
Evaluation of the Effect of Organic Matter from Invasive Plants on Soil Nematode Communities

Abstract
:1. Introduction
2. Results
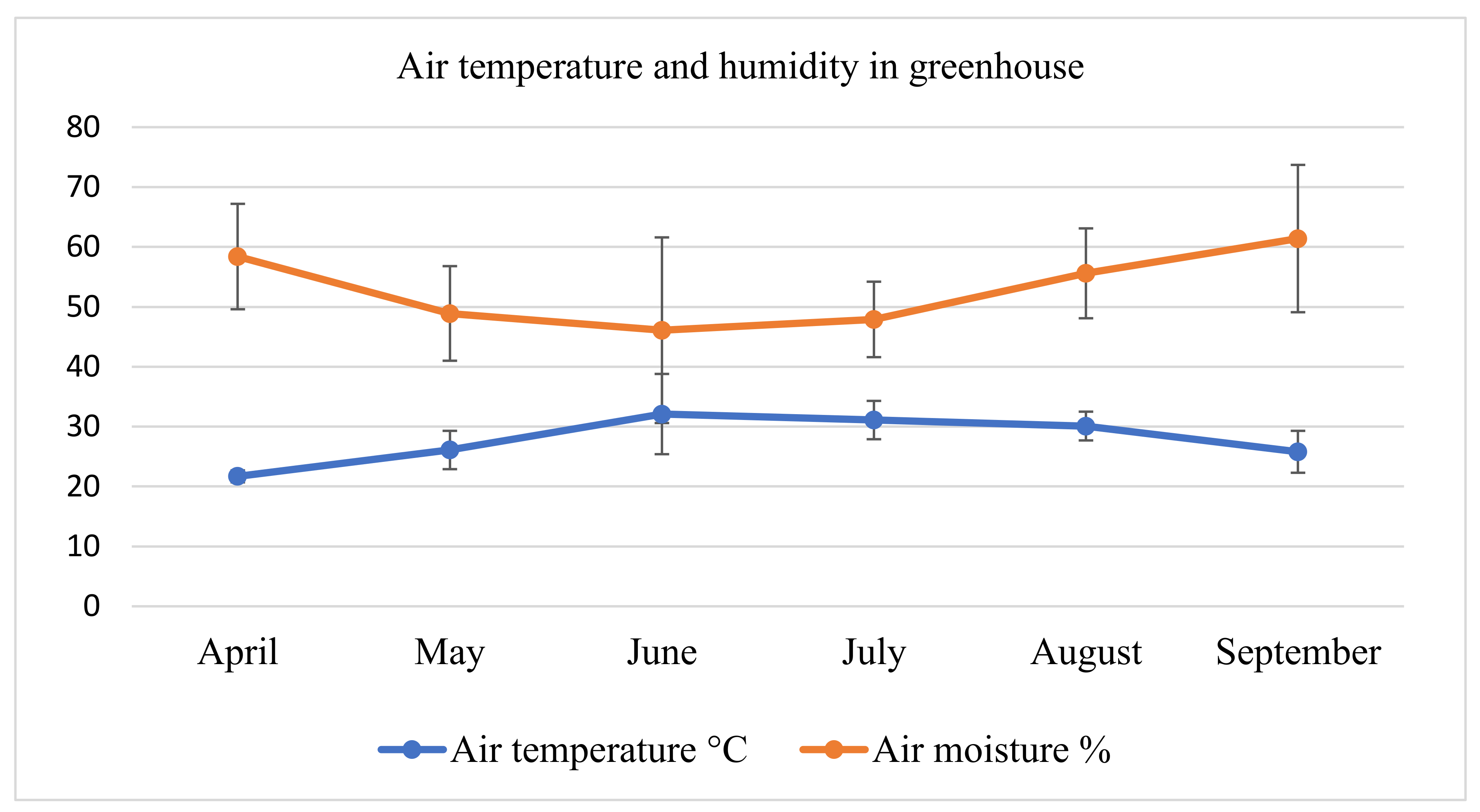
2.1. Soil Physical and Chemical Properties
2.2. Total Number of Genera and Composition, Abundance, Diversity Index and Biomass of the Nematode Communities
2.3. Ecological and Functional Indices of Nematode Communities
2.4. Nematode Abundance in the Trophic and cp Groups
3. Discussion
3.1. OMF and S/OMF
3.2. OMS and S/OMS
4. Material and Methods
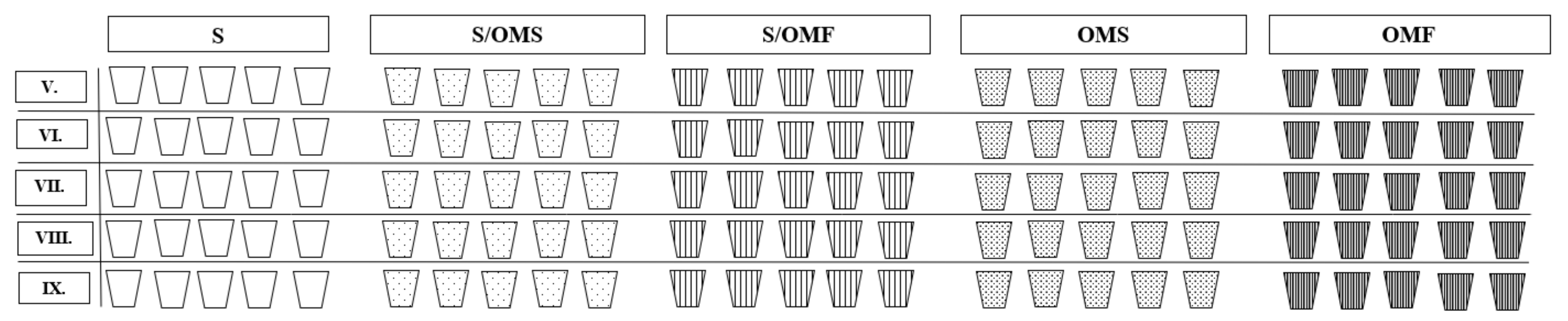
4.1. Experimental Plots, Litter and Soil Sampling
4.2. Soil Properties
4.3. Nematode Extraction, Identification and Evaluation
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 1–17. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Callaway, R.M. Direct and Indirect Effects of Invasive Plants on Soil Chemistry and Ecosystem Function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef]
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- McLeod, M.L.; Bullington, L.; Cleveland, C.C.; Rousk, J.; Lekberg, Y. Invasive plant-derived dissolved organic matter alters microbial communities and carbon cycling in soils. Soil Biol. Biochem. 2021, 156, 108191. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef]
- Wakjira, M.; Berecha, G.; Tulu, S. Allelopathic effects of an invasive alien weed Parthenium hysterophorus L. compost on lettuce germination and growth. Afr. J. Agric. Res. 2009, 4, 1325–1330. [Google Scholar]
- Jiao, Y.; Li, Y.; Yuan, L.; Huang, J. Allelopathy of uncomposted and composted invasive aster (Ageratina adenophora) on ryegrass. J. Hazard. Mater. 2021, 402, 123727. [Google Scholar] [CrossRef]
- Das, M.B.B.; Acharya, B.D.; Saquib, M.; Chettri, M.K. Effect of aqueous extract and compost of invasive weed Ageratina adenophora on seed germination and seedling growth of some crops and weeds. J. Biodivers. Conserv. Bioresour. Manag. 2018, 4, 11–20. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000. [Google Scholar]
- Van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.I.; González-Moreno, P.; Groom, Q.J.; Hulme, P.; et al. The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef]
- Shaw, R.H.; Seiger, L.A. Japanese Knotweed. In Biological Control of Invasive Plants in the Eastern United States; Van Driesche, R., Lyon, S., Blossey, B., Hoddle, M., Reardon, R., Eds.; US Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2002; pp. 159–166. [Google Scholar]
- Richards, C.L.; Walls, R.L.; Bailey, J.P.; Parameswaran, R.; George, T.; Pigliucci, M. Plasticity in salt tolerance traits allows for invasion of novel habitat by Japanese knotweed s.l. (Fallopia japonica and F.×bohemica, Polygonaceae). Am. J. Bot. 2008, 95, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Prather, T.S.; Miller, T.W.; Robins, S. Knotweed Shrubs: Identification, Biology, and Management; University of Idaho Extension: Moscow, ID, USA, 2009. [Google Scholar]
- Abgrall, C.; Forey, E.; Mignot, L.; Chauvat, M. Invasion by Fallopia japonica alters soil food webs through secondary metabolites. Soil Biol. Biochem. 2018, 127, 100–109. [Google Scholar] [CrossRef]
- Pal, R.W.; Chen, S.; Nagy, D.U.; Callaway, R.M. Impacts of Solidago gigantea on other species at home and away. Biol. Invasions 2015, 17, 3317–3325. [Google Scholar] [CrossRef]
- Nagy, D.U.; Stranczinger, S.; Godi, A.; Weisz, A.; Rosche, C.; Suda, J.; Mariano, M.; Pal, R.W. Does higher ploidy level increase the risk of invasion? A case study with two geo-cytotypes of Solidago gigantea Aiton (Asteraceae). J. Plant Ecol. 2018, 11, 317–327. [Google Scholar]
- Benelli, G.; Pavela, R.; Cianfaglione, K.; Nagy, D.U.; Canale, A.; Maggi, F. Evaluation of two invasive plant invaders in Europe (Solidago canadensis and Solidago gigantea) as possible sources of botanical insecticides. J. Pest Sci. 2019, 92, 805–821. [Google Scholar] [CrossRef]
