

Unraveling Large and Polyploidy Genome of the Crucifer Orychophragmus violaceus in China, a Potential Oil Crop
Abstract
:1. Introduction
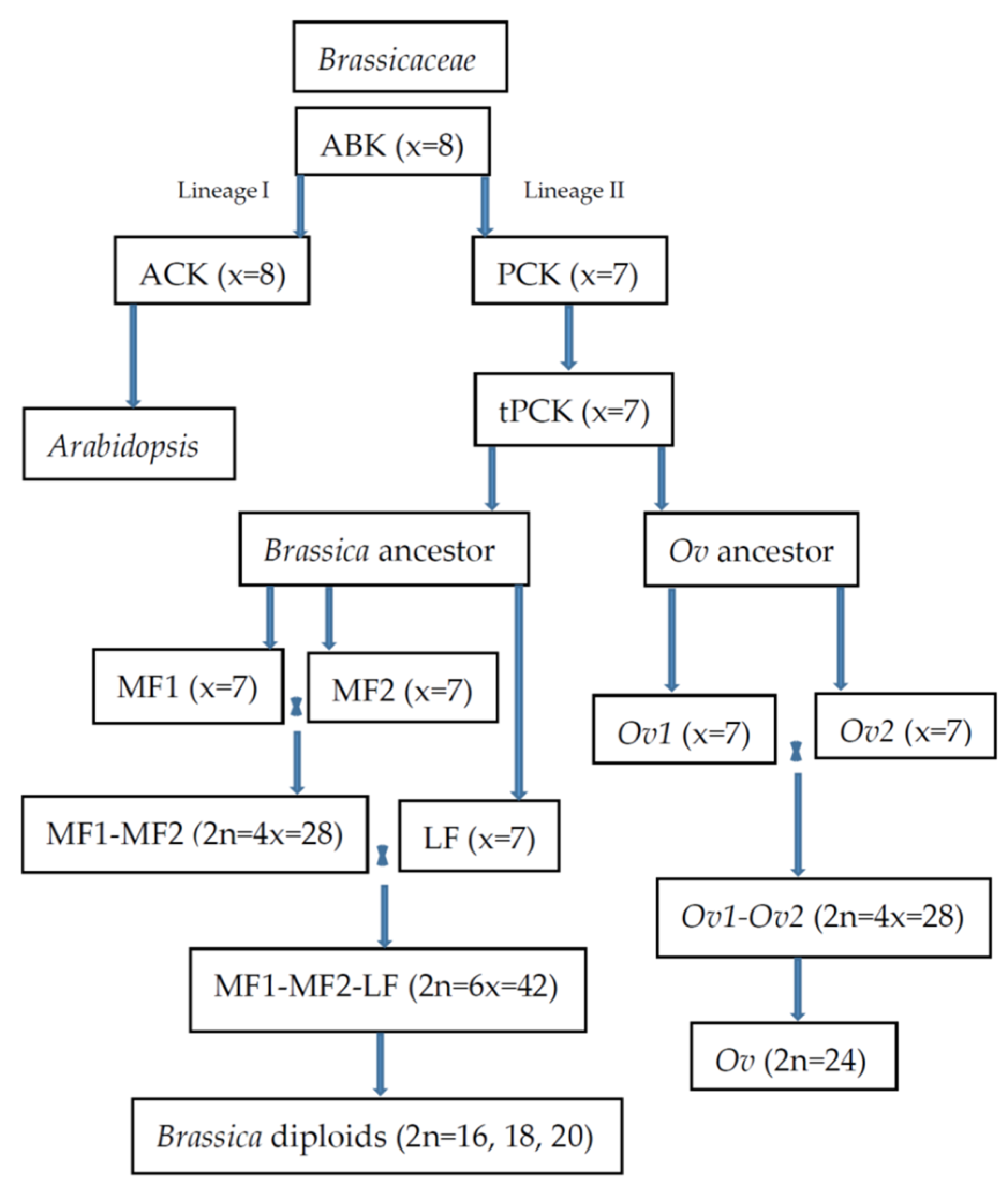
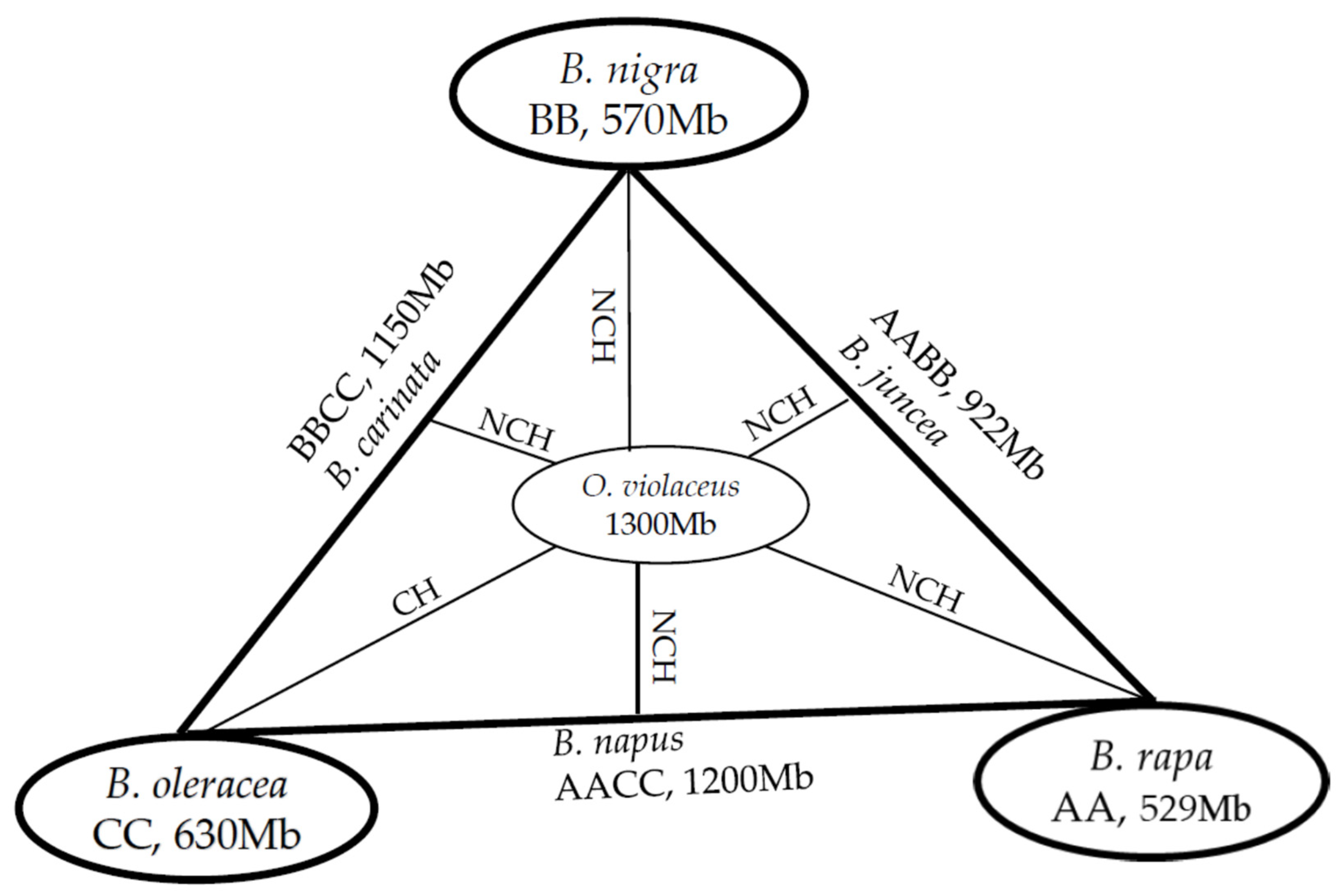
2. Phylogenetics
3. Large and Polyploidy Genome
4. Maintenance of Chromosome Features in Wide Hybrids
5. Genome Dissection via Development of Alien Additional Lines
6. A Potential Oil Crop
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, T.Y.; Guan, K.J.; Guo, R.L. Flora of China; Science Press: Beijing, China, 1987; Volume 33, pp. 40–43. [Google Scholar]
- Zhou, L.R.; Liu, Z.B.; Wu, J.; Wang, J.M.; Yang, Y.; Li, X.F. Karyotype variation and evolution in populations of the Chinese endemic Orychophragmus violaceus complex (Brassicaceae). Nord. J. Bot. 2008, 26, 375–383. [Google Scholar] [CrossRef]
- Zhou, L.R.; Yu, Y.; Song, R.X.; He, X.J.; Jiang, Y.; Li, X.F.; Yang, Y. Phylogenetic relationships within the Orychophragmus violaceus complex (Brassicaceae) endemic to China. Acta Bot. Yunnanica 2009, 31, 127–137. [Google Scholar] [CrossRef]
- Hu, H.; Al-Shehbaz, I.A.; Sun, Y.S.; Hao, G.Q.; Wang, Q.; Liu, J.Q. Species delimitation in Orychophragmus (Brassicaceae) based on chloroplast and nuclear DNA barcodes. Taxon 2015, 64, 714–726. [Google Scholar] [CrossRef]
- Tan, Z.M.; Xu, J.M.; Zhao, B.X.; Zhang, X.L. New taxa of Orychophragmus (Cruciferae) from China. Acta Phytotaxon. Sin. 1998, 36, 544–548. [Google Scholar]
- Luo, P.; Lan, Z.Q.; Huang, J.; Li, Z.Y. Study on valuable plant resource Orychophragmus violaceus (L.) O. E. Schulz. J. Nat. Resour. 1991, 6, 206–210. [Google Scholar]
- Huang, F.; Chen, P.; Tang, X.Y.; Zhong, T.; Yang, T.H.; Nwafor, C.C.; Yang, C.; Ge, X.H.; An, H.; Li, Z.Y.; et al. Genome assembly of the Brassicaceae diploid Orychophragmus violaceus reveals complex whole genome duplication and evolution of dihydroxy fatty acid metabolism. Plant Commu. 2022, 3, 100432. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, Y.Q.; Zhang, X.; Zhang, L.K.; Fu, Y.; Guo, Z.W.; Chen, S.M.; Wu, J.; Schnable, J.C.; Yi, K.K.; et al. The genome of Orychophragmus violaceus provides genomic insights into the evolution of Brassicaceae polyploidization and its distinct traits. Plant Commu. 2022, 3, 100431. [Google Scholar] [CrossRef]
