

Essential Oils of Artemisia frigida Plants (Asteraceae): Conservatism and Lability of the Composition

 , , , , and
, , , , and
Abstract
:1. Introduction
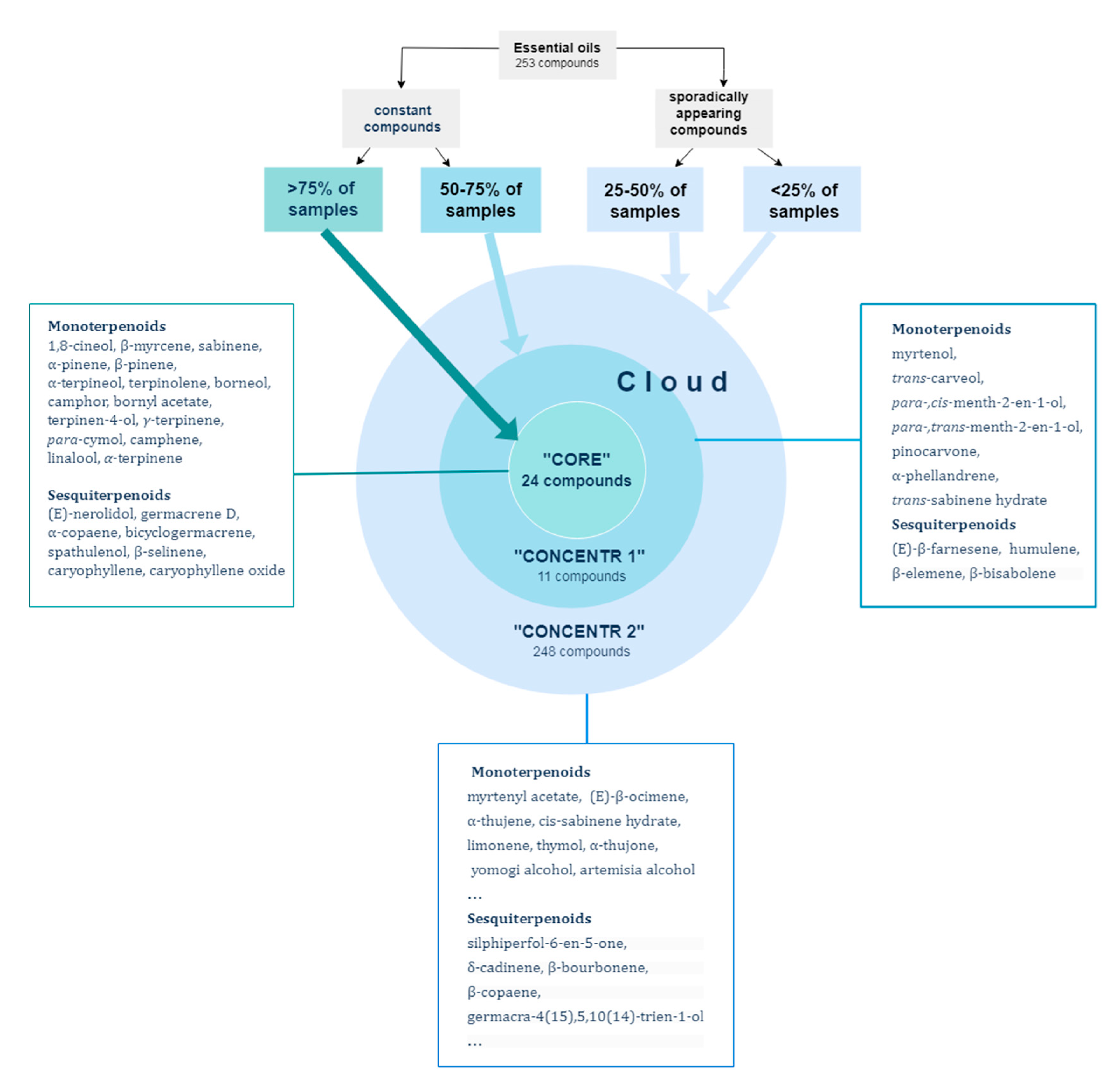
2. Results and Discussion
2.1. Composition of Essential Oils of Artemisia frigida (Based on Plants Growing within the Territory of Buryatia (Russia) and Qinghai (China) from 2016–2021
2.2. Composition of Essential Oils of A. frigida during Ontogenesis and Phenological Development of Plants
2.3. Composition of Essential Oils of A. frigida from the Same Population Depends on the Weather Conditions in Different Years
2.4. Composition of Essential Oils of A. frigida from Different Populations in Buryatia Depends Not Only on Climatic Conditions
2.5. Composition of Essential Oils in Plants of A. frigida Confined to “Typical” and “Atypical” Habitats
2.6. Conservatism s and Lability of the Composition of A. frigida Essential Oils
3. Materials and Methods
3.1. Plant Materials
3.2. Essential Oil Analyses
3.2.1. Isolation of Essential Oils
3.2.2. GC-MS Analysis of Essential Oils
3.2.3. Essential Oil Compound Identification
3.3. Climatic Conditions
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Obyazov, V.A. Izmeneniya Sovremennogo Klimata i ocenka ih Posledstvij dlya Prirodnyh i Prirodno-Klimaticheskih Sistem Zabajkal'ya (Changes in the Modern Climate and Assessment of Their Consequences for the Natural and Climatic Systems of Transbaikalia). Ph.D. Thesis, Kazan State University, Kazan, Russia, 2014; 38p. (In Russian). [Google Scholar]
- Namzalov, B.B. Stepi Yuzhnoj Sibiri (Steppes of Southern Siberia); BSC SB RAS: Novosibirsk–Ulan-Ude, Russia, 1994; 309p. (In Russian) [Google Scholar]
- Zvereva, G.A. Kharakteristike Zharostojkosti Stepnyh i Pustynnyh Rastenij Central'noj Tuvy (To the Characteristics of Heat Resistance of Steppe and Desert Plants of Central Tuva). In Stepnaja Rastitel'nost' Sibiri i Nekotorye Cherty ee Jekologii (Steppe Vegetation of Siberia and some Features of Its Ecology); Gorshkova, A.A., Ed.; Nauka (Sib. Office): Novosibirsk, Russia, 1982; pp. 62–68. (In Russian) [Google Scholar]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Young, E.C.; Veit, D.; Gershenzon, J.; Schuman, M.C. The Sesquiterpenes(E)-β-Farnesene and (E)-α-Bergamotene Quench Ozone but Fail to Protect the Wild Tobacco Nicotiana attenuata from Ozone, UVB, and Drought Stresses. PLoS ONE 2015, 10, e0127296. [Google Scholar] [CrossRef] [PubMed]
- Boncan, D.A.T.; Tsang, S.S.K.; Li, C.; Lee, I.H.T.; Lam, H.-M.; Chan, T.-F.; Hui, J.H.L. Terpenes and Terpenoids in Plants: Interactions with Environment and Insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef] [PubMed]
- Zwenger, S.; Basu, C. Plant terpenoids: Application and future potentials. Biotechnol. Mol. Biol. Rev. 2008, 3, 1–7. [Google Scholar]
- Peñuelas, J.; Staudt, M. BVOCs and global change. Trends Plant Sci. 2010, 15, 133–144. [Google Scholar] [CrossRef]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and sub-cellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Structure and dynamics of the isoprenoid pathway network. Mol. Plant. 2012, 5, 318–333. [Google Scholar] [CrossRef]
