
The Perspective of Arctic–Alpine Species in Southernmost Localities: The Example of Kalmia procumbens in the Pyrenees and Carpathians
 , , , ,
, , , ,
Abstract
:1. Introduction
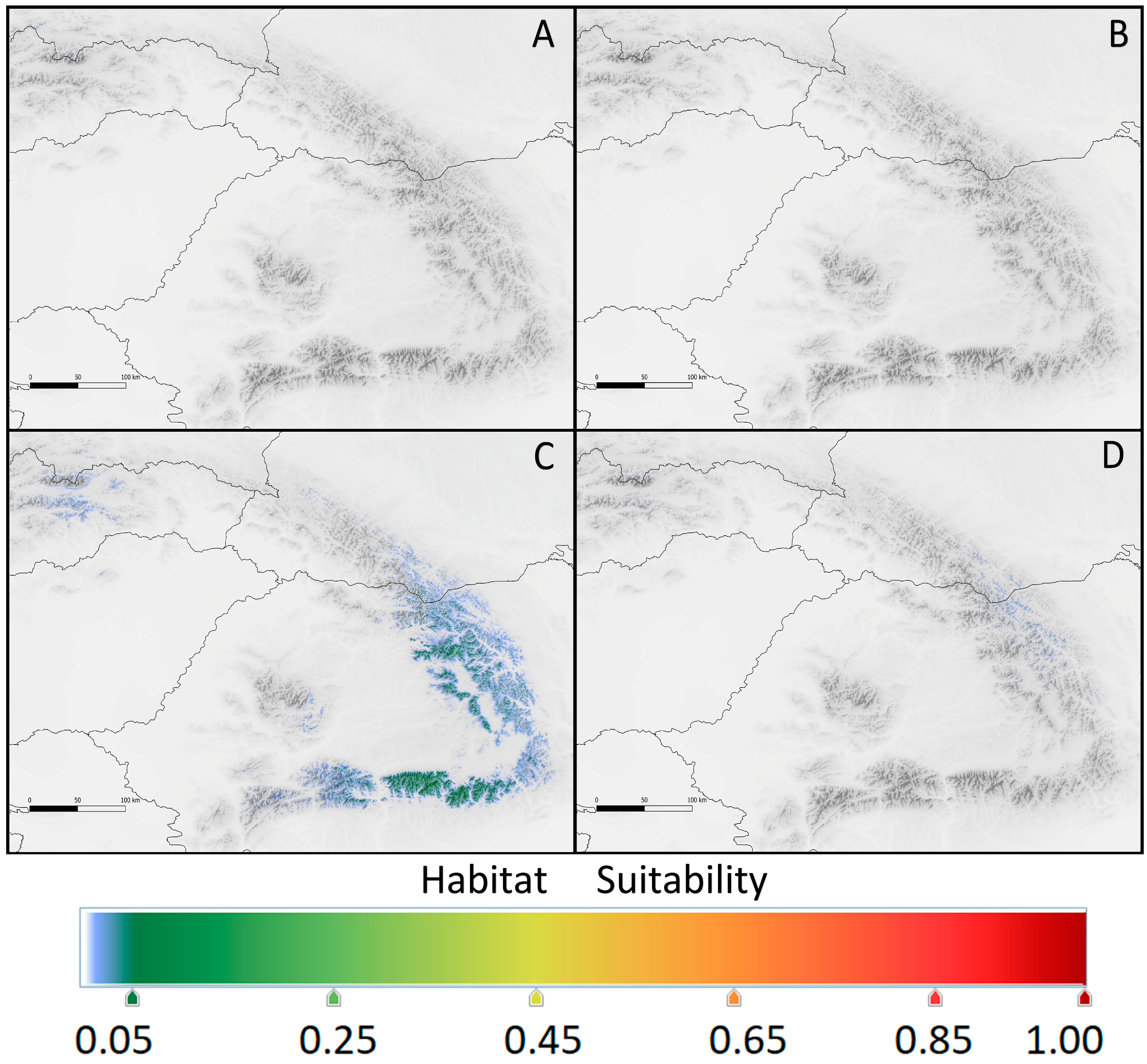
2. Results
2.1. Realized Geographic Niches
2.2. Future Geographic Niches
3. Discussion
3.1. Realized Potential Niches in the Carpathians and Pyrenees
3.2. Environmental Conditions of the Realized Niches of K. procumbens
3.3. Possible Influence of Climate Differences
3.4. Future Ecological Niches in the Carpathians and Pyrenees
4. Materials and Methods
4.1. Study Areas
4.2. Data Sampling and Geographic Analyses
4.3. Environmental Variables
4.4. Niche Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hultén, E. Outline of the History of Arctic and Boreal Biota during the Quarternary Period; Bokförlags Aktiebolaget Thule: Stockholm, Sweden, 1937. [Google Scholar]
- Billings, W.D. Adaptations and origins of alpine plants. Arct. Alp. Res. 1974, 6, 129–142. [Google Scholar] [CrossRef]
- Hagen, O.; Vaterlaus, L.; Albouy, C.; Brown, A.; Leugger, F.; Onstein, R.E.; Novaes de Santana, C.; Scotese, C.R.; Pellissier, L. Mountain building, climate cooling and the richness of cold-adapted plants in the Northern Hemisphere. J. Biogeogr. 2019, 46, 1792–1807. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of the high mountain systems of Europe: An overview. In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Mercè Aniz, M., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2017; Volume 62, pp. 63–74. [Google Scholar]
- Fischer, A.; Fickert, T.; Schwaizer, G.; Patzelt, G.; Groß, G. Vegetation dynamics in Alpine glacier forelands tackled from space. Sci. Rep. 2019, 9, 13918. [Google Scholar] [CrossRef]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe: 0–13000 Years Ago; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Ralska-Jasiewiczowa, M.; Latałowa, M.; Wasylikowa, K.; Tobolski, K.; Madeyska, E.; Wright, H.E.; Turner, C. Late Glacial and Holocene History of Vegetation in Poland Based on Isopollen Maps; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2004. [Google Scholar]