- Jakobs, G.; Weber, E.; Edwards, P.J. Introduced plants of the invasive Solidago gigantea (Asteraceae) are larger and grow denser than conspecifics in the native range. Divers. Distrib. 2004, 10, 11–19. [Google Scholar]
- Chapuis-Lardy, L.; Vanderhoeven, S.O.N.I.A.; Dassonville, N.; Koutika, L.S.; Meerts, P. Effect of the exotic invasive plant Solidago gigantea on soil phosphorus status. Biol. Fertil. Soils 2006, 42, 481–489. [Google Scholar] [CrossRef]
- Scharfy, D.; Güsewell, S.; Gessner, M.O.; Venterink, H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: Effects on soil microbes, nutrients and plant–soil feedbacks. J. Ecol. 2010, 98, 1379–1388. [Google Scholar] [CrossRef]
- Bobuľská, L.; Demková, L.; Čerevková, A.; Renčo, M. Invasive goldenrod Solidago gigantea influences soil microbial activities in forest and grassland ecosystems in central Europe. Diversity 2019, 11, 134. [Google Scholar] [CrossRef]
- Bongers, T. The Maturity Index, the evolution of nematode life history traits, adaptive radiation and CP-scaling. Plant Soil 1990, 212, 13–22. [Google Scholar] [CrossRef]
- Du Preez, G.; Daneel, M.; De Goede, R.; Du Toit, M.J.; Ferris, H.; Fourie, H.; Geisen, S.; Kakouli-Duarte, T.; Korthals, G.; Sánchez-Moreno, S.; et al. Nematode-based indices in soil ecology: Application, utility, and future directions. Soil Biol. Biochem. 2022, 169, 108640. [Google Scholar] [CrossRef]
- Čerevková, A.; Bobuľská, L.; Miklisová, D.; Renčo, M. A case study of soil food web components affected by Fallopia japonica Polygonaceae in three natural habitats in Central Europe. J. Nematol. 2019, 51, e2019–e2042. [Google Scholar] [CrossRef]
- Renčo, M.; Čerevková, A.; Homolová, Z. Nematode communities indicate the negative impact of Reynoutria japonica invasion on soil fauna in ruderal habitats of tatra national park in Slovakia. Glob. Ecol. Conserv. 2021, 26, e01470. [Google Scholar] [CrossRef]
- Čerevková, A.; Miklisová, D.; Bobuľská, L.; Renčo, M. Impact of the invasive plant Solidago gigantea on soil nematodes in a semi-natural grassland and a temperate broadleaved mixed forest. J. Helminthol. 2020, 94, e51. [Google Scholar] [CrossRef] [PubMed]
- Quist, C.W.; Vervoort, M.T.; Van Megen, H.; Gort, G.; Bakker, J.; Van der Putten, W.H.; Helder, J. Selective alteration of soil food web components by invasive giant goldenrod Solidago gigantea in two distinct habitat types. Oikos 2014, 123, 837–845. [Google Scholar] [CrossRef]
- Harkes, P.; van Heumen, L.J.; van den Elsen, S.J.; Mooijman, P.J.; Vervoort, M.T.; Gort, G.; Holterman, M.H.M.; van Steenbrugge, J.J.M.; Quist, C.W.; Helder, J. Characterization of the Habitat- and Season-Independent Increase in Fungal Biomass Induced by the Invasive Giant Goldenrod and Its Impact on the Fungivorous Nematode Community. Microorganisms 2021, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Steel, H.; de la Peña, E.; Fonderie, P.; Willekens, K.; Borgonie, G.; Bert, W. Nematode succession during composting and the potential of the nematode community as an indicator of compost maturity. Pedobiologia 2010, 53, 181–190. [Google Scholar] [CrossRef]
- Parepa, M.; Bossdorf, O. Testing for allelopathy in invasive plants: It all depends on the substrate! Biol. Invasions 2016, 18, 2975–2982. [Google Scholar] [CrossRef]
- Dassonville, N.; Guillaumaud, N.; Piola, F.; Meerts, P.; Poly, F. Niche construction by the invasive Asian knotweeds (species complex Fallopia): Impact on activity, abundance and community structure of denitrifies and nitrifies. Biol. Invasions 2011, 13, 1115–1133. [Google Scholar] [CrossRef]
