
A Novel Nutrient- and Antioxidant-Based Formulation Can Sustain Tomato Production under Full Watering and Drought Stress in Saline Soil
 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Growth and Yield Traits, and Water Use Efficiency
2.2. Tomato Fruit Quality Traits
2.3. Leaf Photosynthetic Pigments
2.4. Leaf Macro Mineral Content
2.5. Leaf Micro Mineral Contents
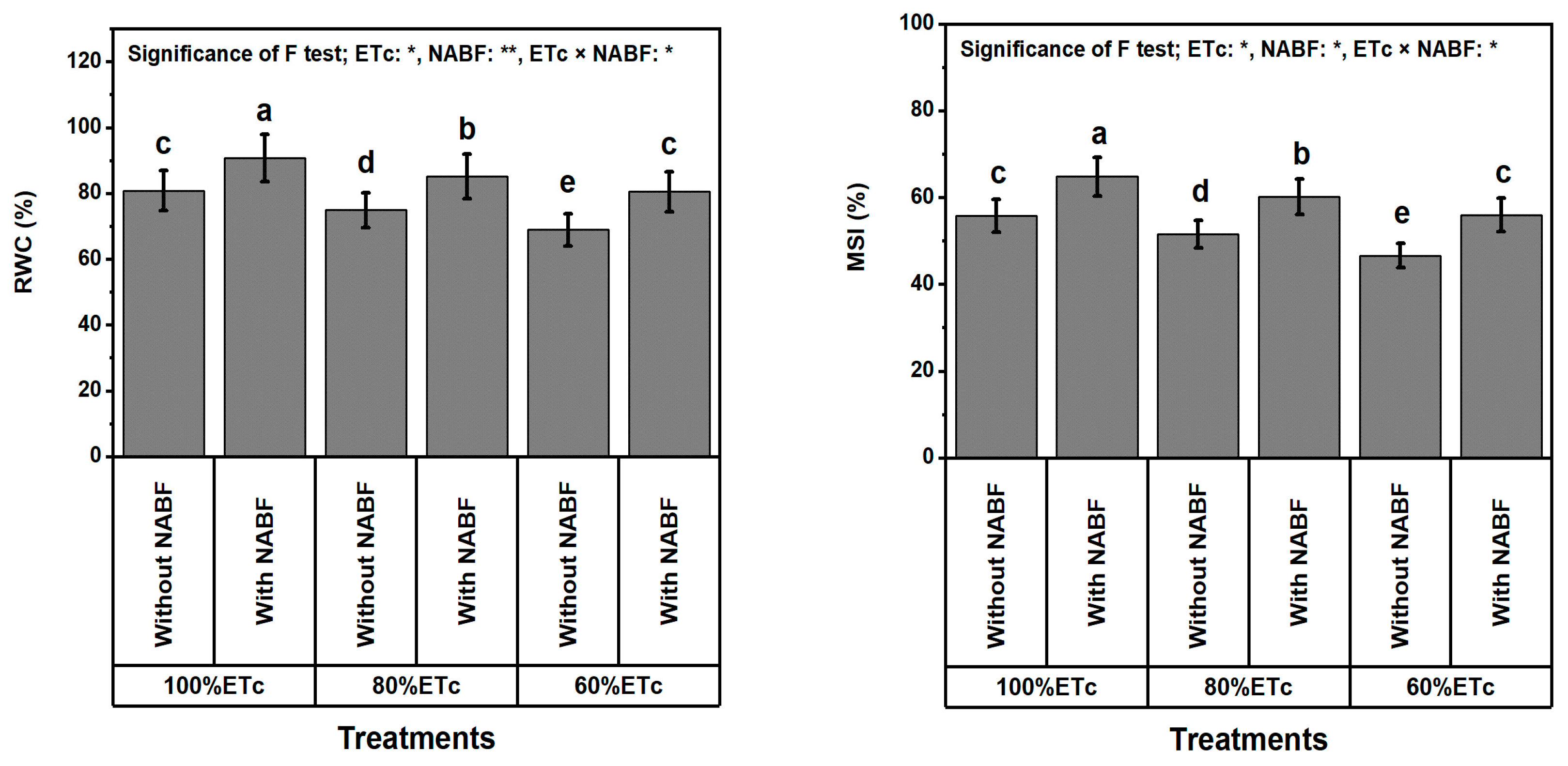
2.6. Integrity of Leaves
2.7. Ionic Leakage, Lipid Peroxidation, and Oxidative Stress Markers
2.8. Osmoprotectants (OPs) and Antioxidant Compounds
2.9. Enzymatic Antioxidant Activities and Soluble Protein Content (SPC)
2.10. Levels of Transcription of Antioxidant Enzyme-Encoding Genes of Tomato Plants
2.11. Phytohormone Levels
3. Discussion
4. Materials and Methods
4.1. Trial Location and Soil Analysis
4.2. Transplanting, Experimental Treatments, and Layout
4.3. Fertilization Program
4.4. Irrigation Water Applied (IWA)
4.5. Plant Sampling
4.6. Fruit Yield Characteristics, and Fruit Quality Traits
4.7. Assessment of Efficiency of Photosynthetic Machinery
4.8. Nutrient Content Assessments
4.9. Assessment of Leaf Tissue Integrity and Oxidative Stress Markers
4.10. Assessments of Osmoprotectant (OP) and Antioxidant Levels
4.11. Assay of Antioxidant Enzyme Activities and Enzyme Gene Expression
4.12. Determination of Phytohormone Contents
4.13. Statistical Analysis Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knapp, S. Tobacco to Tomatoes: A Phylogenetic Perspective on Fruit Diversity in the Solanaceae. J. Exp. Bot. 2002, 53, 2001–2022. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Zehra, A.; Swapnil, P.; Dubey, M.K.; Patel, C.B.; Upadhyay, R.S. Effect on Lycopene, β-Carotene, Ascorbic Acid and Phenolic Content in Tomato Fruits Infected by Alternaria alternata and Its Toxins (TeA, AOH and AME). Arch. Phytopathol. Plant Prot. 2017, 50, 317–329. [Google Scholar] [CrossRef]
- Dorais, M.; Ehret, D.L.; Papadopoulos, A.P. Tomato (Solanum lycopersicum) Health Components: From the Seed to the Consumer. Phytochem. Rev. 2008, 7, 231–250. [Google Scholar] [CrossRef]
- US Department of Agriculture, Agricultural Research Service, Nutrient Data Laboratory. USDA National Nutrient Database for Standard Reference, Release 28 (Slightly Revised). Version Current: May 2016. Available online: http://www.ars.usda.gov/ba/bhnrc/ndl (accessed on 20 April 2023).
- FAO Food and Agriculture Organization of the United Nations Statistics Division. 2014. Available online: http://Faostat3.Fao.Org/Browse/Q/QC/E (accessed on 20 April 2023).