- Couvreur, L.P.; Franzke, A.; Al-Shehbaz, I.A.; Bakker, F.T.; Koch, M.A.; Mummenhoff, K. Molecular phylogenetics, temporal diversification, and principles of evolution in the mustard family (Brassicaceae). Mol. Biol. Evol. 2010, 27, 55–71. [Google Scholar] [CrossRef]
- Yang, J.H.; Liu, D.Y.; Wang, X.W.; Ji, C.M.; Cheng, F.; Liu, B.N.; Hu, Z.Y.; Chen, S.; Pental, D.; Ju, Y.H.; et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeologous gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef]
- Song, J.M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.X.; Wang, B.; Lu, S.P.; Zhou, R.; et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef]
- Song, X.M.; Wei, Y.P.; Xiao, D.; Gong, K.; Sun, P.C. Brassica carinata genome characterization clarifies U’s triangle model of evolution and polyploidy in Brassica. Plant Physiol. 2021, 186, 388–406. [Google Scholar] [CrossRef] [PubMed]
- Belser, C.; Istace, B.; Denis, E.; Dubarry, M.; Baurens, F.; Falentin, C.; Genete, M.; Berrabah, W.; Chèvre, A.M.; Delourme, R.; et al. Chromosome-scale assemblies of plant genomes using nanopore long reads and optical maps. Nat. Plants 2018, 4, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Perumal, S.; Koh, C.S.; Jin, L.; Buchwaldt, M.; Higgins, E.E.; Zheng, C.F.; Sankoff, D.; Robinson, S.J.; Kagale, S.; Navabi1, Z.; et al. High contiguity long read assembly of Brassica nigra allows localization of active centromeres and provides insights into the ancestral Brassica genome. Nat. Plants 2020, 6, 929–941. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Cao, X.D.; Liu, D.X.; Liu, J.; Zhang, Z.S.; Jia, Y.J. A study on the karyotype of some Chinese variants of Zhuge Cai, Orychophragmus violaceus. Acta Agron. Sin. 1994, 20, 595–600. [Google Scholar]
- Hua, Y.W.; Liu, M.; Li, Z.Y. Parental genome separation and elimination of cells and chromosomes revealed by GISH and AFLP analysis in intergeneric hybrids between Brassica carinata and Orychophragmus violaceus. Ann. Bot. 2006, 97, 993–998. [Google Scholar] [CrossRef]
- Li, Z.Y.; Cartagena, J.; Kukui, K. Simultaneous detection of 45s and 5s rDNA genes in Orychophragmus violaceus by double fluorescence in situ hybridization. Cytologia 2005, 70, 459–466. [Google Scholar] [CrossRef]
- Zhao, Z.G.; Hu, T.T.; Ge, X.; Du, X.Z.; Ding, L.; Li, Z.Y. Production and characterization of intergeneric somatic hybrids between Brassica nausa and Orychophragms violaceus and their backcrossing progenies. Plant Cell Rep. 2008, 27, 1611–1621. [Google Scholar] [CrossRef]