- Stoll, P.; Gatzsch, K.; Rusterholz, H.P.; Baur, B. Response of plant and gastropod species to knotweed invasion. Basic Appl. Ecol. 2012, 13, 232–240. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Gruber, W.; Meerts, P. Invasion by Fallopia japonica increases topsoil mineral nutrient concentrations. Ecoscience 2007, 14, 230–240. [Google Scholar] [CrossRef]
- Cvejić, R.; Klages, S.; Pintar, M.; Resman, L.; Slatnar, A.; Mihelič, R. Invasive Plants in Support of Urban Farming: Fermentation-Based Organic Fertilizer from Japanese Knotweed. Agronomy 2021, 11, 1232. [Google Scholar] [CrossRef]
- De Almeida, T.; Forey, E.; Chauvat, M. Alien invasive plant effect on soil fauna is habitat dependent. Diversity 2022, 14, 61. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Stanek, M.; Majewska, M.L.; Nobis, M.; Zubek, S. Invasive plant species identity affects soil microbial communities in a mesocosm experiment. Appl. Soil Ecol. 2019, 136, 168–177. [Google Scholar] [CrossRef]
- Daayf, F.; Schmitt, A.; Belanger, R.R. The Effects of Plant Extracts of Reynoutria sachalinensis on Powdery Mildew Development and Leaf Physiology of Long English Cucumber. Plant Dis. 1995, 79, 577–580. [Google Scholar] [CrossRef]
- Aguilera, A.G.; Alpert, P.; Dukes, J.S.; Harrington, R. Impacts of the invasive plant Fallopia japonica (Houtt.) on plant communities and ecosystem processes. Biol. Invasions 2010, 12, 1243–1252. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Campos-Herrera, R.; Sánchez-Moreno, S.; Vittoz, P.; Rasmann, S. The Abundance, Diversity, and Metabolic Footprint of Soil Nematodes Is Highest in High Elevation Alpine Grasslands. Front. Ecol. Evol. 2016, 4, 84. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, W.; Wang, B.; Tang, J.; Chen, X. Secondary metabolites from the invasive Solidago canadensis L. accumulation in soil and contribution to inhibition of soil pathogen Pythium ultimum. Appl. Soil Ecol. 2011, 48, 280–286. [Google Scholar] [CrossRef]
- Carson, W.P.; Peterson, C.J. The role of litter in an old-field community: Impact of litter quantity in different seasons on plant species richness and abundance. Oecologia 1990, 85, 8–13. [Google Scholar] [CrossRef]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 103, 239–251. [Google Scholar] [CrossRef]
- Taylor, L.S.; Phillips, G.; Bernard, E.C.; DeBruyn, J.M. Soil nematode functional diversity, successional patterns, and indicator taxa associated with vertebrate decomposition hotspots. PLoS ONE 2020, 15, e0241777. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Biol. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, K.; Zhou, J.; Wu, B. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China. Sci. Total Environ. 2018, 631, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Klimek, B.; Jaźwa, M.; Gołębiewski, M.; Sikora, M.; Deja-Sikora, E. No apparent effect of invasive alien goldenrod on soil microbial communities or soil fauna feeding activity. Acta Oecologica 2020, 109, 103669. [Google Scholar] [CrossRef]
- Majewska, M.L.; Rola, K.; Zubek, S. The growth and phosphorus acquisition of invasive plants Rudbeckia laciniata and Solidago gigantea are enhanced by arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Matute, M.M. Structural and functional succession in the nematode fauna of a soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Bochníček, O. Climate Atlas of Slovakia; Slovak Hydrometeorological Institute: Bratislava, Slovakia, 2015. [Google Scholar]
- Miklós, L. Landscape Atlas of the Slovak Republic; Ministry of Environment of the Slovak Republic, Slovak Environmental Agency: Banská Bystrica, Slovakia, 2002. [Google Scholar]
- Fiala, K.; Barančíková, G.; Brečková, V.; Burik, V.; Houšková, B.; Chomaničová, A.; Kobza, J.; Litavec, T.; Makovníková, L.; Pechová, B.; et al. Partial Monitoring System–Soil, Binding Methods (in Slovak); VÚPOP: Bratislava, Slovakia, 1999. [Google Scholar]
