


Phytotoxic Action of Silver Nanoparticles on Lemna minor: Multi-Parameter Analysis of Different Physiological Processes
Abstract
:1. Introduction
2. Results and Discussion
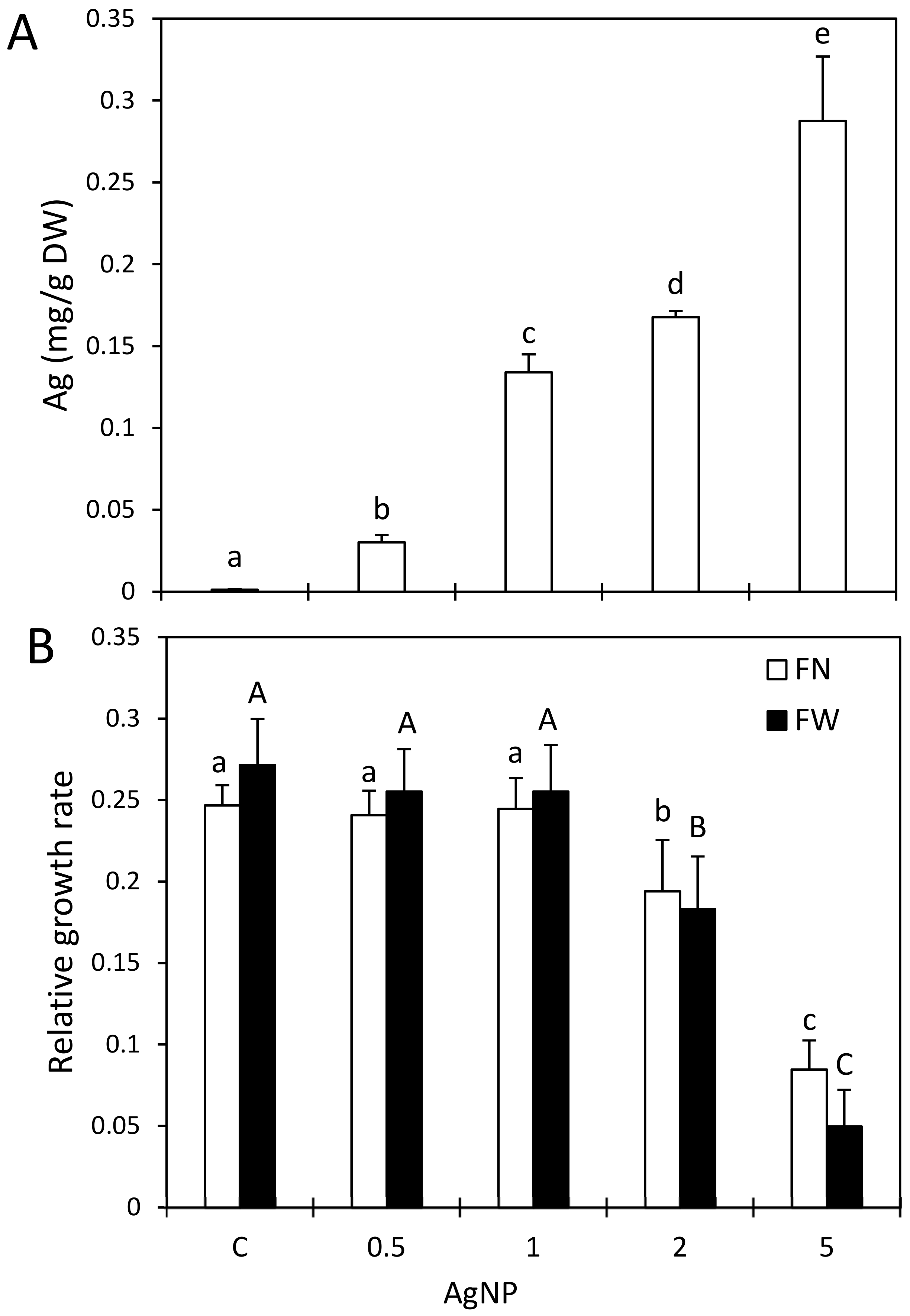
2.1. Impact of AgNPs and AgNO3 on Ag Accumulation
2.2. Impact of AgNPs on Growth and Selected Nutrients
2.3. Impact of AgNPs on Photosynthetic Pigments and Photosynthesis
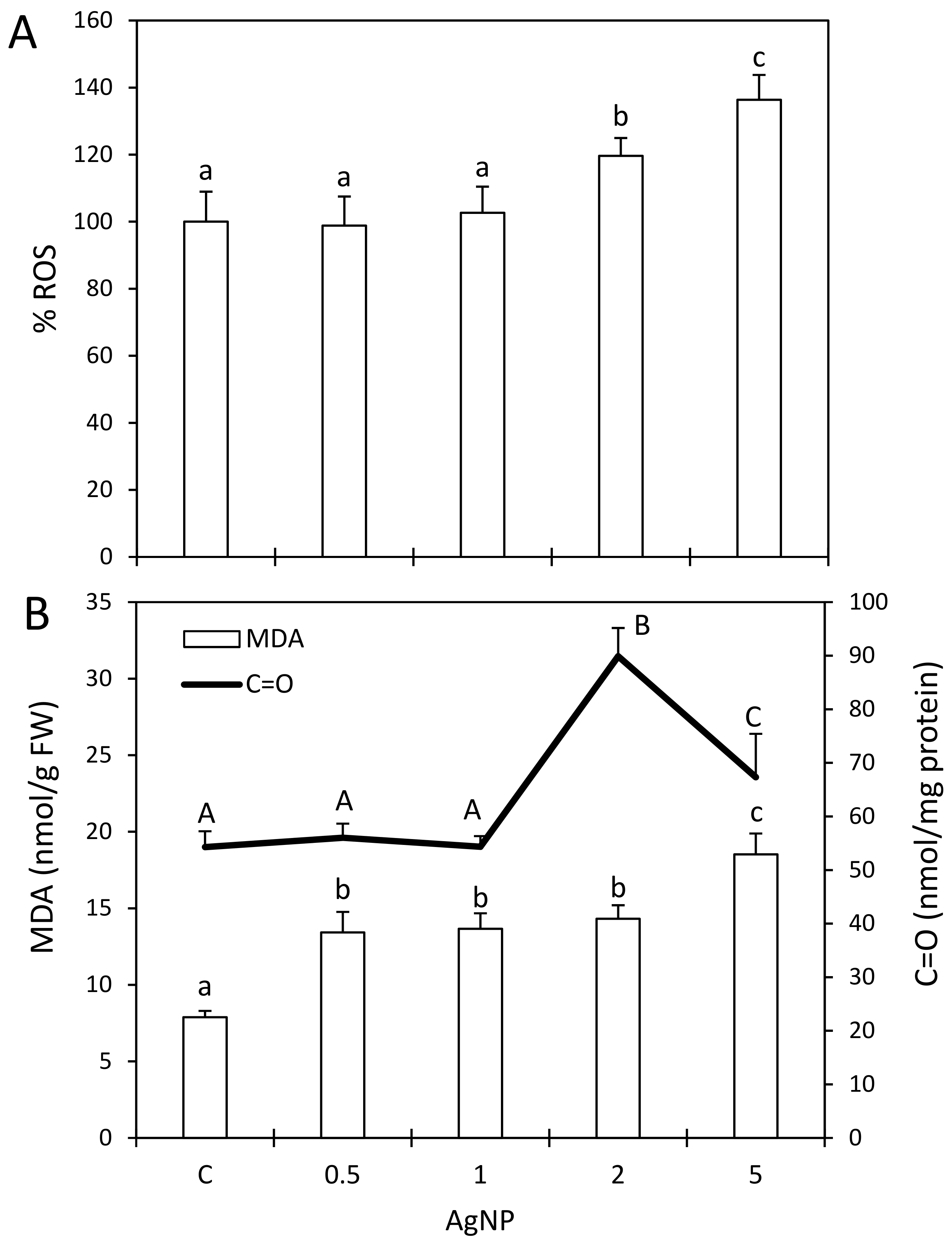
2.4. Impact of AgNPs on Cellular Redox Balance
3. Materials and Methods
3.1. Chemicals and Characterization of AgNPs in a Nutrient Medium
3.2. Plant Material and Experimental Design
3.3. Level of Ag and Selected Nutrients in Plant Material
3.4. Toxicity Parameters: Growth Rate
3.5. Measurement of Photosynthetic Pigments and Chlorophyll a Fluorescence
3.6. Oxidative Stress Parameters
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Lekamge, S.; Miranda, A.F.; Abraham, A.; Li, V.; Shukla, R.; Bansal, V.; Nugegoda, D. The toxicity of silver nanoparticles (AgNPs) to three freshwater invertebrates with different life strategies: Hydra vulgaris, Daphnia carinata, and Paratya australiensis. Front. Environ. Sci. 2018, 6, 152. [Google Scholar] [CrossRef] [Green Version]
- Angel, B.M.; Batley, G.E.; Jarolimek, C.V.; Rogers, N.J. The impact of size on the fate and toxicity of nanoparticulate silver in aquatic systems. Chemosphere 2013, 93, 359–365. [Google Scholar] [CrossRef] [PubMed]
- McShan, D.; Ray, P.C.; Yu, H. Molecular toxicity mechanism of nanosilver. J. Food Drug. Anal. 2014, 22, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvjetko, P.; Milošić, A.; Domijan, A.M.; Vinković-Vrček, I.; Tolić, S.; Peharec Štefanić, P.; Letofsky-Papst, I.; Tkalec, M.; Balen, B. Toxicity of silver ions and differently coated silver nanoparticles in Allium cepa roots. Ecotoxicol. Environ. Safe 2017, 137, 18–28. [Google Scholar] [CrossRef]