- Ding, L.; Zhao, Z.G.; Ge, X.H.; Li, Z.Y. Intergeneric addition and substitution of Brassica napus with different chromosomes from Orychophragmus violaceus: Phenotype and cytology. Sci. Hortic. 2013, 164, 303–309. [Google Scholar] [CrossRef]
- Li, Z.Y.; Liu, H.L. A study on meiotic pairings in Orychophragmus violaceus. J. Huazhong Agri. Univ. (Nat. Sci. Edit.) 1995, 14, 435–439. [Google Scholar]
- Li, Z.Y.; Liu, H.L.; Luo, P. Production and cytogenetics of intergeneric hybrids between Brassica napus and Orychophragmus violaceus. Theor. Appl. Genet. 1995, 91, 131–136. [Google Scholar] [CrossRef]
- Yin, J.M.; Zhong, R.Q.; Lin, N.; Tang, Z.L.; Li, J.N. Microspore culture and observations on meiotic chromosome pairing of the haploid Orychophragmus violaceus. Acta Agron. Sin. 2020, 46, 194–203. [Google Scholar]
- Lysak, M.A.; Cheung, K.; Kitschke, M.; Bureš, P. Ancestral chromosomal blocks are triplicated in Brassiceae species with varying chromosome number and genome size. Plant Physiol. 2007, 145, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Wu, J.G.; Liu, Y.; Liu, H.L.; Heneen, W.K. Production and cytogenetics of intergeneric hybrids between Brassica juncea × Orychophragmus violaceus and B. carinata × O. violaceus. Theor. Appl. Genet. 1998, 96, 251–265. [Google Scholar] [CrossRef]
- Li, Z.Y.; Heneen, W.K. Production and cytogenetics of intergeneric hybrids between the three cultivated Brassica diploids and Orychophragmus violaceus. Theor. Appl. Genet. 1999, 99, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.M.; Luo, P.; Lan, Z.Q.; Huang, B.Q. Production of intergeneric hybrids from Brassica alboglabra × Orychophragmus violaceus. Acta Horticul. Sin. 1998, 25, 297–299. [Google Scholar]
- Li, Z.Y.; Liu, Y. Cytogenetics of intergeneric hybrids between Brassica species and Orychophragmus violaceus. Prog. Nat. Sci. 2002, 12, 8–12. [Google Scholar]
- Li, Z.Y.; Ge, X.G. Unique chromosome behavior and genetic control in Brassica × Orychophragmus wide hybrids: A review. Plant Cell Rep. 2007, 26, 701–710. [Google Scholar] [CrossRef]
- Fukui, K.; Nakayama, S.; Ohmido, N.; Yoshiaki, H.; Yamabe, M. Quantitative karyotyping of three diploid Brassica species by imaging methods and localization of 45S rDNA loci on the identified chromosomes. Theor. Appl. Genet. 1998, 96, 325–330. [Google Scholar] [CrossRef]