- Van Bezooijen, J. Methods and Techniques for Nematology; Wageningen University: Wageningen, The Netherlands, 2006. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G. NINJA: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.; Freckman, D.W.; Georgieva, S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Wasilewska, L. Soil invertebrates as bioindicators, with special reference to soil-inhabiting nematodes. Russ. J. Nematol. 1997, 5, 113–126. [Google Scholar]
- Ettema, C.H.; Bongers, T. Characterization of nematode colonization and succession in disturbed soil using the Maturity Index. Biol. Fertil. Soils 1993, 16, 79–85. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]



{kind=link}
{kind=link}
| Measured Parameters | S | S/OMF | S/OMS | OMF | OMS | p |
|---|---|---|---|---|---|---|
| pH | 7.6 ± 0.1 a | 7.1 ± 0.1 b | 7.0 ± 0.0 b | 6.3 ± 0.5 c | 6.3 ± 0.5 c | *** |
| Moisture | 29.6 ± 7.3 a | 24.2 ± 9.2 b | 28.4 ± 12.5 ab | 31.4 ± 7.4 ac | 36.5 ± 9.5 c | *** |
| Organic carbon | 7.2 ± 0.65 a | 15.0 ± 0.97 b | 14.8 ± 0.50 b | 9.15 ± 0.12 c | 9.35 ± 0.46 c | ** |
| Total nitrogen | 1.11 ± 0.06 a | 1.79 ± 0.08 b | 1.81 ± 0.14 b | 1.19 ± 0.06 a | 1.32 ± 0.09 ab | ** |
| S | S/OMF | S/OMS | OMF | OMS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nematode Genera | c-p | A ± SD | D% | A ± SD | D% | A ± SD | D% | A ± SD | D% | A ± SD | D% |
| Bacterivores | |||||||||||
| Acrobeles | 2 | 0.1 ± 0.4 | 0.07 | 0.1 ± 0.26 | 0.04 | 0.1 ± 0.39 | 0.06 | 0.2 ± 0.92 | 0.14 | 0.3 ± 1.12 | 0.19 |
| Acrobeloides | 2 | 6.8 ± 4.95 | 3.99 | 14.2 ± 12.09 | 9.69 | 19.3 ± 19.93 | 10.74 | 16 ± 16.28 | 12.31 | 12 ± 14.03 | 8.11 |
| Alaimus | 4 | 2.5 ± 3.68 | 1.5 | 0.3 ± 0.83 | 0.22 | 0.9 ± 2.05 | 0.5 | 0.2 ± 0.49 | 0.14 | - | - |
| Aulolaimus | 3 | - | - | 0 ± 0.24 | 0.03 | - | - | - | - | - | - |
| Cephalobus | 2 | 8.5 ± 11.89 | 4.98 | 4.3 ± 4.83 | 2.92 | 3.6 ± 6.26 | 2.01 | 0.4 ± 0.82 | 0.32 | 1.5 ± 2.75 | 1.04 |
| Cervidellus | 2 | 0.2 ± 0.6 | 0.12 | 1 ± 2.11 | 0.66 | 0.4 ± 0.69 | 0.2 | 1.9 ± 3.44 | 1.47 | 1 ± 1.77 | 0.7 |
| Diploscapter | 1 | 0.4 ± 0.96 | 0.25 | 0.3 ± 0.83 | 0.17 | 1.3 ± 2.31 | 0.73 | 1 ± 3.46 | 0.78 | 0.8 ± 2.42 | 0.55 |
| Eucephalobus | 3 | 7.5 ± 6.84 | 4.44 | 10.4 ± 10.25 | 7.12 | 8.2 ± 6.18 | 4.57 | 6.9 ± 8.1 | 5.32 | 2.1 ± 2.25 | 1.44 |
| Chiloplacus | 2 | 0 ± 0.24 | 0.03 | 0.1 ± 0.32 | 0.04 | 1.1 ± 2.75 | 0.6 | - | - | - | - |
| Monhystera | 2 | - | - | 0.1 ± 0.32 | 0.06 | - | - | 0.1 ± 0.26 | 0.04 | - | - |
| Panagrolaimus | 1 | 0.1 ± 0.34 | 0.04 | 0.1 ± 0.34 | 0.05 | - | - | - | - | - | - |
| Pelodera | 1 | 3.4 ± 4.6 | 2.01 | 10.9 ± 17.07 | 7.44 | 4.6 ± 6 | 2.56 | 7.1 ± 11.1 | 5.5 | 4.6 ± 7.07 | 3.1 |
| Plectus | 2 | 2.7 ± 2.87 | 1.58 | 2.6 ± 3.12 | 1.77 | 1.4 ± 2.14 | 0.8 | 1 ± 1.16 | 0.77 | 0.7 ± 1.39 | 0.45 |
| Prismatolaimus | 3 | 0.4 ± 0.77 | 0.21 | 0.5 ± 2.35 | 0.36 | 0.4 ± 1.02 | 0.23 | 0.1 ± 0.26 | 0.04 | 0 ± 0.24 | 0.03 |
| Protorhabditis | 1 | - | - | 0.2 ± 0.5 | 0.12 | 1.2 ± 3.23 | 0.68 | - | - | - | - |
| Rhabditis | 1 | 30.6 ± 16.5 | 18 | 28 ± 21.42 | 19.16 | 48.9 ± 54.5 | 27.14 | 54.4 ± 35.08 | 41.89 | 82.8 ± 89.41 | 55.87 |
| Wilsonema | 2 | 0.2 ± 0.58 | 0.15 | 0.2 ± 0.46 | 0.11 | 0.1 ± 0.55 | 0.08 | 0.1 ± 0.4 | 0.09 | - | - |
| Fungivores | |||||||||||
| Aphelenchoides | 2 | 1.8 ± 2.08 | 1.08 | 6.5 ± 5.42 | 4.43 | 13.2 ± 10.49 | 7.32 | 3.7 ± 3.36 | 2.84 | 10.7 ± 8.73 | 7.2 |
| Aphelenchus | 2 | 9.4 ± 8.85 | 5.51 | 6.2 ± 4.5 | 4.22 | 5.4 ± 4.79 | 3 | 1.7 ± 4.78 | 1.28 | 0.9 ± 1.76 | 0.63 |
| Diphtherophora | 3 | 1.6 ± 1.61 | 0.93 | 0.7 ± 1.39 | 0.48 | 0.9 ± 2.68 | 0.52 | 0.5 ± 1.33 | 0.36 | - | - |
| Filenchus | 2 | 11.8 ± 19.77 | 6.96 | 11.3 ± 17.91 | 7.7 | 19.5 ± 25.21 | 10.85 | 1.6 ± 3.01 | 1.2 | 6.6 ± 12.41 | 4.44 |
| Tylencholaimus | 4 | 11.4 ± 6.92 | 6.7 | 11.3 ± 13.67 | 7.7 | 13.9 ± 18.74 | 7.69 | 0.1 ± 0.28 | 0.04 | 0.1 ± 0.44 | 0.09 |