- Bolouri, P.; Salami, R.; Kouhi, S.; Kordi, M.; Asgari Lajayer, B.; Hadian, J.; Astatkie, T. Applications of Essential Oils and Plant Extracts in Different Industries. Molecules 2022, 27, 8999. [Google Scholar] [CrossRef]
- Gupta, I.; Singh, R.; Muthusamy, S.; Sharma, M.; Grewal, K.; Singh, H.P.; Batish, D.R. Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints. Plants 2023, 12, 2916. [Google Scholar] [CrossRef]
- Giuffrè, A.M.; Riccardo, N. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emir. J. Food Agric. 2020, 32, 522–532. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, B.; Wani, M.S.; Gupta, R.C.; Habeeb, T.H. Determination of the Volatile Composition in Essential Oil of Azadirachta indica A. Juss from different areas of North Indian Plains by Gas Chromatography/Mass Spectrometry (GC/MS). Anal. Chem. Lett. 2021, 11, 73–82. [Google Scholar] [CrossRef]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The peel essential oil composition of bergamot fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A Review. Emir. J. Food Agric. 2020, 32, 835–845. [Google Scholar] [CrossRef]
- Chatterjee, S.; Gupta, S.; Variyar, S.P. Comparison of Essential Oils Obtained from Different Extraction Techniques as an Aid in Identifying Aroma Significant Compounds of Nutmeg (Myristica fragrans). Nat. Prod. Commun. 2015, 10, 1443–1446. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Jiang, L.; Liu, H.; Song, Z.; Xiang, C. Ontogenetic trait variability and nitrogen stoichiometric homeostasis explained high stability of Artemisia frigida—Dominated grassland. Ecol. Indic. 2022, 145, 109635. [Google Scholar] [CrossRef]
- Deguerry, F.; Pastore, L.; Wu, S.; Clark, A.; Chappell, J.; Schalk, M. The diverse sesquiterpene profile of patchouli, Pogostemon cablin, is correlated with a limited number of sesquiterpene synthases. Arch. Biochem. Biophys. 2006, 454, 123–136. [Google Scholar] [CrossRef]
- Góra, J.; Lis, A.; Kula, J.; Staniszewska, M.; Wołoszyn, A. Chemical composition variability of essential oils in the ontogenesis of some plants. Flavour. Fragr. J. 2002, 17, 445–451. [Google Scholar] [CrossRef]
- Dabibeh, R.M.; Barhoumi, L.M.; Bdair, O.; Al-Jaber, H.L.; Afifi, F.U.; Al-Qudah, M.A.; Abaza, I.F.; Abu-Zarga, M.H.; Abu-Orabi, S.T. Effect of Geographic Location, Ontogenesis on Essential Oil Composition and Spontaneously Emitted Volatile Organic Compounds of Inula viscosa (L.) Greuter. (Astraceae) Grown in Jordan. Asian J. Chem. 2020, 32, 2559–2566. [Google Scholar] [CrossRef]
- Bachheti, A.; Sharma, A.; Bachheti, R.K.; Husen, A.; Pandey, D.P. Plant Allelochemicals and Their Various Applications. In Co-Evolution of Secondary Metabolites, Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; pp. 441–465. [Google Scholar] [CrossRef]
- Wang, Z.; Lv, S.; Han, G.; Wang, Z.; Li, Z.; Ren, H.; Wang, J.; Sun, H.; Zhang, G. Heavy grazing reduced the spatial heterogeneity of Artemisia frigida in desert steppe. BMC Plant Biol. 2022, 22, 337. [Google Scholar] [CrossRef]