- Heneen, W.K.; Geleta, M.; Brismar, K.; Xiong, Z.Y.; Pires, J.C.; Hasterok, R.; Stoute, A.I.; Scott, R.J.; King, G.J.; Kurup, S. Seed colour loci, homoeology and linkage groups of the C genome chromosomes revealed in Brassica rapa-B. oleracea monosomic alien addition lines. Ann. Bot. 2012, 109, 1227–1242. [Google Scholar] [CrossRef]
- Heneen, W.K. Meiotic Karyotypes of the B Genomes of Brassica nigra and B. carinata. Hereditas 1999, 130, 131–135. [Google Scholar]
- Heneen, W.K.; Chen, B.Y.; Cheng, B.F.; Jonsson, A.; Simonsen, V.; Jørgensen, R.B.; Davik, J. Characterization of the A and C genomes of Brassica campestris and B. alboglabra. Hereditas 1995, 123, 251–267. [Google Scholar] [CrossRef]
- Fu, W.Q.; Chen, D.Z.; Pan, Q.; Li, F.F.; Zhao, Z.G.; Ge, X.H.; Li, Z.Y. Production of red-flowered oilseed rape via the ectopic expression of Orychophragmus violaceus OvPAP2. Plant Biotechnol. J. 2018, 16, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Wang, T.; Fu, W.; Harun, A.; Ge, X.; Li, Z.Y. Dosage-dependent gynoecium development and gene expression in Brassica napus-Orychophragmus violaceus addition lines. Plants 2021, 10, 1766. [Google Scholar] [CrossRef]
- Luo, P.; Lan, Z.Q.; Li, Z.Y. Orychophragmus violaceus, a potential edible-oil crop. Plant Breed. 1994, 113, 83–85. [Google Scholar] [CrossRef]
- Ma, N.; Li, Z.Y. Development of Novel Brassica napus lines with canola quality and higher levels of oleic and linoleic acids derived from intergeneric hybrids between B. napus and Orychophragmus violaceus. Euphytica 2007, 157, 231–238. [Google Scholar] [CrossRef]
- Li, X.J.; Teitgen, A.M.; Shirani, A.; Ling, J.; Busta, L.; Cahoon, R.E.; Zhang, W.; Li, Z.Y.; Chapman, K.D.; Berman, D.; et al. Discontinuous fatty acid elongation yields hydroxylated seed oil with improved function. Nat. Plants 2018, 4, 711–720. [Google Scholar] [CrossRef]
- Zhu, N.L.; Wu, H.F.; Xu, Z.Q.; Liu, C.Q.; Tian, Y.; Hu, M.G.; Sun, Z.H.; Li, P.F.; Ma, G.X.; Xu, X.D. New alkaloids with unusual spermidine moieties from the seeds of Orychophragmus violaceus and their cytoprotective properties. RSC Adv. 2017, 7, 41495–41498. [Google Scholar] [CrossRef]