- Ratte, H.T. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. Impact of silver nanoparticles on plants: A focus on the phytotoxicity and underlying mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef]
- Pereira, S.P.P.; Jesus, F.; Aguiar, S.; de Oliviera, R.; Fernandes, M.; Renville, J.; Nogueira, A.J.A. Phytotoxicity of silver nanoparticles to Lemna minor: Surface coating and exposure period-related effects. Sci. Total. Environ. 2018, 618, 1389–1399. [Google Scholar] [CrossRef]
- Vishwakarama, K.; Upadhyay, S.N.; Singh, J.; Liu, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Tripathi, D.K.; Sharma, S. Differential phytotoxic impact of plant mediated silver nanoparticles (AgNPs) and silver nitrate (AgNO3) on Brassica sp. Front. Plant. Sci. 2017, 8, 1501. [Google Scholar] [CrossRef]
- Tolaymat, T.M.; El Badawy, A.M.; Genaidy, A.; Scheckel, K.G.; Luxton, T.P.; Suidan, M. An evidence-based environmental perspective of manufactured silver nanoparticle in syntheses and applications: A systematic review and critical appraisal of peer-reviewed scientific papers. Sci. Total. Environ. 2010, 408, 999–1006. [Google Scholar] [CrossRef]
- Radić, S.; Gregorović, G.; Stipaničev, D.; Cvjetko, P.; Šrut, M.; Vujčić, V.; Oresčanin, V.; Klobučar, G.I.V. Assessment of surface water in the vicinity of fertilizer factory using fish and plants. Ecotoxicol. Environ. Saf. 2013, 96, 32–40. [Google Scholar] [CrossRef]
- OECD. Test No. 221: Lemna sp. Growth Inhibition Test, OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2006. [Google Scholar] [CrossRef]
- Jiang, H.S.; Li, M.; Chang, F.Y.; Li, W.; Yin, L.Y. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 2012, 31, 1880–1886. [Google Scholar] [CrossRef]
- Ke, M.; Qu, Q.; Peijnenburg, W.; Li, X.; Zhang, M.; Zhang, Z.; Lu, T.; Pan, X.; Qian, H. Phytotoxic effects of silver nanoparticles and silver ions to Arabidopsis thaliana as revealed by analysis of molecular responses and of metabolic pathways. Sci. Total Environ. 2018, 644, 1070–1079. [Google Scholar] [CrossRef]