- Sarker, L.S.; Galata, M.; Demissie, Z.A.; Mahmoud, S.S. Molecular cloning and functional characterization of borneol dehydrogenase from the glandular trichomes of Lavandula x intermedia. Arch. Biochem. Biophys. 2012, 528, 163–170. [Google Scholar] [CrossRef]
- Korolyuk, E.A.; Pokrovskii, L.M.; Tkachev, A.V. Himicheskij sostav jefirnogo masla predstavitelej roda Gallatella Cass. (Asteraceae Dumont) iz Zapadnoj Sibiri (The chemical composition of essential oil of representatives of the genus Gallatella Cass. (Asteraceae Dumont) from Western Siberia). Khimija Rastitel’nogo Syr’ja 2002, 1, 5–18. (In Russian) [Google Scholar]
- Zhigzhitzhapova, S.V.; Randalova, T.E.; Radnaeva, L.D.; Dylenova, E.P.; Chen, S.; Zhang, F. Chemical composition of essentials oils of Artemisia frigida Willd. (Asteraceae) grown in the North and Central Asia. J. Essent. Oil Bear. Plants 2017, 20, 915–926. [Google Scholar] [CrossRef]
- Moghaddam, H.H.; Jafari, A.A.; Sefidkon, F.; Sepideh, K.J. Influence of climatic factors on essential oil content and composition of 20 populations of Nepeta binaludensis Jamzad from Iran. Appl. Biol. Chem. 2023, 66, 1–10. [Google Scholar] [CrossRef]
- Aćimović, M.; Lončar, B.; Stanković Jeremić, J.; Cvetković, M.; Pezo, L.; Pezo, M.; Todosijević, M.; Tešević, V. Weather Conditions Influence on Lavandin Essential Oil and Hydrolate Quality. Horticulturae 2022, 8, 281. [Google Scholar] [CrossRef]
- Ayse Ozlem Tursun, A.O. Impact of soil types on chemical composition of essential oil of purple basil. Saudi J. Biol. Sci. 2022, 29, 103314. [Google Scholar] [CrossRef]
- Najar, B.; Demasi, S.; Caser, M.; Gaino, W.; Cioni, P.L.; Pistelli, L.; Scariot, V. Cultivation Substrate Composition Influences Morphology, Volatilome and Essential Oil of Lavandula Angustifolia Mill. Agronomy 2019, 9, 411. [Google Scholar] [CrossRef]
- Komariah; Pitaloka, D.D.A.; Batubara, I.; Nurcholis, W.; Sandrawati, A.; Setyawati, A.; Syamsiyah, J.; Dewi, W.S. The effects of soil temperature from soil mulching and harvest age on phenol, flavonoid and antioxidant contents of Java tea (Orthosiphon aristatus B.). Chem. Biol. Technol. Agric. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Bodoev, N.V.; Bazarova, S.V.; Pokrovsky, L.M.; Namzalov, B.B.; Tkachev, A.V. Himicheskij sostav jefirnogo masla polyni holodnoj (artemisia frigida willd.), proizrastajushhej v zabajkal''e (Chemical composition of the essential oil of Artemisia frigida Willd., growing in Transbaikal). Khimija Rastitel’nogo Syr’ja 2000, 3, 41–44. (In Russian) [Google Scholar]
- Zhigzhitzhapova, S.V.; Radnaeva, L.D.; Chimitcyrenova, L.I. Komponentnyj sostav jefirnogo masla polyni holodnoj flory burjatii (Component composition of essential oils Artemisia frigida Willd. from Buryatia). Bull. Buryat State Univ. Med. Pharm. 2013, 12, 71–74. (In Russian) [Google Scholar]