- Huo, X.W.; Liu, C.Q.; Gao, L.; Xu, X.D.; Zhu, N.L.; Cao, L. Hepatoprotective effect of aqueous extract from the seeds of Orychophragmus violaceus against liver injury in mice and HepG2 cells. Intl. J. Mol. Sci. 2017, 18, 1197–1212. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Liu, C.Q.; Wang, S.J. Studies on the Karst Landform Adaptations of Orychophragmus violaceus; Guizhou Press of Science and Technology: Guiyang, China, 2004; pp. 1–154. [Google Scholar]
- Xu, X.X.; Xu, Z.H. Organogenesis in tissue culture of Orychophragmus violaceus. Acta Biol. Exp. Sin. 1987, 20, 503–507. [Google Scholar]
- Xu, X.X.; Xu, Z.H. Plant regeneration from mesophyll protoplasts of Orychophragmus violaceus. Acta Phytophysiol. Sin. 1988, 14, 170–174. [Google Scholar]
- Zhang, X.M.; Luo, P. Direct somatic embryogenesis from the explants of Orychophragmus violaceus. J. Sichuan Univ. Nat. Sci. Edit. 1995, 32, 587–593. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | 2n | Locations | Altitudes (m) |
|---|---|---|---|
| O. diffusus | 20 | Shanghai, East China | 50 |
| O. hupehensis | 22 | Hubei Province, Central China | 400~1000 |
| O. violaceus | 24 | Over China | Wide ranges |
| O. taibaiensis | 48 | Taibai Mountain, Northwest China | 1200 |
| AALs | Traits |
|---|---|
| MAAL1/DAAL1 | Leaf serration of O. violaceus. |
| MAAL2 | Biased to B. napus but with small oval leaves and flowers. |
| MAAL3 | Flower petals with white and shrunken margins. |
| MAAL4/DAAL4 | MAAL4 with yellow flowers, DAAL4 with red flowers of O. violaceus. Active rDNA loci. |
| MAAL5 | Basal branching of O. violaceus. Active rDNA loci. |
| MAAL6 | Early flowering and short plants. Active rDNA loci. |
| MAAL7 | Female sterility. |
| MAAL8 | Leaves curled upward at the seedling stage. |
| MAAL9 | Leaves of light color. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Q.; Zeng, P.; Li, Z. Unraveling Large and Polyploidy Genome of the Crucifer Orychophragmus violaceus in China, a Potential Oil Crop. Plants 2023, 12, 374. https://doi.org/10.3390/plants12020374
Pan Q, Zeng P, Li Z. Unraveling Large and Polyploidy Genome of the Crucifer Orychophragmus violaceus in China, a Potential Oil Crop. Plants. 2023; 12(2):374. https://doi.org/10.3390/plants12020374
Chicago/Turabian StylePan, Qi, Pan Zeng, and Zaiyun Li. 2023. "Unraveling Large and Polyploidy Genome of the Crucifer Orychophragmus violaceus in China, a Potential Oil Crop" Plants 12, no. 2: 374. https://doi.org/10.3390/plants12020374