- Peharec Štefanić, P.; Košpić, K.; Lyons, D.M.; Jurković, L.; Balen, B.; Tkalec, M. Phytotoxicity of silver nanoparticles on tobacco plants: Evaluation of coating effects on photosynthetic performance and chloroplast ultrastructure. Nanomaterials 2021, 11, 744. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Peng, X.; Han, X.; Ren, J.; Sun, L.; Fu, Z. Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana. J. Environ. Sci. 2013, 25, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Yin, L.Y.; Ren, N.N.; Zhao, S.T.; Li, Z.; Zhi, Y.; Shao, H.; Li, W.; Gontero, B. Silver nanoparticles induced reactive oxygen species via photosynthetic energy transport imbalance in an aquatic plant. Nanotoxicology 2017, 11, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Dewez, D.; Goltsev, V.; Kalaji, H.M.; Oukarroum, A. Inhibitory effects of silver nanoparticles on photosystem II performance in Lemna gibba probed by chlorophyll fluorescence. Curr. Plant Biol. 2018, 16, 15–21. [Google Scholar] [CrossRef]
- Jiang, H.S.; Qiu, X.N.; Li, G.B.; Li, W.; Yin, L.Y. Silver nanoparticles induced accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 2014, 33, 1398–1405. [Google Scholar] [CrossRef]
- Radić, S.; Domijan, A.M.; Glavaš Ljubimir, K.; Maldini, K.; Ivešić, M.; Peharec Štefanić, P.; Krivohlavek, A. Toxicity of nanosilver and fumonisin B1 and their interactions on duckweed (Lemna minor L.). Chemosphere 2019, 229, 86–93. [Google Scholar] [CrossRef]
- Iannelli, M.A.; Bellini, A.; Venditti, I.; Casentini, B.; Battocchio, C.; Scalici, M.; Ceschin, S. Differential phytotoxic effect of silver nitrate (AgNO3) and bifunctionalized silver nanoparticles (AgNPs-Cit-L-Cys) on Lemna plants (duckweeds). Aquat. Toxicol. 2022, 250, 106260. [Google Scholar] [CrossRef]