- Zhigzhitzhapova, S.V.; Randalova, T.E.; Radnaeva, L.D. Sostav jefirnyh masel Artemisia frigida Willd., proizrastajushhih na territorii vodosbornogo bassejna ozera Bajkal (The composition of the essential oils of Artemisia frigida Willd., growing in the watershed of Lake Baikal). Khimija Rastitel’nogo Syr’ja 2014, 3, 151–158. (In Russian) [Google Scholar]
- Pushkareva, E.S.; Efremov, E.A. Component composition of the essential oil of Artemisia frigida from Krasnoyarsk region and of its individual fractions. Sorbtsionnye Khromatograficheskie Protsessy 2012, 12, 619–623. (In Russian) [Google Scholar]
- Korolyuk, E.A.; Tkachev, A.V. Chemical composition of the essential oil from two wormwood species Artemisia frigida and Artemisia argyrophylla. Russ. J. Bioorg. Chem. 2010, 36, 884–893. [Google Scholar] [CrossRef]
- Tkachev, A.V.; Prokusheva, D.L.; Domrachev, D.V. Dikorastushhie Jefirnomas-Lichnye Rastenija Juzhnoj Sibiri (Wild Growing Aromatic Plants of the Southern Siberia); Offset-TM Publishing Hause: Novosibirsk, Russia, 2017; 575p, ISBN 978-5-85957-139-0. (In Russian) [Google Scholar]
- Atazhanova, G.A.; Dembitsky, A.D.; Yakovleva, T.D.; Ishmuratova, M.Y.; Mikhailov, V.G.; Adykenov, S.M. Composition of essential oils Artemisia radicans and A. frigida. Chem. Nat. Comp. 1999, 35, 427–429. [Google Scholar] [CrossRef]
- Jambal, I.; Ayurzana, A.; Shatar, A. Essential oil composition and antioxidant, anticancer activities of Artemisia frigida Willd. grown in Mongolia. J. Agric. Res. 2023, 9, 1–7. [Google Scholar] [CrossRef]
- Lopes-Lutz, D.; Alviano, D.S.; Alviano, C.S.; Kolodziejczyk, P.P. Screening of chemical composition, antimicrobial and antioxidant activities of Artemisia essential oils. Phytochemistry 2008, 69, 1732–1738. [Google Scholar] [CrossRef]
- Liu, X.C.; Li, Y.; Wang, T.; Wang, Q.; Liu, Z.L. Chemical composition and insecticidal activity of essential oil of Artemisia frigida Willd (Compositae) against two grain storage insects. Trop. J. Pharm. Res. 2014, 13, 587–592. [Google Scholar] [CrossRef]
- Zhigzhitzhapova, S.V.; Radnaeva, L.D.; Gao, Q.B.; Chen, S.L.; Zhang, F.Q. Chemical composition of volatile organic compounds of Artemisia vulgaris L. (Asteraceae) from the Qinghai-Tibet Plateau. Ind. Crops Prod. 2016, 83, 462–469. [Google Scholar] [CrossRef]
- Zhigzhitzhapova, S.V.; Dylenova, E.P.; Gulyaev, S.M.; Randalova, T.E.; Taraskin, V.V.; Tykheev, Z.A.; Radnaeva, L.D. Composition and antioxidant activity of the essential oil of Artemisia annua L. Nat. Prod. Res. 2020, 34, 2668–2671. [Google Scholar] [CrossRef]
- Dylenova, E.P.; Zhigzhitzhapova, S.V.; Zhigzhitzhapov, B.V.; Zhigmittsyrenova, B.M.; Taraskina, A.S.; Anenkhonov, O.A. Influence of climatic comditions on the composition of Artemisia jacutica (Asteraceae) essential oil. Contemp. Probl. Ecol. 2023, 16, 357–371. [Google Scholar] [CrossRef]
- Miroshnichenko, Y.M. O rasprostranenii Artemisia frigida Willd. v MNR (On the distribution of Artemisia frigida Willd. in the Mongolian People’s Republic). Bot. Zhurnal 1965, 50, 420–424. (In Russian) [Google Scholar]
- Korolyuk, A.Y. Podhody k analizu struktury rastitel'nogo pokrova landshaftov s peresechennym rel'efom (Analysis of spatial patterns in vegetation of landscapes with complicated relief). Izv. Samar. Nauchnogo Cent. Ross. Akad. Nauk. 2012, 14, 1280–1283. (In Russian) [Google Scholar]