- Battilani, A.; Prieto, M.H.; Argerich, C.; Campillo, C.; Cantore, V. Tomato. In Crop Yield Response to Water, Irrigation and Drainage Paper No. 66.; Steduto, P., Hsiao, T.C., Fereres, E., Raes, D., Eds.; FAO: Rome, Italy, 2012; pp. 192–198. ISBN 978-92-5-107274-5. [Google Scholar]
- Cantore, V.; Wassar, F.; Yamaç, S.S.; Sellami, M.H.; Albrizio, R.; Stellacci, A.M.; Todorovic, M. Yield and Water Use Efficiency of Early Potato Grown under Different Irrigation Regimes. Int. J. Plant Prod. 2014, 8, 409–428. [Google Scholar]
- Manners, R.; Varela-Ortega, C.; van Etten, J. Protein-Rich Legume and Pseudo-Cereal Crop Suitability under Present and Future European Climates. Eur. J. Agron. 2020, 113, 125974. [Google Scholar] [CrossRef]
- Abdelhafez, A.A.; Metwalley, S.M.; Abbas, H. Irrigation: Water Resources, Types and Common Problems in Egypt. In Technological and Modern Irrigation Environment in Egypt; Springer: Chem, Switzerland, 2020; pp. 15–34. [Google Scholar]
- Abd El Mageed, T.A.; Semida, W.; Hemida, K.A.; Gyushi, M.A.H.; Rady, M.M.; Abdelkhalik, A.; Merah, O.; Brestic, M.; Mohamed, H.I.; El Sabagh, A.; et al. Glutathione-Mediated Changes in Productivity, Photosynthetic Efficiency, Osmolytes, and Antioxidant Capacity of Common Beans (Phaseolus vulgaris) Grown under Water Deficit. PeerJ 2023, 11, e15343. [Google Scholar] [CrossRef]
- Ramadan, K.M.A.; El-Beltagi, H.S.; El-Mageed, T.A.A.; Saudy, H.S.; Al-Otaibi, H.H.; Mahmoud, M.A.A. The Changes in Various Physio-Biochemical Parameters and Yield Traits of Faba Bean Due to Humic Acid Plus 6-Benzylaminopurine Application under Deficit Irrigation. Agronomy 2023, 13, 1227. [Google Scholar] [CrossRef]
- Pereira, L.S.; Oweis, T.; Zairi, A. Irrigation Management under Water Scarcity. Agric. Water Manag. 2002, 57, 175–206. [Google Scholar] [CrossRef]
- Kirda, C.; Cetin, M.; Dasgan, Y.; Topcu, S.; Kaman, H.; Ekici, B.; Derici, M.R.; Ozguven, A.I. Yield Response of Greenhouse Grown Tomato to Partial Root Drying and Conventional Deficit Irrigation. Agric. Water Manag. 2004, 69, 191–201. [Google Scholar] [CrossRef]
- Shaaban, A.; Al-Elwany, O.A.A.I.; Abdou, N.M.; Hemida, K.A.; El-Sherif, A.M.A.; Abdel-Razek, M.A.; Semida, W.M.; Mohamed, G.F.; Abd El-Mageed, T.A. Filter Mud Enhanced Yield and Soil Properties of Water-Stressed Lupinus termis L. in Saline Calcareous Soil. J. Soil Sci. Plant Nutr. 2022, 22, 1572–1588. [Google Scholar] [CrossRef]
- Azab, E.S.; Alshallash, K.S.; Alqahtani, M.M.; Safhi, F.A.; ALshamrani, S.M.; Ali, M.A.M.; El-Mageed, T.A.A.; El-Taher, A.M. Physiological, Anatomical, and Agronomic Responses of Cucurbita Pepo to Exogenously Sprayed Potassium Silicate at Different Concentrations under Varying Water Regimes. Agronomy 2022, 12, 2155. [Google Scholar] [CrossRef]
- Dorji, K.; Behboudian, M.H.; Zegbe-Domínguez, J.A. Water Relations, Growth, Yield, and Fruit Quality of Hot Pepper under Deficit Irrigation and Partial Rootzone Drying. Sci. Hortic. 2005, 104, 137–149. [Google Scholar] [CrossRef]
- Cantore, V.; Lechkar, O.; Karabulut, E.; Sellami, M.H.; Albrizio, R.; Boari, F.; Stellacci, A.M.; Todorovic, M. Combined Effect of Deficit Irrigation and Strobilurin Application on Yield, Fruit Quality and Water Use Efficiency of “Cherry” Tomato (Solanum lycopersicum L.). Agric. Water Manag. 2016, 167, 53–61. [Google Scholar] [CrossRef]
- Agami, R.A.; Alamri, S.A.M.; Abd El-Mageed, T.A.; Hashem, M.S.M.; Mohamed, A. Role of Exogenous Nitrogen Supply in Alleviating the Deficit Irrigation Stress in Wheat Plants. Agric. Water Manag. 2018, 210, 261–270. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Gyushi, M.A.H.; Hemida, K.A.; El-Saadony, M.T.; Abd El-Mageed, S.A.; Abdalla, H.; AbuQamar, S.F.; El-Tarabily, K.A.; Abdelkhalik, A. Coapplication of Effective Microorganisms and Nanomagnesium Boosts the Agronomic, Physio-Biochemical, Osmolytes, and Antioxidants Defenses Against Salt Stress in Ipomoea batatas. Front. Plant Sci. 2022, 13, 883274. [Google Scholar] [CrossRef]
- Navarro-León, E.; López-Moreno, F.J.; Fernández, M.A.; Maldonado, J.J.; Yánez, J.; Blasco, B.; Ruiz, J.M. A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants. Agronomy 2022, 12, 410. [Google Scholar] [CrossRef]
- Al-Elwany, O.A.A.I.; Mohamed, G.F.; Abdurrahman, H.A.; Rady, M.M.; Latef, A.A.A. Exogenous Glutathione-Mediated Tolerance to Deficit Irrigation in Salt-Affected Capsicum frutescence (L.) Plants Is Connected with Higher Antioxidant Content and Ionic Homeostasis. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1957–1979. [Google Scholar] [CrossRef]
- Geng, G.; Cakmak, I.; Ren, T.; Lu, Z.; Lu, J. Effect of Magnesium Fertilization on Seed Yield, Seed Quality, Carbon Assimilation and Nutrient Uptake of Rapeseed Plants. Field Crop. Res. 2021, 264, 108082. [Google Scholar] [CrossRef]
- Bhadwal, S.; Sharma, S. Selenium Alleviates Physiological Traits, Nutrient Uptake and Nitrogen Metabolism in Rice under Arsenate Stress. Environ. Sci. Pollut. Res. 2022, 29, 70862–70881. [Google Scholar] [CrossRef]
- Blasco, B.; Ríos, J.J.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Leyva, R.; Romero, L.; Ruiz, J.M. Study of the Interactions Between Iodine and Mineral Nutrients in Lettuce Plants. J. Plant Nutr. 2012, 35, 1958–1969. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Chen, Q.; Feng, H.; Hao, M.; Niu, G.; Liu, Y.; Li, C. Interactive Effect of Irrigation and Blend Ratio of Controlled Release Potassium Chloride and Potassium Chloride on Greenhouse Tomato Production in the Yellow River Basin of China. Agric. Water Manag. 2022, 261, 107346. [Google Scholar] [CrossRef]