- Gubbins, E.J.; Batty, L.C.; Lead, J.R. Phytotoxicity of silver nanoparticles to Lemna minor L. Environ. Pollut. 2011, 159, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Geisler-Lee, J.; Wang, Q.; Yao, Y.; Zhang, W.; Geisler, M.; Li, K.; Huang, Y.; Chen, Y.; Kolmakov, A.; Ma, X. Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology 2013, 7, 323–337. [Google Scholar] [CrossRef]
- Li, C.C.; Dang, F.; Li, M.; Zhu, M.; Zhong, H.; Hintelmann, H.; Zhou, D.M. Effects of exposure pathways on the accumulation and phytotoxicity of silver nanoparticles in soybean and rice. Nanotoxicology 2017, 11, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Zuverza-Mena, N.; Armendariz, R.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Effects of silver nanoparticles on radish sprouts: Root growth reduction and modifications in the nutritional value. Front. Plant. Sci. 2016, 7, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klíma, P.; Laňková, M.; Vandenbussche, F.; Van Der Straeten, D.; Petrášek, J. Silver ions increase plasma membrane permeability through modulation of intracellular calcium levels in tobacco BY-2 cells. Plant Cell Rep. 2018, 37, 809–818. [Google Scholar] [CrossRef]
- Cvjetko, P.; Zovko, M.; Peharec Štefanić, P.; Biba, R.; Tkalec, M.; Domijan, A.-M.; Vinković-Vrček, I.; Letofsky-Papst, I.; Šikić, S.; Balen, B. Phytotoxic effects of silver nanoparticles in tobacco plants. Environ. Sci. Pollut. Res. 2018, 25, 5590–5602. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Chen, X.; Song, G.; Wang, Q.; Chang, C.C. Effects of copper and cadmium on heavy metal polluted waterbody restoration by duckweed (Lemna minor). Plant Physiol. Biochem. 2007, 45, 62–69. [Google Scholar] [CrossRef]
- Shabnam, N.; Sharmila, P.; Pardha-Saradhi, P. Impact of ionic and nanoparticle speciation states of silver on light harnessing photosynthetic events in Spirodela polyrhiza. Int. J. Phytoremediat. 2017, 19, 80–86. [Google Scholar] [CrossRef]