- Anenkhonov, O.A.; Sandanov, D.V.; Naidanov, B.B.; Chimitov, D.G.; Liu, H.; Xu, C.; Guo, W.; Korolyuk, A.Y.; Zvereva, A.A. Using data on the thermal conditions of soils for the differentiation of vegetation in the exposure-related forest-steppe of Transbaikalia. Contemp. Probl. Ecol. 2020, 13, 522–532. [Google Scholar] [CrossRef]
- Liu, H.; He, S.; Anenkhonov, O.A.; Hu, G.; Sandanov, D.V.; Badmaeva, N.K. Topography-Controlled Soil Water Content and the Coexistence of Forest and Steppe in Northern China. Phys. Geogr. 2012, 33, 561–573. [Google Scholar] [CrossRef]
- Namzalov, B.B. Extrazonal steppe phenomena in the mountains of Southern Siberia: Features of spatial organization and centers of the latest speciation and cenogenesis. Contemp. Probl. Ecol. 2020, 13, 495–504. [Google Scholar] [CrossRef]
- Fähnrich, A.; Brosemann, A.; Teske, L.; Neumann, M.; Piechulla, B. Synthesis of ‘cineole cassette’ monoterpenes in Nicotiana section Alatae: Gene isolation, expression, functional characterization and phylogenetic analysis. Plant Mol. Biol. 2012, 79, 537–553. [Google Scholar] [CrossRef]
- Chen, F.; Ro, D.-K.; Petri, J.; Gershenzon, J.; Bohlmann, J.; Pichersky, E.; Tholl, D. Characterization of a root-specific Arabidopsis terpene synthase responsible for the formation of the volatile monoterpene 1,8-cineole. Plant Physiol. 2004, 135, 1956–1966. [Google Scholar] [CrossRef]
- Chen, F.; Rholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthase in plants: A mid-sixe family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Fähnrich, A.; Krause, K.; Piechulla, B. Product Variability of the ‘Cineole Cassette’ Monoterpene Synthases of Related Nicotiana Species. Mol. Plant 2011, 4, 965–984. [Google Scholar] [CrossRef]
- Lima, A.S.; Schimmel, J.; Lukas, B.; Novak, J.; Barroso, J.G.; Figueiredo, A.C.; Pedro, L.G.; Degenhardt, J.; Trindade, H. Genomic characterization, molecular cloning and expression analysis of two terpene synthases from Thymus caespititius (Lamiaceae). Planta 2013, 238, 191–204. [Google Scholar] [CrossRef]
- Krause, S.T.; Liao, P.; Crocoll, C.; Boachon, B.; Förster, C.; Leidecker, F.; Wiese, N.; Zhao, D.; Wood, J.C.; Buell, C.R.; et al. The biosynthesis of thymol, carvacrol, and thymohydroquinone in Lamiaceae proceeds via cytochrome P450s and a short-chain dehydrogenase. Proc. Natl. Acad. Sci. USA 2021, 118, e2110092118. [Google Scholar] [CrossRef]
- Sallaud, C.; Rontein, D.; Onillon, S.; Jabès, F.; Duffé, P.; Giacalone, C.; Thoraval, S.; Escoffier, C.; Herbette, G.; Leonhardt, N.; et al. A novel pathway for sesquiterpene biosynthesis from Z,Z-farnesyl pyrophosphate in the wild tomato Solanum habrochaites. Plant Cell 2009, 21, 301–317. [Google Scholar] [CrossRef]
- Semenov, A.A.; Karcev, V.G. Osnovy Himii Prirodnyh Soedinenij (Outline of the Chemistry of Natural Compounds); ICSPF: Moscow, Russia, 2009; Volume 1, 624p, ISBN 978-5-903078-12-7. (In Russian) [Google Scholar]