| Herbivores | |||||||||||
| Aglenchus | 2 | 0.3 ± 1.72 | 0.2 | - | - | - | - | - | - | - | - |
| Axonchium | 5 | - | - | - | - | 0 ± 0.24 | 0.03 | - | - | - | - |
| Boleodorus | 2 | 0.7 ± 1.73 | 0.41 | 0.3 ± 0.74 | 0.17 | - | - | - | - | - | - |
| Criconema | 3 | 1.6 ± 2.55 | 0.93 | 1 ± 2.2 | 0.69 | 0.4 ± 1.29 | 0.23 | - | - | - | - |
| Helicotylenchus | 3 | 23.7 ± 10.65 | 13.94 | 7.2 ± 7.19 | 4.89 | 9.2 ± 10.35 | 5.12 | 0.1 ± 0.39 | 0.09 | 0.2 ± 0.58 | 0.14 |
| Heterodera | 3 | 2.6 ± 6.56 | 1.54 | 1.1 ± 3.82 | 0.77 | 1 ± 1.86 | 0.55 | - | - | - | - |
| Paratylenchus | 2 | 10.9 ± 10.03 | 6.39 | 5.3 ± 8.23 | 3.62 | 4.8 ± 5.41 | 2.66 | 0.1 ± 0.37 | 0.08 | 0.1 ± 0.48 | 0.09 |
| Pratylenchoides | 3 | - | - | - | - | 0.1 ± 0.34 | 0.04 | - | - | - | - |
| Pratylenchus | 3 | 0.4 ± 1.18 | 0.26 | 0.7 ± 1.49 | 0.48 | 0.3 ± 1.11 | 0.19 | 0.1 ± 0.5 | 0.11 | 0.3 ± 1.12 | 0.19 |
| Rotylenchus | 3 | 10 ± 7.85 | 5.91 | 1.7 ± 1.67 | 1.15 | 3.6 ± 4.01 | 1.99 | - | - | 0.1 ± 0.34 | 0.05 |
| Trichodorus | 4 | 0.4 ± 1.63 | 0.26 | - | - | 0.2 ± 0.72 | 0.11 | 0.1 ± 0.28 | 0.04 | - | - |
| Tylenchorhynchus | 3 | 8.4 ± 11.76 | 4.93 | 2.3 ± 3.03 | 1.57 | 7.8 ± 8.59 | 4.35 | - | - | - | - |
| Tylenchus | 2 | 0.1 ± 0.34 | 0.04 | 0.2 ± 0.59 | 0.11 | - | - | - | - | - | - |
| Omnivores | |||||||||||
| Discolaimus | 4 | - | - | - | - | - | - | 0.1 ± 0.28 | 0.04 | - | - |
| Dorylaimus | 4 | 0.1 ± 0.34 | 0.04 | - | - | - | - | - | - | - | - |
| Enchodelus | 4 | 0.7 ± 1.56 | 0.44 | 0.3 ± 0.77 | 0.2 | - | - | 0.1 ± 0.28 | 0.04 | 0.1 ± 0.3 | 0.04 |
| Eudorylaimus | 4 | 6.2 ± 7.18 | 3.63 | 6.9 ± 9.11 | 4.7 | 1.9 ± 3.09 | 1.05 | 19.6 ± 28.47 | 15.11 | 0.9 ± 1.75 | 0.61 |
| Mesodorylaimus | 4 | 0.2 ± 0.73 | 0.14 | 0.1 ± 0.26 | 0.04 | - | - | - | - | 0.1 ± 0.36 | 0.05 |
| Predators | |||||||||||
| Anatonchus | 4 | 0.1 ± 0.32 | 0.04 | 0.1 ± 0.44 | 0.09 | - | - | - | - | 0.1 ± 0.42 | 0.08 |
| Clarkus | 4 | 0.1 ± 0.74 | 0.09 | 0.1 ± 0.26 | 0.04 | 0.1 ± 0.53 | 0.08 | 0.2 ± 0.69 | 0.18 | 0.3 ± 0.72 | 0.22 |
| Iotonchus | 4 | 0.1 ± 0.32 | 0.04 | - | - | - | - | - | - | - | - |
| Mylonchulus | 4 | 0.9 ± 1.35 | 0.54 | 0.5 ± 1.43 | 0.32 | 0.1 ± 0.26 | 0.03 | - | - | - | - |
| Oxydirus | 5 | 1.8 ± 1.43 | 1.09 | 2.4 ± 3.38 | 1.62 | 1.4 ± 2.44 | 0.75 | 0.1 ± 0.3 | 0.05 | 0.1 ± 0.72 | 0.1 |
| Thonus | 4 | 0.8 ± 2.06 | 0.49 | 1.5 ± 4.37 | 1.03 | 0.6 ± 1.44 | 0.33 | 1.2 ± 2.94 | 0.89 | - | - |
| Tripyla | 3 | 0.2 ± 0.51 | 0.11 | 5.9 ± 12.54 | 4.01 | 4 ± 8.01 | 2.21 | 11.5 ± 15.07 | 8.83 | 21.6 ± 32.68 | 14.59 |
| Number of genera | 41 | 40 | 35 | 29 | 25 | ||||||
| Index Name | S | S/OMF | S/OMS | OMF | OMS | |
|---|---|---|---|---|---|---|
| A ± SD | A ± SD | A ± SD | A ± SD | A ± SD | p | |
| Abundance, ind. | 169 ± 53.8 a | 146 ± 77.2 a | 180 ± 104 a | 129 ± 34.5 a | 148 ± 84.8 a | - |
| Number of genera | 17.9 ± 3.93 a | 16.1 ± 5.70 a | 14.9 ± 4.89 a | 8.48 ± 1.72 b | 7.68 ± 1.57 b | *** |
| Diversity index for genera | 1.0 ± 0.1 a | 1.0 ± 0.2 a | 0.9 ± 0.2 ab | 0.6 ± 0.1 b | 0.5 ± 0.1 b | *** |
| Total biomass, mg | 0.5 ± 0.4 ab | 0.4 ± 0.3 a | 0.5 ± 0.5 ab | 0.6 ± 0.3 ab | 0.8 ± 0.7 b | * |
| Maturity Index | 2.2 ± 0.3 a | 2.2 ± 0.5 a | 2.1 ± 0.5 a | 1.9 ± 0.7 ab | 1.7 ± 0.8 b | *** |
| Plant Parasitic Index | 2.8 ± 0.2 a | 2.8 ± 0.2 a | 2.8 ± 0.2 a | 2.9 ± 0.6 a | 2.8 ± 0.2 a | - |
| Channel Index | 16.2 ± 14.9 ab | 20.1 ± 14.9 a | 20.2 ± 14.4 a | 4.2 ± 6.5 c | 14.5 ± 22.7 b | *** |
| Basal Index | 15.9 ± 6.1 ab | 15.7 ± 7.1 ab | 18.9 ± 9.7 a | 10.2 ± 9.6 c | 13.8 ± 15.7 bc | *** |
| Enrichment Index | 75.3 ± 9.28 ab | 71.5 ± 13.9 a | 70.7 ± 16.0 a | 85.9 ± 11.3 b | 82.6 ± 17.5 ab | ** |
| Structure Index | 66.8 ± 12.2 a | 65.5 ± 16.8 ab | 51.2 ± 23.5 ab | 50.3 ± 36.8 ab | 42.2 ± 30.9 b | * |
| Herbivores ind. | 59.2 ± 24.5 a | 19.7 ± 20.3 b | 27.4 ± 18.1 ab | 0.4 ± 0.8 c | 0.7 ± 1.7 c | *** |