- Jansson, H.; Hansson, Ö. Competitive inhibition of electron donation to photosystem 1 by metal-substituted plastocyanin. Biochim. Biophys. Acta 2008, 1777, 1116–1121. [Google Scholar] [CrossRef]
- Joliot, P.; Johnson, G.N. Regulation of cyclic and linear electron flow in higher plants. Proc. Natl. Acad. Sci. USA 2011, 108, 13317–13322. [Google Scholar] [CrossRef] [Green Version]
- van Doorn, W.G.; Ketsa, S. Cross reactivity between ascorbate peroxidase and phenol (guaiacol) peroxidase. Postharvest Biol. Technol. 2014, 95, 64–69. [Google Scholar] [CrossRef]
- Ederli, L.; Reale, L.; Ferranti, F.; Pasqualini, S. Responses induced by high concentration of cadmium in Phragmites australis roots. Physiol. Plant 2004, 121, 66–74. [Google Scholar] [CrossRef]
- Halušková, L.; Valentovičová, K.; Huttová, J.; Mistrík, I.; Tamás, L. Effect of heavy metals on root growth and peroxidase activity in barley root tip. Acta Physiol. Plant 2010, 32, 59–65. [Google Scholar] [CrossRef]
- Gao, J.; Chen, B.; Lin, H.; Liu, Y.; Wei, Y.; Chen, F.; Li, W. Identification and characterization of the glutathione S-Transferase (GST) family in radish reveals a likely role in anthocyanin biosynthesis and heavy metal stress tolerance. Gene 2020, 743, 144484. [Google Scholar] [CrossRef] [PubMed]
- Pirson, A.; Seidel, F. Zell-und stoffwechselphysiologiche Untersuchngen an der Wurzel von Lemna minor unter besonderer Berűcksichtigung von Kalium-und Calciummangel. Planta 1950, 38, 431–473. [Google Scholar] [CrossRef]
- ISO/TR 11905; Water Quality—Determination of Nitrogen—Part 2: Determination of Bound Nitrogen, after Combustion and Oxidation to Nitrogen Dioxide, Chemiluminescence Detection. ISO: Geneva, Switzerland, 1997.
- ISO 6878; Water Quality—Determination of Phosphorus—Ammonium Molybdate Spectrometric Method. ISO: Geneva, Switzerland, 2004.