- Bülow, N.; König, W.A. The role of germacrene D as a precursor in sesquiterpene biosynthesis: Investigations of acid catalyzed, photochemically and thermally induced rearrangements. Phytochemistry 2000, 55, 141–168. [Google Scholar] [CrossRef]
- Tkachev, A.V. Issledovanie Letuchih Veshhestv Rastenij (The Study of Plant Volotiles); Offset-TM Publishing House: Novosibirsk, Russia, 2008; 969p, ISBN 978-5-85957-056-0. (In Russian) [Google Scholar]
- Eddin, L.B.; Jha, N.K.; Goyal, S.N.; Agrawal, Y.O.; Subramanya, S.B.; Bastaki, S.M.A.; Ojha, S. Health Benefits, Pharmacological Effects, Molecular Mechanisms, and Therapeutic Potential of α-Bisabolol. Nutrients 2022, 14, 1370. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, F.; Cellini, A.; Marchetti, L.; Mudigere, K.; Piovene, C. Emission and Function of Volatile Organic Compounds in Response to Abiotic Stress. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; InTech: London, UK, 2011. [Google Scholar] [CrossRef]
- Bertamini, M.; Grando, M.S.; Zocca, P.; Pedrotti, M.; Lorenzi, S.; Cappelin, L. Linking monoterpenes and abiotic stress resistance in grapevines. BIO Web Conf. 2019, 13, 01003. [Google Scholar] [CrossRef]
- Fontana, A.; Held, M.; Fantaye, C.A.; Turlings, T.C.; Degenhardt, J.; Gershenzon, J. Attractiveness of Constitutive and Herbivore-Induced Sesquiterpene Blends of Maize to the Parasitic Wasp Cotesia marginiventris (Cresson). J. Chem. Ecol. 2011, 37, 582–591. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Nicolia, A.; Diretto, G. Terpenoid Transport in Plants: How Far from the Final Picture? Plants 2023, 12, 634. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dong, W.; Li, H.; D’Onofrio, C.; Bai, P.; Chen, R.; Yang, L.; Wu, J.; Wang, X.; Wang, B.; et al. Molecular basis of (E)-β-farnesene-mediated aphid location in the predator Eupeodes corolla. Curr. Biol. 2022, 32, 951–962.e7. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and Biological Functions of Terpenoids in Plants. In Biotechnology of Isoprenoids. Advances in Biochemical Engineering/Biotechnology; Schrader, J., Bohlmann, J., Eds.; Springer: Cham, Switzerland, 2015; pp. 63–106. [Google Scholar] [CrossRef]
- Blagojević, P.; Radulović, N.; Palić, R.; Stojanović, G. Chemical composition of the essential oil of Serbian wild-growing Artemisia absinthium and Artemisia vulgaris. J. Agric. Food Chem. 2006, 54, 4780–4789. [Google Scholar] [CrossRef]
- Song, C.; Ma, J.; Li, G.; Pan, H.; Zhu, Y.; Jin, Q.; Cai, Y.; Han, B. Natural Composition and Biosynthetic Pathways of Alkaloids in Medicinal Dendrobium Species. Front. Plant Sci. 2022, 13, 850949. [Google Scholar] [CrossRef]
- Gelman, A.; Hill, J.; Vehtary, A. Regression and Other Stories; Cambridge University Press: Cambridge, UK, 2020; 548p, ISBN 9781107023987. [Google Scholar]
- Pena, E.A.; Slate, E.H. Global Validation of Linear Model Assumptions. J. Am. Stat. Assoc. 2006, 101, 341. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and Anti-Inflammatory Activities of Essential Oils: A Short Review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RI * | Component | Peak Area (%) | RI * | Component | Peak Area (%) |