- Kim, Y.X.; Kwon, M.C.; Lee, S.; Jung, E.S.; Lee, C.H.; Sung, J. Effects of Nutrient and Water Supply During Fruit Development on Metabolite Composition in Tomato Fruits (Solanum lycopersicum L.) Grown in Magnesium Excess Soils. Front. Plant Sci. 2020, 11, 562399. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.D.; Liu, H.X.; Wang, Y.P.; Guo, J.H. Induction of Drought Tolerance in Cucumber Plants by a Consortium of Three Plant Growth-Promoting Rhizobacterium Strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef] [PubMed]
- Bocchini, M.; D’Amato, R.; Ciancaleoni, S.; Fontanella, M.C.; Palmerini, C.A.; Beone, G.M.; Onofri, A.; Negri, V.; Marconi, G.; Albertini, E.; et al. Soil Selenium (Se) Biofortification Changes the Physiological, Biochemical and Epigenetic Responses to Water Stress in Zea mays L. by Inducing a Higher Drought Tolerance. Front. Plant Sci. 2018, 9, 389. [Google Scholar] [CrossRef]
- Dobosy, P.; Vetési, V.; Sandil, S.; Endrédi, A.; Kröpfl, K.; Óvári, M.; Takács, T.; Rékási, M.; Záray, G. Effect of Irrigation Water Containing Iodine on Plant Physiological Processes and Elemental Concentrations of Cabbage (Brassica Oleracea l. Var. Capitata l.) and Tomato (Solanum lycopersicum L.) Cultivated in Different Soils. Agronomy 2020, 10, 720. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Saifullah; Ashraf, M.Y.; Ehsanullah. Role of Mineral Nutrition in Alleviation of Drought Stress in Plants Ejaz. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar] [CrossRef]
- Hameed, A.; Bibi, N.; Akhter, J.; Iqbal, N. Differential Changes in Antioxidants, Proteases, and Lipid Peroxidation in Flag Leaves of Wheat Genotypes under Different Levels of Water Deficit Conditions. Plant Physiol. Biochem. 2011, 49, 178–185. [Google Scholar] [CrossRef]
- Mirdad, Z.M. Effect of K+ and Salicylic Acid on Broccoli (Brassica oleraceae Var. Italica) Plants Grown Under Saline Water Irrigation. J. Agric. Sci. 2014, 6, 57–66. [Google Scholar] [CrossRef]
- El-Tohamy, W.A.; El-Abagy, H.M.; Badr, M.A.; Gruda, N. Drought Tolerance and Water Status of Bean Plants (Phaseolus vulgaris L.) as Affected by Citric Acid Application. J. Appl. Bot. Food Qual. 2013, 86, 212–216. [Google Scholar] [CrossRef]
- Mekki, B.E.D.; Hussien, H.A.; Salem, H. Role of Glutathione, Ascorbic Acid and α-Tocopherol in Alleviation of Drought Stress in Cotton Plants. Int. J. ChemTech Res. 2015, 8, 1573–1581. [Google Scholar]
- Ali, Z.; Basra, S.M.A.; Munir, H.; Mahmood, A.; Yousaf, S. Mitigation of Drought Stress in Maize by Natural and Synthetic Growth Promoters. J. Agric. Soc. Sci. 2011, 7, 56–62. [Google Scholar]
- Yasmeen, A.; Basra, S.M.A.; Farooq, M.; Rehman, H.u.; Hussain, N.; Athar, H.u.R. Exogenous Application of Moringa Leaf Extract Modulates the Antioxidant Enzyme System to Improve Wheat Performance under Saline Conditions. Plant Growth Regul. 2013, 69, 225–233. [Google Scholar] [CrossRef]
- Rademacher, W. Plant Growth Regulators: Backgrounds and Uses in Plant Production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Bargaz, A.; Nassar, R.M.A.; Rady, M.M.; Gaballah, M.S.; Thompson, S.M.; Brestic, M.; Schmidhalter, U.; Abdelhamid, M.T. Improved Salinity Tolerance by Phosphorus Fertilizer in Two Phaseolus vulgaris Recombinant Inbred Lines Contrasting in Their P-Efficiency. J. Agron. Crop Sci. 2016, 202, 497–507. [Google Scholar] [CrossRef]
- Cakmak, I.; Engels, C. Role of Mineral Nutrients in Photosynthesis and Yield Formation. In Mineral Nutrition of Crops: Fundamental Mechanisms and Implications; Food Products Press: Binghamton, NY, USA, 1999; pp. 141–168. [Google Scholar]
- Duca, M. Plant Physiology; Springer: Cham, Switzerland, 2015; p. 315. [Google Scholar]
- El-Mageed, T.A.A.; Semida, W.M.; Abdou, N.M.; El-Mageed, S.A.A. Coupling Effects of Potassium Fertilization Rate and Application Time on Growth and Grain Yield of Wheat (Triticum aestivum L.) Plants Grown Under Cd-Contaminated Saline Soil. J. Soil Sci. Plant Nutr. 2023, 23, 1070–1084. [Google Scholar] [CrossRef]
- Gaafar, A.A.; Ali, S.I.; El-Shawadfy, M.A.; Salama, Z.A.; Sekara, A.; Ulrichs, C.; Abdelhamid, M.T. Ascorbic Acid Induces the Increase of Secondary Metabolites, Antioxidant Activity, Growth, and Productivity of the Common Bean under Water Stress Conditions. Plants 2020, 9, 627. [Google Scholar] [CrossRef]
- Rady, M.M.; Sadak, M.S.; El-Lethy, S.R.; Elhamid, E.M.A.; Abdelhamid, M.T. Exogenous α-Tocopherol Has a Beneficial Effect on Glycine max (L.) Plants Irrigated with Diluted Sea Water. J. Hortic. Sci. Biotechnol. 2015, 90, 195–202. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-Mageed, T.A.; Mohamed, S.E.; El-Sawah, N.A. Combined Effect of Deficit Irrigation and Foliar-Applied Salicylic Acid on Physiological Responses, Yield, and Water-Use Efficiency of Onion Plants in Saline Calcareous Soil. Arch. Agron. Soil Sci. 2017, 63, 1227–1239. [Google Scholar] [CrossRef]
- Abd El–Mageed, T.A.; Ahmed Shaaban; Abd El-Mageed, S.A.; Semida, W.M.; Rady, M.O.A. Silicon Defensive Role in Maize (Zea mays L.) against Drought Stress and Metals-Contaminated Irrigation Water. Silicon 2021, 13, 2165–2176. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, H.; Snider, J.L.; Zahoor, R.; Iqbal, B.; Chen, B.; Meng, Y.; Zhou, Z. A Comparative Study of Integrated Crop Management System vs. Conventional Crop Management System for Cotton Yield and Fiber Quality with Respect to Fruiting Position under Different Soil Fertility Levels. Front. Plant Sci. 2018, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Nankishore, A.; Farrell, A.D. The Response of Contrasting Tomato Genotypes to Combined Heat and Drought Stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef]