| Fungivores ind. | 35.9 ± 19.6 a | 35.9 ± 29.1 ab | 52.9 ± 29.2 a | 7.4 ± 8.8 c | 18.3 ± 11.3 bc | *** |
| Bacterivores ind. | 63.5 ± 27.5 a | 73.1 ± 45.8 a | 91.7 ± 71.9 a | 89.6 ± 45.6 a | 105.0 ± 87.8 a | - |
| Predators ind. | 4.0 ± 3.21 a | 9.2 ± 11.7 a | 5.5 ± 7.7 a | 11.8 ± 14.6 a | 22.2 ± 31.6 a | - |
| Omnivores ind. | 7.3 ± 8.0 a | 8.4 ± 9.6 a | 2.5 ± 4.1 b | 20.8 ± 28.2 a | 1.0 ± 1.7 b | *** |
| c-p1 ind. | 34.5 ± 16.9 a | 39.4 ± 33.7 a | 56.1 ± 59.6 a | 62.5 ± 41.1 a | 88.2 ± 90.2 a | - |
| c-p2 ind. | 49.0 ± 23.9 ab | 56.6 ± 40.6 ab | 72.3 ± 43.4 b | 33.3 ± 21.2 a | 35.6 ± 19.9 a | *** |
| c-p3 ind. | 2.1 ± 1.72 a | 7.14 ± 12.2 a | 5.34 ± 8.11 a | 11.9 ± 15.0 a | 21.6 ± 31.9 a | - |
| c-p4 ind. | 23.2 ± 13.9 a | 20.9 ± 15.3 a | 17.4 ± 17.9 a | 21.3 ± 28.1 a | 1.60 ± 1.72 b | *** |
| c-p5 ind. | 1.9 ± 1.4 a | 2.4 ± 3.3 a | 1.4 ± 2.4 abc | 0.1 ± 0.3 c | 0.1 ± 0.7 bc | *** |
| Index Name | S | S/OMF | S/OMS | OMF | OMS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Rows | Columns | Mean | Rows | Columns | Mean | Rows | Columns | Mean | Rows | Columns | Mean | Rows | Columns | |
| Total number, ind. | 169 | 146 | 180 | 130 | 149 | ||||||||||
| V. | 220 | ab (1) | A (2) | 264 | ab | A | 341 | a | A | 101 | b | A | 259 | ab | A |
| VI. | 212 | a | A | 158 | a | AB | 162 | a | AB | 160 | a | A | 165 | a | A |
| VII. | 143 | ab | AB | 149 | ab | AB | 169 | b | AB | 114 | ab | A | 70.5 | a | B |
| VIII. | 154 | a | AB | 72.2 | a | B | 126 | a | B | 132 | a | A | 127 | a | AB |
| IX. | 120 | a | B | 87.6 | a | B | 103 | a | B | 142 | a | A | 120 | a | AB |
| Number of genera | 17.9 | 16.1 | 18.7 | 8.5 | 8.1 | ||||||||||
| V. | 23.2 | ab | A | 25.6 | a | A | 22.0 | b | A | 8.2 | c | A | 8.6 | c | A |
| VI. | 20.0 | a | AB | 16.4 | b | AB | 15.0 | b | AB | 9.4 | c | A | 8.6 | c | A |
| VII. | 17.0 | a | BC | 15.8 | ab | BC | 14.8 | b | B | 8.8 | c | A | 8.6 | c | A |
| VIII. | 15.4 | a | BC | 10.8 | b | C | 10.2 | bc | B | 8.2 | c | A | 7.6 | c | A |
| IX. | 14.2 | a | C | 12.0 | ac | BC | 12.8 | ac | B | 7.8 | bc | A | 7.0 | b | A |
| Diversity index | 1.0 | 1.0 | 0.9 | 0.6 | 0.5 | ||||||||||
| V. | 1.2 | ab | A | 1.2 | b | A | 1.0 | bc | A | 0.6 | ac | A | 0.4 | c | A |
| VI. | 1.1 | a | A | 0.9 | ab | B | 0.9 | ab | A | 0.6 | b | A | 0.4 | b | A |
| VII. | 1.0 | a | AB | 1.0 | ab | AB | 0.9 | ac | A | 0.7 | bc | A | 0.5 | c | A |
| VIII. | 0.9 | a | BC | 0.8 | ab | B | 0.9 | ab | A | 0.5 | b | A | 0.6 | b | A |
| IX. | 0.9 | a | C | 0.9 | a | B | 0.8 | ab | A | 0.6 | ab | A | 0.6 | b | A |
| Total biomass, mg | 0.5 | 0.4 | 0.5 | 0.6 | 0.8 | ||||||||||
| V. | 0.9 | ab | A | 0.7 | ab | AB | 1.4 | ab | A | 0.5 | b | A | 1.8 | a | A |
| VI. | 0.4 | a | AB | 0.7 | ac | B | 0.5 | ab | B | 0.9 | bc | A | 1.1 | c | AB |
| VII. | 0.3 | a | B | 0.2 | a | C | 0.3 | a | BC | 0.4 | a | A | 0.2 | a | C |
| VIII. | 0.4 | a | AB | 0.3 | a | AC | 0.2 | a | BC | 0.6 | a | A | 0.6 | a | BC |
| IX. | 0.3 | ab | B | 0.3 | ab | AC | 0.2 | a | C | 0.6 | b | A | 0.6 | ab | BC |
| Maturity Index | 2.2 | 2.2 | 2.1 | 2.0 | 1.7 | ||||||||||
| V. | 2.3 | ab | A | 2.1 | ac | A | 1.6 | bc | A | 1.3 | bc | AB | 1.2 | b | A |
| VI. | 2.3 | a | A | 1.6 | ab | A | 1.7 | ab | AB | 1.3 | b | B | 1.2 | b | A |
| VII. | 2.3 | a | A | 2.5 | a | B | 2.0 | a | BC | 1.9 | a | A | 1.8 | a | B |
| VIII. | 2.0 | a | A | 2.4 | a | BC | 2.2 | a | C | 2.4 | b | A | 1.9 | a | BC |
| IX. | 2.0 | a | A | 2.6 | ab | C | 2.8 | b | D | 2.8 | b | A | 2.3 | ab | C |
| Plant Parasitic Index | 2.8 | 2.8 | 2.9 | 2.8 | 2.8 | ||||||||||
| V. | 2.6 | a | A | 2.6 | a | A | 2.7 | a | A | 3.0 | a | A | 2.7 | a | A |
| VI. | 2.7 | ab | AC | 2.7 | ab | AB | 2.8 | b | A | 2.0 | ab | A | ND | a | A |
| VII. | 2.9 | a | B | 2.8 | ab | AB | 2.9 | ab | A | ND | a | A | 3.0 | ab | A |