|---|---|---|---|---|---|
| 932 | pinene, α- | 0.77 | 1162 | pinocarvone | 0.24 |
| 947 | camphene | 1.44 | 1166 | borneol | 22.46 |
| 975 | pinene, β- | 0.38 | 1177 | terpinen-4-ol | 4.79 |
| 991 | myrcene, β- | 1.13 | 1191 | terpineol, α | 1.81 |
| 1017 | terpinene, α- | 0.73 | 1287 | bornyl acetate | 3.57 |
| 1024 | cymol, para- | 0.63 | 1378 | copaene, α- | 0.22 |
| 1028 | limonene | 0.05 | 1387 | bourbonene, β- | 0.13 |
| 1031 | cineol, 1,8- | 25.11 | 1392 | elemene, β- | 0.10 |
| 1058 | terpinene, γ- | 1.33 | 1422 | caryophyllene | 0.36 |
| 1066 | sabinene hydrate, trans- | 1.44 | 1484 | germacrene D | 0.48 |
| 1088 | terpinolene | 0.28 | 1488 | selinene, β- | 1.18 |
| 1098 | sabinene hydrate, cis- | 1.08 | 1527 | cadinene, δ- | 0.23 |
| 1100 | libalool | 0.88 | 1580 | spathulenol | 0.76 |
| 1121 | menth-2-en-1-ol., para-, cis- | 0.43 | 1586 | caryophyllene oxide | 0.56 |
| 1144 | camphor | 22.58 | ∑monoterpenes | 91.33 | |
| ∑sesquiterpenes | 4.02 | ||||
| x | a | Y | b | c | z | p-Value | R2 (adjR2) |
|---|---|---|---|---|---|---|---|
| model z = a*x + b | |||||||
| Kextr.year | 30.13 | 0.10 | trans-carveol | 0.001 | 0.62 | ||
| model z = a*x + b*y + c | |||||||
| ∑RJune–August | −0.002 | ∑tveg. | −3.89 × 10−4 | 1.17 | menth-2-en-1-ol, para-, trans- | 6.70 × 10−3 | 0.52 |
| ΣtJune–August | 0.08 | ∑RMay–June | 0.77 | −155.63 | camphor | 7.30 × 10−4 | 0.68 |
| ∑RJune–August | 0.03 | ΣtJune–August | 1.14 × 10−2 | −18.32 | bornyl acetate | 2.91 × 10−3 | 0.59 |
| Year of Collection | Meteorological Station | Σt | R | HTC |
|---|---|---|---|---|
| Western Siberia | ||||
| 1999 | 36038 Zmeinogorsk | 2208.6 | 157.1 | 0.7 |
| 36229 Ust-Kosa | 1818.8 | 279.5 | 1.5 | |
| 2000 | 29915 Slavgorod | 2178.9 | 210.1 | 1.0 |
| 2001 | 36259 Kosh-Agach | 1532.1 | 68.2 | 0.4 |
| 2003 | 360555 Kyzyl-Ozyok | 1951.1 | 268.0 | 1.4 |
| 2003 | 36045 Soloneshnoye | 2116.7 | 258.7 | 1.2 |
| 2005 | 29915 Slavgorod | 2297.1 | 123.8 | 0.5 |
| 2007 | 36177 Semey | 2341.9 | 181.3 | 0.8 |
| 2006 | 36096 Kyzyl | 1884.7 | 193.7 | 1.0 |
| Eastern Siberia | ||||
| 2006 | 36307 Erzin | 1829.8 | 96.7 | 0.5 |
| 2006 | 29862 Abakan | 1836.5 | 212.3 | 1.2 |
| 1999 | 30636 Barguzin | 1890.2 | 238.5 | 1.3 |
| 2014 | 30859 Aginskoye | 1954.6 | 96.6 | 0.5 |
| 2014 | 30825 Ivolginsk | 1881.0 | 97.9 | 0.5 |
| 2015 | 30745 Sosnovo-Ozerskoye | 1708.3 | 175.9 | 1.0 |
| 2017 | 30632 Bolshoy Ushkany Island | 1325.1 | 87.2 | 0.7 |
| 2017 | 30925 Kyakhta | 2150.6 | 232.5 | 1.0 |
| 2017 | 30823 Ulan-Ude | 2116.2 | 62.8 | 0.3 |
| 2021 | 30823 Ulan-Ude | 1874.5 | 258.8 | 1.4 |
| Sample Code | Collection Date | Location | Latitude Longitude | Altitude (m) | Yield V/M (%) |
|---|---|---|---|---|---|
| BURi 12.08.2016 | 12 August 2016 | Russia, Buryatia, Ivolginsky district, steppe | N 51°42 E 107°12 | 645 | 0.7 |
| BURi 01.07.2017 | 1 July 2017 | 0.6 | |||
| BURi 30.08.2017 | 30 August 2017 | 0.7 | |||
| BURi 22.08.2018 | 22 August 2018 | 1.0 | |||