- Abdou, N.M.; Abdel-razek, M.A.; El-mageed, S.A.A.; Semida, W.M. High Nitrogen Fertilization Modulates Morpho-Physiological Responses, Yield, and Water Productivity of Lowland Rice under Deficit Irrigation. Agronomy 2021, 11, 1291. [Google Scholar] [CrossRef]
- Navarro, M.; Munné-Bosch, S. Reduced Phosphate Availability Improves Tomato Quality Through Hormonal Modulation in Developing Fruits. J. Plant Growth Regul. 2022, 41, 153–162. [Google Scholar] [CrossRef]
- Semida, W.M.; El-Mageed, T.A.A.; Abdelkhalik, A.; Hemida, K.A.; Abdurrahman, H.A.; Howladar, S.M.; Leilah, A.A.A.; Rady, M.O.A. Selenium Modulates Antioxidant Activity, Osmoprotectants, and Photosynthetic Efficiency of Onion under Saline Soil Conditions. Agronomy 2021, 11, 855. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.; Abd El-Mageed, T.A.; Abd El-Mageed, S.A.; Rady, M.M.; Ali, E.F. Foliar Application of Zinc Oxide Nanoparticles Promotes Drought Stress Tolerance in Eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef]
- Etienne, P.; Diquelou, S.; Prudent, M.; Salon, C.; Maillard, A.; Ourry, A. Macro and Micronutrient Storage in Plants and Their Remobilization When Facing Scarcity: The Case of Drought. Agriculture 2018, 8, 14. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of Drought Stress on Photosynthesis and Photosynthetic Electron Transport Chain in Young Apple Tree Leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-Induced Drought Stress Tolerance in Mung Bean: Coordinated Roles of the Antioxidant Defence and Methylglyoxal Detoxification Systems. AoB Plants 2015, 7, plv069. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, S.S.; Kusvuran, S. Integrative Application of Cyanobacteria and Antioxidants Improves Common Bean Performance under Saline Conditions. Sci. Hortic. 2018, 233, 61–69. [Google Scholar] [CrossRef]
- El-Tayeb, M.A. Response of Barley Grains to the Interactive Effect of Salinity and Salicylic Acid. Plant Growth Regul. 2005, 45, 215–224. [Google Scholar] [CrossRef]
- Asrar, A.W.A.; Elhindi, K.M. Alleviation of Drought Stress of Marigold (Tagetes erecta) Plants by Using Arbuscular Mycorrhizal Fungi. Saudi J. Biol. Sci. 2011, 18, 93–98. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Santhanakrishnan, P.; Balasubramanian, P. Responses of Field Grown Tomato Plants to Arbuscular Mycorrhizal Fungal Colonization under Varying Intensities of Drought Stress. Sci. Hortic. 2006, 107, 245–253. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Wahid, A.; Ahmad, N.; Saleem, B.A. Improving the Drought Tolerance in Rice (Oryza sativa L.) by Exogenous Application of Salicylic Acid. J. Agron. Crop Sci. 2009, 195, 237–246. [Google Scholar] [CrossRef]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient Use Efficiency in Plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Higa, T.; Parr, J.F. Beneficial and Effective Microorganisms for a Sustainable Agriculture and Environment; International Nature Farming Research Center: Atami, Japan, 1994; Volume 1. [Google Scholar]
- Wang, W.; Zheng, W.; Lv, H.; Liang, B.; Jin, S.; Li, J.; Zhou, W. Animal-Derived Plant Biostimulant Alleviates Drought Stress by Regulating Photosynthesis, Osmotic Adjustment, and Antioxidant Systems in Tomato Plants. Sci. Hortic. 2022, 305, 111365. [Google Scholar] [CrossRef]
- Sikder, R.K.; Wang, X.; Zhang, H.; Gui, H.; Dong, Q.; Jin, D.; Song, M. Nitrogen Enhances Salt Tolerance by Modulating the Antioxidant Defense System and Osmoregulation Substance Content in Gossypium Hirsutum. Plants 2020, 9, 450. [Google Scholar] [CrossRef]
- Abdelkhalik, A.; Abd El-Mageed, T.A.; Mohamed, I.A.A.; Semida, W.M.; Al-Elwany, O.A.A.I.; Ibrahim, I.M.; Hemida, K.A.; El-Saadony, M.T.; AbuQamar, S.F.; El-Tarabily, K.A.; et al. Soil Application of Effective Microorganisms and Nitrogen Alleviates Salt Stress in Hot Pepper (Capsicum annum L.) Plants. Front. Plant Sci. 2023, 13, 1079260. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in Plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Karlidag, H.; Yildirim, E.; Turan, M. Salicylic Acid Ameliorates the Adverse Effect. Sci. Agric. 2009, 66, 180–187. [Google Scholar] [CrossRef]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of Salicylic Acid-Induced Phenolic Compounds and Raised Activities of Secondary Metabolic and Antioxidative Enzymes in Salvia miltiorrhiza Cell Culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of Exogenous Salicylic Acid under Changing Environment: A Review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Al-Elwany, O.A.A.I.; Hemida, K.A.; Abdel-Razek, M.A.; El-Mageed, T.A.A.; El-Saadony, M.T.; AbuQamar, S.F.; El-Tarabily, K.A.; Taha, R.S. Impact of Folic Acid in Modulating Antioxidant Activity, Osmoprotectants, Anatomical Responses, and Photosynthetic Efficiency of Plectranthus amboinicus Under Salinity Conditions. Front. Plant Sci. 2022, 13, 887091. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Al Mahmud, J.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, H.; Sahoo, L.; Panda, S.K. Molecular Physiology of Osmotic Stress in Plants. In Molecular Stress Physiology of Plants; Rout, G.R., Das, A.B., Eds.; Springer: New Delhi, India, 2013; pp. 179–192. [Google Scholar]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of Plants to Water Stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.N.; Kakani, R.K.; Sharma, L.K.; Agarwal, D.; John, S.; Sharma, Y. Effect of Water Stress on Morpho-Physiological Parameters of Fenugreek (Trigonella foenum-graecum L.) Genotypes. Legum. Res. 2019, 42, 60–65. [Google Scholar] [CrossRef]