| VIII. | 2.9 | a | BC | 2.9 | a | B | 3.0 | a | A | ND | a | A | ND | a | A |
| IX. | 2.9 | ab | BC | 2.9 | ab | B | 3.0 | b | A | 3.3 | ab | A | ND | a | A |
| Channel Index | 16.2 | 20.1 | 20.2 | 4.2 | 14.5 | ||||||||||
| V. | 20.8 | ab | A | 23.9 | b | A | 11.1 | bc | A | 1.2 | c | A | 2.5 | ac | A |
| VI. | 10.1 | ab | A | 2.9 | ac | B | 20.7 | b | A | 1.2 | c | A | 3.4 | bc | A |
| VII. | 14.2 | a | A | 32.4 | a | A | 21.4 | a | A | 10.5 | a | A | 51.6 | a | B |
| VIII. | 9.1 | a | A | 14.2 | a | A | 15.7 | a | A | 4.3 | a | A | 6.5 | a | A |
| IX. | 26.6 | a | A | 26.9 | a | A | 32.3 | ab | A | 3.9 | b | A | 8.8 | ab | AB |
| Basal Index | 15.9 | 15.7 | 18.9 | 10.2 | 13.8 | ||||||||||
| V. | 16.3 | ab | A | 22.9 | a | A | 13.6 | ab | A | 9.90 | ab | AB | 4.13 | b | A |
| VI. | 17.8 | a | A | 9.61 | ab | B | 19.1 | a | A | 7.58 | ab | AB | 4.44 | b | A |
| VII. | 15.6 | a | A | 22.4 | a | A | 29.7 | a | A | 25.4 | a | B | 39.6 | a | B |
| VIII. | 11.5 | ab | A | 9.94 | ab | B | 18.9 | a | A | 4.19 | b | A | 11.8 | ab | A |
| IX. | 18.7 | a | A | 13.9 | ab | AB | 12.9 | ab | A | 4.00 | b | A | 8.89 | ab | A |
| Enrichment Index | 75.3 | 71.5 | 70.7 | 85.9 | 82.7 | ||||||||||
| V. | 74.9 | ac | A | 67.5 | a | AB | 85.1 | ab | A | 89.8 | bc | A | 95.8 | b | A |
| VI. | 70.1 | a | A | 88.6 | ab | A | 79.1 | ab | A | 92.3 | b | A | 95.5 | b | A |
| VII. | 72.8 | a | A | 57.6 | a | B | 60.4 | a | A | 69.5 | a | B | 57.0 | a | B |
| VIII. | 84.2 | a | A | 80.3 | a | AC | 70.8 | a | A | 87.7 | a | A | 83.4 | a | A |
| IX. | 74.7 | ab | A | 63.4 | ab | BC | 58.2 | a | A | 90.3 | b | A | 81.5 | ab | A |
| Structure Index | 66.8 | 65.5 | 51.2 | 50.3 | 42.2 | ||||||||||
| V. | 69.6 | a | A | 56.6 | ab | A | 37.8 | ab | AB | 9.11 | b | A | 19.9 | b | A |
| VI. | 69.1 | a | A | 53.5 | ab | A | 33.9 | ab | AB | 23.2 | b | AB | 32.5 | ab | A |
| VII. | 70.6 | a | A | 66.4 | a | A | 43.9 | ab | A | 38.9 | ab | BC | 9.69 | b | B |
| VIII. | 70.8 | a | A | 70.4 | a | A | 56.7 | a | A | 86.5 | a | BC | 64.1 | a | AC |
| IX. | 53.8 | a | A | 80.7 | ab | A | 83.8 | ab | B | 93.8 | b | C | 84.9 | ab | C |
| Herbivores ind. | 59.2 | 19.7 | 27.5 | 0.4 | 0.7 | ||||||||||
| V. | 72.7 | a | A | 49.8 | ab | A | 40.3 | ab | A | 1.0 | b | A | 2.8 | b | A |
| VI. | 69.4 | a | A | 9.3 | ab | BC | 20.1 | ab | A | 0.3 | b | A | 0.0 | b | A |
| VII. | 69.6 | a | A | 26.4 | ab | AB | 37.6 | ab | A | 0.0 | b | A | 0.7 | b | A |
| VIII. | 46.4 | a | A | 6.7 | ab | C | 25.8 | ab | A | 0.0 | b | A | 0.0 | b | A |
| IX. | 37.9 | a | A | 6.2 | ab | C | 13.5 | ab | A | 0.8 | b | A | 0.0 | b | A |
| Fungivores ind. | 36.0 | 35.9 | 52.9 | 7.4 | 18.3 | ||||||||||
| V. | 48.3 | ab | A | 77.5 | a | A | 81.6 | a | A | 3.73 | b | A | 19.8 | ab | A |
| VI. | 29.7 | a | A | 14.5 | ab | B | 62.8 | a | A | 4.66 | b | A | 18.6 | ab | A |
| VII. | 24.3 | ab | A | 55.5 | b | A | 37.8 | ab | A | 15.5 | a | A | 29.1 | ab | A |
| VIII. | 39.1 | a | A | 11.1 | ab | B | 35.7 | a | A | 6.51 | b | A | 13.3 | ab | A |
| IX. | 38.6 | a | A | 20.9 | a | B | 46.6 | a | A | 6.78 | a | A | 10.6 | a | A |
| Bacterivores ind. | 63.5 | 73.1 | 91.8 | 89.3 | 106.0 | ||||||||||
| V. | 75.4 | a | AB | 115.0 | ac | A | 207.0 | bc | A | 94.9 | ab | AB | 232.0 | c | A |
| VI. | 93.3 | ab | B | 125 | ab | A | 76.9 | b | B | 152 | b | B | 143 | b | AB |
| VII. | 43.1 | ab | A | 60.9 | ab | AB | 88.9 | b | AB | 81.2 | ab | AB | 38.3 | a | C |
| VIII. | 64.0 | a | AB | 34.7 | a | B | 60.5 | a | B | 69.3 | a | A | 72.5 | a | BC |
| IX. | 41.6 | a | A | 29.2 | a | B | 24.4 | a | C | 48.8 | a | A | 43.2 | a | C |
| Predators ind. | 4.0 | 9.2 | 5.5 | 11.8 | 22.3 | ||||||||||
| V. | 5.6 | ab | AB | 10.4 | a | AC | 3.7 | ab | AB | 0.3 | b | A | 1.4 | ab | A |
| VI. | 7.5 | a | A | 2.2 | ab | B | 1.0 | b | A | 2.5 | ab | AB | 3.7 | ab | AB |