| BURi 08.08.2020 | 8 August 2020 | 1.0 | |||
| BURi 26.08.2020 | 26 August 2020 | 0.7 | |||
| BURi 27.07.2021 | 27 July 2021 | 0.3 | |||
| BURi 20.08.2021 | 20 August 2021 | 0.3 | |||
| BURi 17.09.2021 | 17 September 2021 | 0.3 | |||
| BURs 24.08.2016 | 24 August 2016 | Russia, Buryatia, Selenginsky district, steppe | N 51°22 E 106°32 | 618 | 0.3 |
| BURs 30.08.2017 | 30 August 2017 | 0.9 | |||
| BURe 24.08.2017 | 24 August 2017 | Russia, Buryatia, Eravninsky district, steppe | N 47°47 E 107°13 | 926 | 0.3 |
| BURe 25.08.2016 | 25 August 2016 | 0.3 | |||
| BURroad 19.08.2021 | 19 August 2021 | Russia, Buryatia, Ivolginsky district, road | N 51°51 E 105°30 | 506 | 0.3 |
| BURroad 17.09.2021 | 17 September 2021 | 0.3 | |||
| BURhill 27.07.2021 | 27 July 2021 | Russia, Buryatia, Ivolginsky district, the hill top | N 51°51 E 105°30 | 549 | 0.3 |
| BURhill 19.08.2021 | 19 August 2021 | 0.3 | |||
| BUR 1343 m 17.08.2016 | 17 August 2016 | Russia, Buryatia, Okinsky district, steppe | N 33°21 E 96°58 | 1343 | 0.3 |
| BURss 24.08.2016 | 24 August 2016 | Russia, Buryatia, Selenginsky district, southern slope | N 51°26 E 106°34 | 689 | 0.3 |
| BUR 27.08.2020 | 27 August 2020 | Russia, Buryatia, Khorinsky district, steppe | N 52°13 E 109°57 | 772 | 0.7 |
| BUR 08.07.2017 | 8 July 2017 | Russia, Buryatia, Zaigraevsky district, steppe | N 52°27 E 108°71 | 575 | 0.3 |
| BUR 31.08.2017 | 31 August 2017 | Russia, Buryatia, Selenginsky district, steppe | N 51°26 E 106°34 | 0.3 | |
| BUR 12.08.2017 | 2 August 2017 | Russia, Buryatia, Zaigraevsky district, steppe | N 51°42 E 107°12 | 572 | 0.7 |
| OLNp 05.08.2017 | 5 August 2017 | Russia, Irkytskaya oblast, Olkhon district, steppe | N 53°02 E 106°93 | 469 | 0.9 |
| OLNs 05.08.2017 | 5 August 2017 | Russia, Irkutskaya oblast, Olkhon district, vicinity of the Sokhor cove, steppe | N 53°02 E 106°76 | 491 | 1.0 |
| OLNsh 05.08.2021 | 5 August 2021 | Russia, Irkutskaya oblast, Olkhon district, vicinity of the Aya cove, steppe | N 52°79 E 106°60 | 780 | 0.7 |
| CHN, 2016 4333 m | 15 July 2016 | China, Qinghai Province | N 33°08 E 96°43 | 4333 | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhigzhitzhapova, S.V.; Dylenova, E.P.; Zhigzhitzhapov, B.V.; Goncharova, D.B.; Tykheev, Z.A.; Taraskin, V.V.; Anenkhonov, O.A. Essential Oils of Artemisia frigida Plants (Asteraceae): Conservatism and Lability of the Composition. Plants 2023, 12, 3422. https://doi.org/10.3390/plants12193422
Zhigzhitzhapova SV, Dylenova EP, Zhigzhitzhapov BV, Goncharova DB, Tykheev ZA, Taraskin VV, Anenkhonov OA. Essential Oils of Artemisia frigida Plants (Asteraceae): Conservatism and Lability of the Composition. Plants. 2023; 12(19):3422. https://doi.org/10.3390/plants12193422
Chicago/Turabian StyleZhigzhitzhapova, Svetlana V., Elena P. Dylenova, Bato V. Zhigzhitzhapov, Danaya B. Goncharova, Zhargal A. Tykheev, Vasiliy V. Taraskin, and Oleg A. Anenkhonov. 2023. "Essential Oils of Artemisia frigida Plants (Asteraceae): Conservatism and Lability of the Composition" Plants 12, no. 19: 3422. https://doi.org/10.3390/plants12193422