- Roy, R.; Núñez-Delgado, A.; Sultana, S.; Wang, J.; Munir, A.; Battaglia, M.L.; Sarker, T.; Seleiman, M.F.; Barmon, M.; Zhang, R. Additions of Optimum Water, Spent Mushroom Compost and Wood Biochar to Improve the Growth Performance of Althaea rosea in Drought-Prone Coal-Mined Spoils. J. Environ. Manag. 2021, 295, 113076. [Google Scholar] [CrossRef]
- El-beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, L.; Wu, X.; Chen, W.; Song, D.; et al. Phosphorous Fertilization Alleviates Drought Effects on Alnus cremastogyne by Regulating Its Antioxidant and Osmotic Potential. Sci. Rep. 2018, 8, 5644. [Google Scholar] [CrossRef]
- Cruz De Carvalho, M.H. Drought Stress and Reactive Oxygen Species: Production, Scavenging and Signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant Sugars Are Crucial Players in the Oxidative Challenge during Abiotic Stress: Extending the Traditional Concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef]
- Kang, Y.; Yao, Y.; Liu, Y.; Shi, M.; Zhang, W.; Zhang, R.; Li, H.; Qin, S.; Yang, X. Exogenous Glutathione Enhances Tolerance of the Potato (Solanum tuberosum L.) to Cadmium Stress by Regulating the Biosynthesis of Phenylpropanoid and the Signal Transduction of Plant Hormones. Chem. Biol. Technol. Agric. 2023, 10, 24. [Google Scholar] [CrossRef]
- Azcón, R.; Gómez, M.; Tobar, R. Physiological and Nutritional Responses by Lactuca sativa L. to Nitrogen Sources and Mycorrhizal Fungi under Drought Conditions. Biol. Fertil. Soils 1996, 22, 156–161. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Fan, D.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Effect of Drought on Pigments, Osmotic Adjustment and Antioxidant Enzymes in Six Woody Plant Species in Karst Habitats of Southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Kang, G.Z.; Li, G.Z.; Liu, G.Q.; Xu, W.; Peng, X.Q.; Wang, C.Y.; Zhu, Y.J.; Guo, T.C. Exogenous Salicylic Acid Enhances Wheat Drought Tolerance by Influence on the Expression of Genes Related to Ascorbate-Glutathione Cycle. Biol. Plant. 2013, 57, 718–724. [Google Scholar] [CrossRef]
- Ghassemi, S.; Ghassemi-Golezani, K.; Salmasi, S.Z. Changes in Antioxidant Enzymes Activities and Physiological Traits of Ajowan in Response to Water Stress and Hormonal Application. Sci. Hortic. 2019, 246, 957–964. [Google Scholar] [CrossRef]
- Rehman, H.u.; Alharby, H.F.; Bamagoos, A.A.; Abdelhamid, M.T.; Rady, M.M. Sequenced Application of Glutathione as an Antioxidant with an Organic Biostimulant Improves Physiological and Metabolic Adaptation to Salinity in Wheat. Plant Physiol. Biochem. 2021, 158, 43–52. [Google Scholar] [CrossRef]
- Jesus, C.; Meijón, M.; Monteiro, P.; Correia, B.; Amaral, J.; Escandón, M.; Cañal, M.J.; Pinto, G. Salicylic Acid Application Modulates Physiological and Hormonal Changes in Eucalyptus Globulus under Water Deficit. Environ. Exp. Bot. 2015, 118, 56–66. [Google Scholar] [CrossRef]
- Wang, B.; Lai, T.; Huang, Q.W.; Yang, X.M.; Shen, Q.R. Effect of N Fertilizers on Root Growth and Endogenous Hormones in Strawberry Project Supported by the National High Technology Research and Development Program (863 Program) of China (No. 2004AA246080) and the Program for the Development of High-Tech Indu. Pedosphere 2009, 19, 86–95. [Google Scholar] [CrossRef]
- Akter, N.; Rafiqul Islam, M.; Abdul Karim, M.; Hossain, T. Alleviation of Drought Stress in Maize by Exogenous Application of Gibberellic Acid and Cytokinin. J. Crop Sci. Biotechnol. 2014, 17, 41–48. [Google Scholar] [CrossRef]
- Wang, H.; Liu, X.; Yang, P.; Wu, R.; Wang, S.; He, S.; Zhou, Q. Potassium Application Promote Cotton Acclimation to Soil Waterlogging Stress by Regulating Endogenous Protective Enzymes Activities and Hormones Contents. Plant Physiol. Biochem. 2022, 185, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Klute, A.; Dirksen, C. Hydraulic Conductivity and Diffusivity: Laboratory Methods. In Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1986; Volume 9, pp. 687–734. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis Part 2. Chemical and Microbiological Properties; American Society of Agronomy, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- Livens, I. Contribution à L’étude de Azotemineralisable Du Sol. Agric Louvain 1959, 7, 27–44. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall India Pvt. Ltd.: New Delhi, India, 1973. [Google Scholar]
- Dahnke, W.C.; Whitney, D.A. Measurement of Soil Salinity. In Recommended Chemical Soil Test Procedures for the North Central Region; North Central Regional Publication, 221; Dahnke, W.C., Ed.; North Dakota Agricultural Experiment Station Bulletin: Fargo, ND, USA, 1988; Volume 499, pp. 32–34. [Google Scholar]
- Victor, M.; Ponce, R.P.P.; Ercan, S. Characterization of Drought across the Climate Spectrum. J. Hydrol. Eng. ASCE 2000, 5, 222–224. [Google Scholar]
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semida, W.M. Selenium Application in Two Methods Promotes Drought Tolerance in Solanum Lycopersicum Plant by Inducing the Antioxidant Defense System. Sci. Hortic. 2020, 266, 109290. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements, Irrigation and Drainage Paper No. 56; FAO: Rome, Italy, 1998; ISBN 0254-5284. [Google Scholar]
- Fernández, J.E.; Alcon, F.; Diaz-espejo, A.; Hernandez-santana, V.; Cuevas, M.V. Water Use Indicators and Economic Analysis for On-Farm Irrigation Decision: A Case Study of a Super High Density Olive Tree Orchard. Agric. Water Manag. 2020, 237, 106074. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and Ascorbic Acid in Spinach (Spinacia oleracea) Chloroplasts. The Effect of Hydrogen Peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef]