| VII. | 3.4 | a | B | 3. 8 | a | BC | 2.3 | a | AB | 14.5 | a | BC | 2.2 | a | A |
| VIII. | 2.2 | a | B | 7.1 | a | AB | 4.0 | a | AB | 10.5 | a | BC | 38.5 | a | BC |
| IX. | 1.3 | a | B | 22.3 | ab | A | 16.7 | ab | B | 31.3 | b | C | 65.5 | b | C |
| Omnivores ind. | 7.3 | 8.4 | 2.5 | 20.8 | 1.0 | ||||||||||
| V. | 18.1 | a | A | 10.9 | ab | A | 7.9 | ac | A | 0.3 | c | A | 1.1 | bc | A |
| VI. | 12.3 | a | A | 6.8 | ab | A | 0.7 | ab | B | 0.7 | ab | A | 0.0 | b | A |
| VII. | 3.1 | a | B | 2.9 | a | A | 2.2 | a | B | 3.1 | a | A | 0.3 | a | A |
| VIII. | 2.5 | ab | B | 12.6 | ab | A | 0.0 | a | AB | 46.1 | ab | B | 2.4 | b | A |
| IX. | 0.6 | a | B | 9.0 | ab | A | 1.5 | ab | B | 53.9 | b | B | 1.1 | a | A |
| c-p1 ind. | 34.5 | 39.4 | 56.1 | 62.5 | 88.2 | ||||||||||
| V. | 39.9 | a | A | 50.1 | ac | AB | 158.0 | bc | A | 66.9 | ab | AB | 218.0 | b | A |
| VI. | 35.9 | a | A | 91.4 | ab | B | 58.3 | ab | AB | 120 | b | B | 135 | b | AB |
| VII. | 21.1 | a | A | 17.8 | a | AC | 29.1 | a | B | 35.4 | a | A | 15.9 | a | C |
| VIII. | 46.7 | a | A | 23.6 | a | AC | 29.8 | a | B | 54.7 | a | AB | 44.8 | a | BD |
| IX. | 28.8 | a | A | 14.2 | a | C | 5.35 | a | C | 35.4 | a | A | 27.1 | a | CD |
| c-p2 ind. | 49.0 | 56.6 | 72.3 | 33.3 | 35.6 | ||||||||||
| V. | 66.9 | ab | AB | 124.0 | b | A | 121.0 | ab | A | 30.2 | a | A | 33.1 | a | A |
| VI. | 68.4 | a | A | 43.2 | ab | BC | 74.1 | ab | A | 36.9 | ab | AB | 26.3 | b | A |
| VII. | 31.8 | a | B | 67.9 | ab | AB | 85.2 | b | A | 61.3 | ab | A | 51.1 | ab | A |
| VIII. | 33.6 | a | AB | 20.9 | a | C | 49.4 | a | AB | 18.5 | a | AB | 40.7 | a | A |
| IX. | 41.3 | a | AB | 27.2 | a | C | 31.8 | a | B | 16.6 | a | B | 26.7 | a | A |
| c-p3 ind. | 2.1 | 7.14 | 5.34 | 11.9 | 21.6 | ||||||||||
| V. | 2.0 | ab | A | 5.8 | b | AB | 5.4 | ab | AB | 0.0 | a | A | 0.4 | ab | A |
| VI. | 0.6 | a | A | 0.2 | a | A | 0.7 | a | A | 1.3 | a | A | 1.8 | a | A |
| VII. | 3.9 | a | A | 2.1 | a | A | 1.1 | a | A | 14.5 | a | AB | 2.2 | a | A |
| VIII. | 2.8 | a | A | 5.0 | a | AB | 2.6 | a | A | 12.9 | a | AB | 38.5 | a | B |
| IX. | 1.4 | a | A | 22.6 | ab | B | 16.9 | ab | B | 31.2 | b | B | 65.5 | b | B |
| c-p4 ind. | 23.2 | 20.9 | 17.4 | 21.3 | 1.60 | ||||||||||
| V. | 36.6 | a | A | 27.9 | ab | A | 12.8 | ac | A | 0.9 | c | A | 1.4 | bc | A |
| VI. | 34.2 | a | AB | 11.9 | ab | A | 7.9 | ab | A | 1.9 | b | A | 2.1 | b | A |
| VII. | 15.2 | ab | BC | 33.8 | b | A | 14.9 | ab | A | 3.1 | a | A | 0.6 | a | A |
| VIII. | 20.4 | ab | AC | 13.9 | ab | A | 16.7 | ab | A | 46.4 | a | B | 2.7 | b | A |
| IX. | 9.3 | ab | C | 17.2 | ab | A | 34.8 | ab | A | 54.5 | b | B | 1.10 | a | A |
| c-p5 ind. | 1.9 | 2.4 | 1.4 | 0.1 | 0.1 | ||||||||||
| V. | 1.6 | a | A | 6.4 | a | A | 3.7 | a | A | 0.3 | a | A | 0.7 | a | A |
| VI. | 3.3 | a | A | 1.6 | ab | A | 0.3 | ab | A | 0.0 | b | A | 0.0 | b | A |
| VII. | 1.8 | a | A | 1.4 | a | A | 0.9 | a | A | 0.0 | a | A | 0.0 | a | A |
| VIII. | 1.2 | a | A | 2.0 | a | A | 1.6 | a | A | 0.0 | a | A | 0.0 | a | A |
| IX. | 1.3 | a | A | 0.2 | a | A | 0.2 | a | A | 0.0 | a | A | 0.0 | a | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubcsiková, M.; Demková, L.; Renčo, M.; Čerevková, A. Evaluation of the Effect of Organic Matter from Invasive Plants on Soil Nematode Communities. Plants 2023, 12, 3459. https://doi.org/10.3390/plants12193459
Jakubcsiková M, Demková L, Renčo M, Čerevková A. Evaluation of the Effect of Organic Matter from Invasive Plants on Soil Nematode Communities. Plants. 2023; 12(19):3459. https://doi.org/10.3390/plants12193459
Chicago/Turabian StyleJakubcsiková, Michaela, Lenka Demková, Marek Renčo, and Andrea Čerevková. 2023. "Evaluation of the Effect of Organic Matter from Invasive Plants on Soil Nematode Communities" Plants 12, no. 19: 3459. https://doi.org/10.3390/plants12193459