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids, the pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutase. I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. Assay for differentation of glutathione S-transferases. Methods Enzymol. 1981, 77, 398–405. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| mg/L | N | P | K | Mg | Ca |
|---|---|---|---|---|---|
| C | 41.68 (1.10) a | 22.80 (1.25) a | 54.10 (0.94) a | 2.58 (0.058) a | 13.94 (0.54) b |
| AgNP | |||||
| 0.5 | 40.11 (0.82) ab | 20.72 (1.57) a | 54.01 (4.86) a | 2.60 (0.042) a | 14.82 (0.69) b |
| 1 | 40.26 (0.55) ab | 23.20 (1.72) ab | 46.67 (3.16) b | 2.31 (0.043) b | 14.77 (0.23) b |
| 2 | 38.65 (1.45) b | 21.38 (1.77) ab | 46.55 (3.26) b | 2.33 (0.154) b | 15.11 (0.85) ab |
| 5 | 39.48 (0.44) b | 18.87 (2.96) b | 41.18 (2.12) b | 2.25 (0.079) b | 16.21 (0.92) a |
| mg/L | Fv/Fm | ΦPSII | NPQ | Chl a | Chl b | Car |
|---|---|---|---|---|---|---|
| C | 0.67 (0.009) a | 0.58 (0.017) a | 0.061 (0.011) b | 0.71 (0.022) a | 0.27 (0.014) a | 0.29 (0.008) a |
| AgNP | ||||||
| 0.5 | 0.66 (0.011) a | 0.56 (0.015) a | 0.053 (0.007) b | 0.70 (0.019) a | 0.28 (0.008) a | 0.29 (0.008) a |
| 1 | 0.66 (0.009) a | 0.56 (0.017) ab | 0.052 (0.017) b | 0.69 (0.006) a | 0.26 (0.005) a | 0.28 (0.010) a |
| 2 | 0.64 (0.009) b | 0.55 (0.016) b | 0.073 (0.008) ab | 0.63 (0.020) b | 0.24 (0.014) b | 0.26 (0.017) b |
| 5 | 0.63 (0.009) b | 0.54 (0.017) b | 0.088 (0.011) a | 0.62 (0.014) b | 0.22 (0.024) c | 0.24 (0.023) c |
| mg/L | SOD | APX | CAT | GPX | GST | GSH |
|---|---|---|---|---|---|---|
| C | 15.1 (0.77) c | 0.97 (0.064) b | 0.15 (0.008) c | 0.40 (0.023) c | 0.27 (0.013) c | 100.0 (5.31) a |
| AgNP | ||||||
| 0.5 | 17.4 (0.51) b | 0.96 (0.072) b | 0.17 (0.009) bc | 0.46 (0.041) c | 0.29 (0.021) c | 94.0 (3.84) a |
| 1 | 15.9 (0.72) c | 1.14 (0.037) a | 0.17 (0.008) b | 0.60 (0.028) b | 0.34 (0.006) b | 86.4 (4.22) b |
| 2 | 20.8 (1.62) a | 1.12 (0.096) a | 0.24 (0.008) a | 0.57 (0.033) b | 0.38 (0.051) b | 84.0 (2.71) b |
| 5 | 19.5 (1.30) a | 1.13 (0.052) a | 0.25 (0.010) a | 0.72 (0.082) a | 0.45 (0.035) a | 82.2 (2.03) b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glavaš Ljubimir, K.; Domijan, A.-M.; Radić Brkanac, S. Phytotoxic Action of Silver Nanoparticles on Lemna minor: Multi-Parameter Analysis of Different Physiological Processes. Plants 2023, 12, 343. https://doi.org/10.3390/plants12020343
Glavaš Ljubimir K, Domijan A-M, Radić Brkanac S. Phytotoxic Action of Silver Nanoparticles on Lemna minor: Multi-Parameter Analysis of Different Physiological Processes. Plants. 2023; 12(2):343. https://doi.org/10.3390/plants12020343
Chicago/Turabian StyleGlavaš Ljubimir, Katarina, Ana-Marija Domijan, and Sandra Radić Brkanac. 2023. "Phytotoxic Action of Silver Nanoparticles on Lemna minor: Multi-Parameter Analysis of Different Physiological Processes" Plants 12, no. 2: 343. https://doi.org/10.3390/plants12020343