- Sharma, S.K.; Le Maguer, M. Lycopene in Tomatoes and Tomato Pulp Fractions. Ital. J. Food Sci. 1996, 2, 107–113. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000; ISBN 0935584676. [Google Scholar]
- Pequerul, A.; Pérez, C.; Madero, P.; Val, J.; Monge, E. A Rapid Wet Digestion Method for Plant Analysis. In Optimization of Plant Nutrition; Fragoso, M.A.C., Van Beusichem, M.L., Houwers, A., Eds.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 1993; Volume 53. [Google Scholar] [CrossRef]
- Lien, C.C.; Ay, C.; Ting, C.H. Non-Destructive Impact Test for Assessment of Tomato Maturity. J. Food Eng. 2009, 91, 402–407. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Method Tomato Masayasu * National NAGATA * and Ichiji YAMASHITA * of Vegetables Rnamental Plants and Tea, Ministry of Agriculture, Forestry and Fisheries. Forestry 1992, 39, 1–4. [Google Scholar]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenol-Oxidase in Beta vulgaris L. Plant Physiol. 1949, 24, 1–5. [Google Scholar] [CrossRef]
- Avron, M. Photophosphorylation by Swiss-Chard Chloroplasts. BBA—Biochim. Biophys. Acta 1960, 40, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) Response to Ozone Exposure Assessed with a Chlorophyll a Fluorescence Performance Index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Page, A.I.; Miller, R.H.; Keeny, D.R. Methods of Soil Analysis. In Part II. Chemical and Microbiological Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 225–246. [Google Scholar]
- Johnson, C.M.; Ulrich, A. Analytical Methods for Use in Plant Analysis; Bulletin (Agricultural Experiment Station, Berkeley, Calif.); University of California: Berkeley, CA, USA, 1959. [Google Scholar]
- Rady, M.M. Effect of 24-Epibrassinolide on Growth, Yield, Antioxidant System and Cadmium Content of Bean (Phaseolus vulgaris L.) Plants under Salinity and Cadmium Stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Osman, A.S.; Rady, M.M. Effect of Humic Acid as an Additive to Growing Media to Enhance the Production of Eggplant and Tomato Transplants. J. Hortic. Sci. Biotechnol. 2014, 89, 237–244. [Google Scholar] [CrossRef]
- Rady, M.M.; Rehman, H.u. Supplementing Organic Biostimulants into Growing Media Enhances Growth and Nutrient Uptake of Tomato Transplants. Sci. Hortic. 2016, 203, 192–198. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative Parameters in the Seedlings of Pigeonpea (Cajanus cajan (L.) Millspaugh) in Response to Zn and Ni Stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants: Protective Role of Exogenous Polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Kubiś, J. Exogenous Spermidine Differentially Alters Activities of Some Scavenging System Enzymes, H2O2 and Superoxide Radical Levels in Water-Stressed Cucumber Leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 207, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water Stress Induced Changes in Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa (Medicago sativa) Plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Huang, C.; He, W.; Guo, J.; Chang, X.; Su, P.; Zhang, L. Increased Sensitivity to Salt Stress in an Ascorbate-Deficient Arabidopsis Mutant. J. Exp. Bot. 2005, 56, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, A.; Berardino, R.; De Pinto, M.C.; Sanità Di Toppi, L.; Storelli, M.M.; Tommasi, F.; De Gara, L. Increase in Ascorbate-Glutathione Metabolism as Local and Precocious Systemic Responses Induced by Cadmium in Durum Wheat Plants. Plant Cell Physiol. 2008, 49, 362–374. [Google Scholar] [CrossRef]
- Nagy, Z.; Daood, H.; Koncsek, A.; Molnár, H.; Helyes, L. The Simultaneous Determination of Capsaicinoids, Tocopherols, and Carotenoids in Pungent Pepper Powder. J. Liq. Chromatogr. Relat. Technol. 2017, 40, 199–209. [Google Scholar] [CrossRef]
- Konings, E.J.M.; Roomans, H.H.S.; Beljaars, P.R. Liquid Chromatographic Determination of Tocopherols and Tocotrienols in Margarine, Infant Foods, and Vegetables. J. AOAC Int. 1996, 79, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Ching, L.S.; Mohamed, S. Alpha-Tocopherol Content in 62 Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.; Ries, S. Superoxide Dismutases. I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Foster, J.G.; Hess, J.L. Responses of Superoxide Dismutase and Glutathione Reductase Activities in Cotton Leaf Tissue Exposed to an Atmosphere Enriched in Oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef]
- Yu, Q.; Rengel, Z. Drought and Salinity Differentially Influence Activities of Superoxide Dismutases in Narrow-Leafed Lupins. Plant Sci. 1999, 142, 1–11. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification Efficiency: Linking Baseline and Bias in the Analysis of Quantitative PCR Data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, 45. [Google Scholar] [CrossRef] [PubMed]
- Nehela, Y.; Hijaz, F.; Elzaawely, A.A.; El-Zahaby, H.M.; Killiny, N. Phytohormone Profiling of the Sweet Orange (Citrus sinensis (L.) Osbeck) Leaves and Roots Using GC-MS-Based Method. J. Plant Physiol. 2016, 199, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.M.; Talaat, N.B.; Abdelhamid, M.T.; Shawky, B.T.; Desoky, E.S.M. Maize (Zea mays L.) Grains Extract Mitigates the Deleterious Effects of Salt Stress on Common Bean (Phaseolus vulgaris L.) Growth and Physiology. J. Hortic. Sci. Biotechnol. 2019, 94, 777–789. [Google Scholar] [CrossRef]
