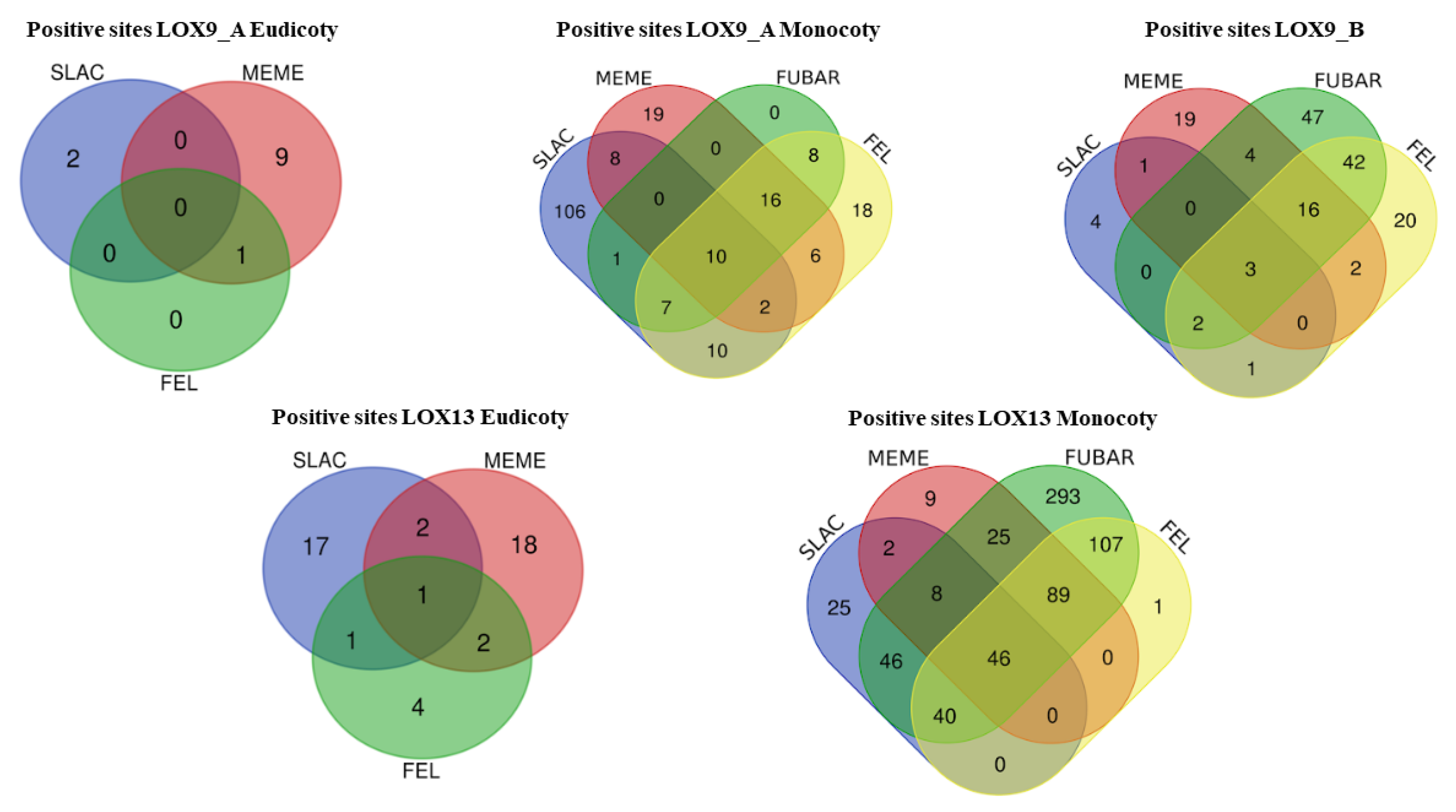{kind=link}
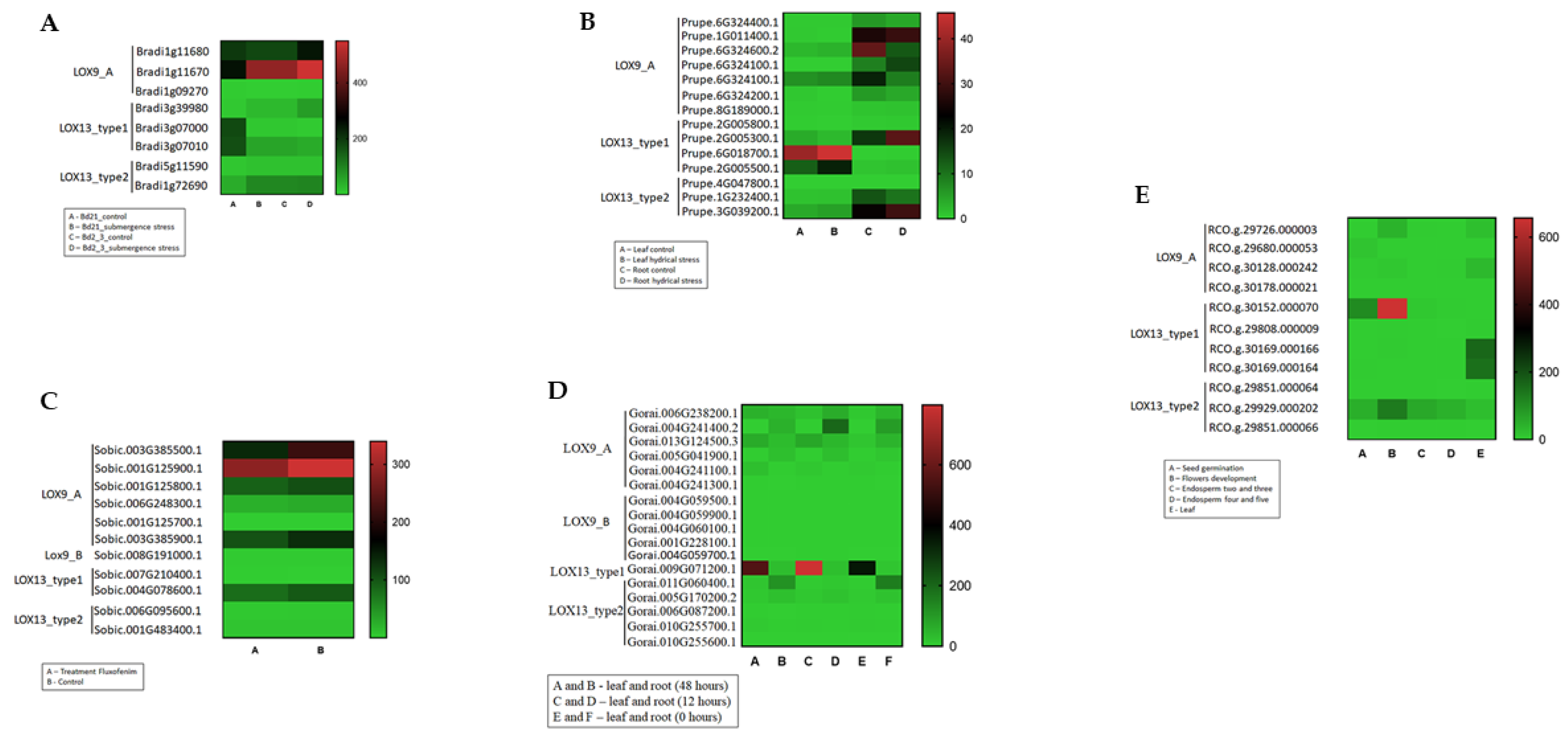{kind=link}
| Species | LOX9-A | LOX9-B | LOX13 | Total |
|---|---|---|---|---|
| Chlamydomonas reinhardtii | . | 1 | 1 | |
| Marchantia polymorpha | 16 | 16 | ||
| Picea abies | 3 | . | 5 | 8 |
| Amborella trichopoda | . | 1 | 3 | 4 |
| Musa acuminata | 9 | 1 | 7 | 17 |
| Setaria italica | 6 | 1 | 5 | 12 |
| Sorghum bicolor | 6 | 1 | 4 | 11 |
| Oryza sativa | 4 | 1 | 6 | 11 |
| Brachypodium distachyon | 4 | 1 | 5 | 10 |
| Daucus carota | 6 | 7 | 13 | |
| Coffea canephora | 4 | 3 | 7 | |
| Capsicum annuum | 7 | 3 | 10 | |
| Utricularia gibba | 1 | 1 | 2 | |
| Cucumis sativus | 6 | 11 | 17 | |
| Prunus persica | 6 | 1 | 7 | 14 |
| Ricinus communis | 3 | 1 | 7 | 11 |
| Populus trichocarpa | 8 | 2 | 10 | 20 |
| Eucalyptus grandis | 7 | 1 | 8 | 16 |
| Gossypium raimondii | 6 | 5 | 6 | 17 |
| Citrus sinensis | 2 | 1 | 7 | 10 |
| Tarenaya hassleriana | 3 | 5 | 8 | |
| Capsella rubella | 2 | 5 | 7 | |
| Arabidopsis thaliana | 2 | 4 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camargo, P.O.; Calzado, N.F.; Budzinski, I.G.F.; Domingues, D.S. Genome-Wide Analysis of Lipoxygenase (LOX) Genes in Angiosperms. Plants 2023, 12, 398. https://doi.org/10.3390/plants12020398
Camargo PO, Calzado NF, Budzinski IGF, Domingues DS. Genome-Wide Analysis of Lipoxygenase (LOX) Genes in Angiosperms. Plants. 2023; 12(2):398. https://doi.org/10.3390/plants12020398
Chicago/Turabian StyleCamargo, Paula Oliveira, Natália Fermino Calzado, Ilara Gabriela Frasson Budzinski, and Douglas Silva Domingues. 2023. "Genome-Wide Analysis of Lipoxygenase (LOX) Genes in Angiosperms" Plants 12, no. 2: 398. https://doi.org/10.3390/plants12020398