- Ünyayar, S.; Topcuoglu, S.F.; Ünyayar, A. A Modified Method for Extraction and Identification of Indole-3-Acetic Acid (IAA), Gibberellic Acid (GA3), Abscisic Acid (ABA) and Zeatin Produced by Phanerochaete Chrysosporium ME446. Bulg. J. Plant Physiol. 1996, 22, 105–110. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1984; pp. 20–212. [Google Scholar]
- Shapiro, A.S.S.; Wilk, M.B. Biometrika Trust An Analysis of Variance Test for Normality (Complete Samples) Published by: Oxford University Press on Behalf of Biometrika Trust Stable. Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | PLN | PLA (m2) | PSDW (g) | PFW (kg) | PFN | HFY (ton) | WUE (kg Fruits m−3 Water) | |
|---|---|---|---|---|---|---|---|---|
| ETc | NABF | |||||||
| 100% | Without | 59.1 ± 4.80 c | 5.19 ± 0.38 c | 63.4 ± 4.8 c | 5.18 ± 0.42 c | 32.4 ± 3.10 c | 69.1 ± 5.2 c | 12.14 ± 1.7 d |
| With | 79.0 ± 6.21 a | 6.98 ± 0.50 a | 99.3 ± 7.2 a | 9.30 ± 0.82 a | 54.1 ± 4.14 a | 121.2 ± 9.8 a | 21.29 ± 1.9 a | |
| 80% | Without | 45.3 ± 3.44 d | 4.01 ± 0.31 d | 49.8 ± 3.5 d | 2.76 ± 0.11 d | 19.7 ± 1.35 d | 36.8 ± 2.41 d | 8.08 ± 1.2 e |
| With | 63.4 ± 5.30 b | 5.54 ± 0.38 b | 71.2 ± 5.7 b | 5.64 ± 0.44 b | 36.1 ± 3.01 b | 76.2 ± 6.3 b | 16.73 ± 1.6 c | |
| 60% | Without | 32.1 ± 2.82 e | 2.70 ± 0.13 e | 33.2 ± 2.8 e | 0.81 ± 0.08 e | 10.1 ± 0.91 e | 20.2 ± 0.8 e | 5.91 ± 0.71 f |
| With | 58.4 ± 4.72 c | 5.05 ± 0.34 c | 63.1 ± 4.9 c | 5.10 ± 0.40 c | 31.7 ± 2.51 c | 68.4 ± 5.1 c | 20.02 ± 1.3 b | |
| Significance of F test: | ||||||||
| ETc | * | * | ** | * | ** | * | ** | |
| NABF | ** | ** | ** | ** | ** | ** | ** | |
| ETc × NABF | * | * | ** | * | ** | * | ** | |
| Treatments | Vit C (mg 100 g−1 Fruit FW) | TSS (°Brix) | TA (% Citric Acid) | Lycopene (mg 100 g−1 Fruit FW) | β-Car (mg 100 g−1 Fruit FW) | Firmness (kg cm−1) | Selenium (mg kg−1 FW) | Iodine (mg kg−1 FW) | |
|---|---|---|---|---|---|---|---|---|---|
| ETc | NABF | ||||||||
| 100% | Without | 5.86 ± 0.15 d | 4.89 ± 0.38 d | 0.39 ± 0.09 e | 4.68 ± 0.19 e | 2.02 ± 0.07 e | 4.60 ± 0.24 c | 3.02 ± 0.08 c | 1.18 ± 0.04 c |
| With | 6.39 ± 0.18 c | 5.28 ± 0.44 c | 0.45 ± 0.12 d | 4.99 ± 0.21 d | 2.25 ± 0.09 d | 4.99 ± 0.27 a | 3.62 ± 0.10 a | 1.48 ± 0.05 a | |
| 80% | Without | 6.41 ± 0.17 c | 5.26 ± 0.46 c | 0.50 ± 0.14 c | 5.24 ± 0.22 c | 2.46 ± 0.10 c | 4.32 ± 0.22 d | 2.52 ± 0.06 d | 1.02 ± 0.03 d |
| With | 7.09 ± 0.20 b | 5.69 ± 0.49 b | 0.59 ± 0.18 b | 5.72 ± 0.25 b | 2.78 ± 0.11 b | 4.74 ± 0.25 b | 3.34 ± 0.07 b | 1.32 ± 0.03 b | |
| 60% | Without | 7.11 ± 0.20 b | 5.70 ± 0.50 b | 0.58 ± 0.19 b | 5.78 ± 0.26 b | 2.80 ± 0.11 b | 4.10 ± 0.18 e | 2.12 ± 0.05 e | 0.82 ± 0.02 e |
| With | 7.94 ± 0.24 a | 6.27 ± 0.54 a | 0.70 ± 0.21 a | 6.38 ± 0.30 a | 3.22 ± 0.12 a | 4.61 ± 0.25 c | 3.01 ± 0.08 c | 1.19 ± 0.04 c | |
| Significance of F test: | |||||||||
| ETc | * | * | * | * | ** | * | ** | ** | |
| NABF | * | ** | ** | * | ** | * | ** | ** | |
| ETc × NABF | * | * | * | * | ** | * | ** | ** | |
| Treatments | SPAD Value | Chlorophyll | Carotenoids | Fv/Fm | PI | PhAc | |
|---|---|---|---|---|---|---|---|
| ETc | NABF | mg g−1 FW | % | ||||
| 100% | Without | 47.6 ± 2.02 c | 3.22 ± 0.16 c | 0.36 ± 0.02 c | 0.79 ± 0.03 c | 16.8 ± 0.32 c | 49.1 ± 1.03 c |
| With | 56.8 ± 2.12 a | 3.52 ± 0.18 a | 0.47 ± 0.03 a | 0.88 ± 0.04 a | 19.2 ± 0.38 a | 58.9 ± 1.14 a | |
| 80% | Without | 42.1 ± 1.94 d | 2.87 ± 0.12 d | 0.32 ± 0.02 d | 0.73 ± 0.02 d | 14.9 ± 0.29 d | 44.1 ± 0.98 d |
| With | 52.8 ± 2.02 b | 3.36 ± 0.15 b | 0.43 ± 0.03 b | 0.84 ± 0.03 b | 17.9 ± 0.35 b | 54.6 ± 1.04 b | |
| 60% | Without | 29.8 ± 1.11 e | 2.11 ± 0.10 e | 0.24 ± 0.01 e | 0.68 ± 0.02 e | 12.1 ± 0.22 e | 34.4 ± 0.82 e |
| With | 47.3 ± 2.01 c | 3.23 ± 0.17 c | 0.36 ± 0.02 c | 0.79 ± 0.03 c | 16.6 ± 0.33 c | 48.9 ± 1.06 c | |
| Significance of F test: | |||||||
| ETc | * | ** | ** | * | * | * | |
| NABF | ** | ** | ** | * | ** | * | |
| ETc × NABF | * | ** | ** | * | * | * | |
| Treatments | Nitrogen | Phosphorus | Potassium | Calcium | Magnesium | Sulfur | |
|---|---|---|---|---|---|---|---|
| ETc | NABF | mg g−1 DW | |||||
| 100% | Without | 18.6 ± 0.42 c | 1.99 ± 0.05 c | 18.1 ± 0.39 c | 6.88 ± 0.16 c | 1.69 ± 0.04 c | 0.84 ± 0.02 c |
| With | 26.4 ± 0.62 a | 2.52 ± 0.08 a | 25.2 ± 0.61 a | 8.78 ± 0.18 a | 2.14 ± 0.06 a | 1.12 ± 0.03 a | |
| 80% | Without | 14.2 ± 0.33 d | 1.73 ± 0.04 d | 15.3 ± 0.32 d | 5.42 ± 0.14 d | 1.46 ± 0.03 d | 0.62 ± 0.02 d |
| With | 21.2 ± 0.53 b | 2.28 ± 0.07 b | 22.8 ± 0.50 b | 8.15 ± 0.15 b | 1.94 ± 0.05 b | 0.98 ± 0.03 b | |
| 60% | Without | 11.3 ± 0.28 e | 1.45 ± 0.03 e | 12.1 ± 0.29 e | 3.94 ± 0.12 e | 1.12 ± 0.03 e | 0.42 ± 0.01 e |
| With | 18.8 ± 0.40 c | 1.98 ± 0.05 c | 18.2 ± 0.41 c | 6.90 ± 0.17 c | 1.68 ± 0.04 c | 0.83 ± 0.02 c | |
| Significance of F test: | |||||||
| ETc | ** | * | ** | ** | * | ** | |
| NABF | * | ** | ** | ** | ** | ** | |
| ETc × NABF | * | * | * | ** | * | ** | |
| Treatments | Iron | Manganese | Zinc | Copper | Selenium | Iodine | |
|---|---|---|---|---|---|---|---|
| ETc | NABF | mg kg−1 DW | |||||
| 100% | Without | 702 ± 16.8 c | 528 ± 13.2 c | 304 ± 7.2 c | 240 ± 6.1 c | 18.2 ± 0.46 c | 3.58 ± 0.12 c |
| With | 844 ± 18.8 a | 632 ± 14.8 a | 381 ± 8.4 a | 292 ± 7.2 a | 22.7 ± 0.61 a | 4.49 ± 0.15 a | |
| 80% | Without | 582 ± 14.5 d | 466 ± 12.6 d | 262 ± 6.4 d | 212 ± 5.4 d | 15.4 ± 0.34 d | 3.06 ± 0.13 d |
| With | 784 ± 15.4 b | 584 ± 14.0 b | 348 ± 7.8 b | 268 ± 6.7 b | 20.1 ± 0.41 b | 4.01 ± 0.14 b | |
| 60% | Without | 440 ± 12.9 e | 392 ± 11.1 e | 212 ± 5.3 e | 182 ± 4.6 e | 12.5 ± 0.29 e | 2.40 ± 0.07 e |
| With | 698 ± 16.7 c | 526 ± 13.0 c | 301 ± 7.1 c | 242 ± 6.3 c | 18.1 ± 0.48 c | 3.60 ± 0.13 c | |
| Significance of F test: | |||||||
| ETc | ** | * | ** | ** | * | ** | |
| NABF | * | ** | ** | ** | ** | ** | |
| ETc × NABF | * | * | * | ** | * | ** | |
| Treatments | S. Sugar Content | Free Proline | AsA Content | GSH Content | αToc | |
|---|---|---|---|---|---|---|
| ETc | NABF | (mg g−1 DW) | (mM g−1 DW) | (µM g−1 FW) | (µM g−1 DW) | |
| 100% | Without | 21.6 ± 0.11 d | 0.12 ± 0.003 f | 1.70 ± 0.03 d | 0.80 ± 0.02 f | 2.28 ± 0.05 d |
| With | 24.8 ± 0.13 c | 0.14 ± 0.004 e | 1.88 ± 0.04 c | 0.86 ± 0.02 e | 2.52 ± 0.06 c | |
| 80% | Without | 25.2 ± 0.13 c | 0.17 ± 0.005 d | 1.89 ± 0.04 c | 0.93 ± 0.03 d | 2.54 ± 0.06 c |
| With | 29.2 ± 0.15 b | 0.20 ± 0.006 c | 2.22 ± 0.05 b | 1.04 ± 0.03 c | 2.88 ± 0.07 b | |
| 60% | Without | 29.0 ± 0.15 b | 0.24 ± 0.007 b | 2.24 ± 0.05 b | 1.66 ± 0.04 b | 2.91 ± 0.07 b |
| With | 35.4 ± 0.18 a | 0.29 ± 0.008 a | 2.74 ± 0.06 a | 1.98 ± 0.04 a | 3.42 ± 0.08 a | |
| Significance of F test: | ||||||
| ETc | * | * | ** | ** | * | |
| NABF | ** | ** | ** | ** | ** | |
| ETc × NABF | * | * | ** | ** | * | |
| Treatments | SOD Activity | CAT Activity | APX Activity | GR Activity | SPC | |
|---|---|---|---|---|---|---|
| ETc | NABF | Unit g−1 Protein | µmol H2O2 min−1 g−1 Protein | g kg−1 DW | ||
| 100% | Without | 36.9 ± 0.85 e | 17.1 ± 0.38 f | 23.2 ± 0.42 e | 16.0 ± 0.24 d | 232 ± 5.2 c |
| With | 41.2 ± 0.92 d | 18.9 ± 0.42 e | 25.9 ± 0.48 d | 17.9 ± 0.28 c | 282 ± 6.7 a | |
| 80% | Without | 41.9 ± 0.93 d | 21.7 ± 0.48 d | 26.2 ± 0.49 d | 18.2 ± 0.30 c | 206 ± 4.8 d |
| With | 47.8 ± 0.97 c | 26.2 ± 0.50 c | 30.6 ± 0.55 c | 22.2 ± 0.35 b | 258 ± 5.8 b | |
| 60% | Without | 54.4 ± 1.02 b | 31.5 ± 0.55 b | 36.2 ± 0.59 b | 22.9 ± 0.36 b | 175 ± 3.9 e |
| With | 65.8 ± 1.10 a | 39.8 ± 0.63 a | 44.4 ± 0.70 a | 29.2 ± 0.42 a | 230 ± 5.1 c | |
| Significance of F test: | ||||||
| ETc | ** | ** | * | ** | * | |
| NABF | ** | ** | * | ** | * | |
| ETc × NABF | * | ** | * | ** | * | |
| Soil Layer (cm) | Particle Size Distribution | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Clay (%) | Silt (%) | Sand (%) | Texture | ||||||||
| 25.7 ± 0.62 | 32.7 ± 0.45 | 41.6 ± 0.52 | Loam | ||||||||
| 0–30 | BD (g cm−3) | Ksat (cm h−1) | FC (%) | WP (%) | AW (%) | ||||||
| 1.58 ± 0.03 | 1.89 ± 0.02 | 24.2 ± 0.34 | 11.4 ± 0.14 | 12.8 ± 0.11 | |||||||
| pH | 7.88 ± 0.03 | N (mg kg−1 soil) | 46.8 ± 0.87 | ||||||||
| ECe (dS m−1) | 10.2 ± 0.05 | P (mg kg−1 soil) | 5.12 ± 0.12 | ||||||||
| CEC (cmol kg−1) | 12.9 ± 0.06 | K (mg kg−1 soil) | 37.2 ± 0.67 | ||||||||
| CaCO3 (%) | 2.27 ± 0.04 | Fe (mg kg−1 soil) | 2.92 ± 0.03 | ||||||||
| OM (%) | 0.98 ± 0.01 | Mn (mg kg−1 soil) | 1.62 ± 0.03 | ||||||||
| ESP | 14.2 ± 0.11 | Zn (mg kg−1 soil) | 1.34 ± 0.02 | ||||||||
| SAR | 10.4 ± 0.06 | Cu (mg kg−1 soil) | 0.72 ± 0.01 | ||||||||
| The Component | % w/w | Molecular Formula | ppm | mM L−1 | MW |
|---|---|---|---|---|---|
| Urea (N) | 7.2 | CO(NH2)2 | 180 | 2.99 | 60.06 |
| MAP (P) | 9.2 | NH4H2PO4 | 230 | 1.99 | 115.03 |
| Potassium sulfate (K) | 20.9 | K2SO4 | 523 | 3.00 | 174.26 |
| Magnesium sulfate (Mg) | 7.2 | MgSO4 | 180 | 1.49 | 120.37 |
| Calcium nitrate (Ca) | 13.1 | Ca(NO3)2 | 328 | 1.99 | 164.09 |
| Sodium selenate (Se) | 3.0 | Na2SeO3 | 75 | 0.43 | 172.94 |
| Potassium iodide (I) | 2.0 | KI | 50 | 0.30 | 166.00 |
| Ascorbic acid (AsA) | 7.0 | C6H8O6 | 175 | 0.99 | 176.12 |
| Glutathione (GSH) | 6.1 | C10H17N3O6S | 152 | 0.49 | 307.32 |
| Salicylic acid (SA) | 3.3 | C19H19N7O6 | 82 | 0.59 | 138.12 |
| Citric acid (CA) | 15.4 | C6H8O7 | 385 | 2.00 | 192.12 |
| EDTA | 5.6 | C10H16N2O8 | 140 | 0.48 | 292.24 |
| Month | Day °C | Night °C | ARH (%) | AWS (km h−1) | AP (mm d−1) |
|---|---|---|---|---|---|
| 2020 and 2021 Seasons, Respectively | |||||
| February | 26.5 and 27.0 | 4.14 and 3.16 | 64.8 and 56.4 | 2.24 and 2.45 | 1.91 and 0.00 |
| March | 30.7 and 34.4 | 4.87 and 5.13 | 57.6 and 53.6 | 2.76 and 2.86 | 0.41 and 0.51 |
| April | 34.5 and 40.4 | 9.34 and 5.62 | 49.2 and 41.1 | 2.92 and 3.26 | 0.05 and 0.02 |
| May | 44.2 and 43.5 | 12.0 and 15.7 | 39.9 and 30.6 | 3.58 and 3.50 | 0.00 and 0.00 |
| June | 42.9 and 41.0 | 17.3 and 16.3 | 35.0 and 30.2 | 3.59 and 3.97 | 0.00 and 0.00 |
| Ion Concentrations (meq L−1) | EC (dS m−1) | pH | SAR | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CO32− | HCO3− | SO42− | Cl− | Mg2+ | Ca2+ | K+ | Na+ | |||
| 0.00 ± 0.00 | 2.10 ± 0.11 | 3.27 ± 0.19 | 12.1 ± 0.46 | 1.76 ± 0.16 | 5.38 ± 0.34 | 1.42 ± 0.14 | 6.16 ± 0.64 | 1.71 ± 0.21 | 7.51 ± 0.71 | 2.94 ± 0.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Mageed, T.A.; Ihab, R.; Rady, M.M.; Belal, H.E.E.; Mostafa, F.A.; Galal, T.M.; Masoudi, L.M.A.; Ali, E.F.; Roulia, M.; Mahmoud, A.E.M. A Novel Nutrient- and Antioxidant-Based Formulation Can Sustain Tomato Production under Full Watering and Drought Stress in Saline Soil. Plants 2023, 12, 3407. https://doi.org/10.3390/plants12193407
Abd El-Mageed TA, Ihab R, Rady MM, Belal HEE, Mostafa FA, Galal TM, Masoudi LMA, Ali EF, Roulia M, Mahmoud AEM. A Novel Nutrient- and Antioxidant-Based Formulation Can Sustain Tomato Production under Full Watering and Drought Stress in Saline Soil. Plants. 2023; 12(19):3407. https://doi.org/10.3390/plants12193407
Chicago/Turabian StyleAbd El-Mageed, Taia A., Radwa Ihab, Mostafa M. Rady, Hussein E. E. Belal, Fatma A. Mostafa, Tarek M. Galal, Luluah M. Al Masoudi, Esmat F. Ali, Maria Roulia, and Amr E. M. Mahmoud. 2023. "A Novel Nutrient- and Antioxidant-Based Formulation Can Sustain Tomato Production under Full Watering and Drought Stress in Saline Soil" Plants 12, no. 19: 3407. https://doi.org/10.3390/plants12193407