- Stivrins, N.; Soininen, J.; Amon, L.; Fontana, S.L.; Gryguc, G.; Heikkilä, M.; Heiri, O.; Kisielienè, D.; Reitalu, T.; Stančikaitè, M.; et al. Biotic turnover rates during the Pleistocene-Holocene transition. Quat. Sci. Rev. 2016, 151, 100–110. [Google Scholar] [CrossRef]
- Kornaś, J.; Medwecka-Kornaś, A. Geografia Roślin; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2002. [Google Scholar]
- Habel, J.C.; Assmann, T. Relict Species. Phylogeography and Conservation Biology; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Nat. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef]
- Lesica, P. Arctic-Alpine plants decline over two decades in Glacier National Park, Montana, USA. Arctic Antarct. Alp. Res. 2014, 46, 327–332. [Google Scholar] [CrossRef]
- Rixen, C.; Wipf, S.; Frei, E.; Stöckli, V. Faster, higher, more? Past, present and future dynamics of alpine and arctic flora under climate change. Alp. Bot. 2014, 124, 77–79. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; García-Calvo, L.; García, P.; Acebes, J.L. Anticipating extinctions of glacial relict populations in mountain refugia. Biol. Conserv. 2016, 201, 243–251. [Google Scholar] [CrossRef]
- Lesica, P.; Crone, E.E. Arctic and boreal plant species decline at their southern range limits in the Rocky Mountains. Ecol. Let. 2017, 20, 166–174. [Google Scholar] [CrossRef]
- Rixen, C.; Wipf, S. Non-equilibrium in alpine plant assemblages: Shifts in Europe’s summit floras. In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Mercè Aniz, M., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2017; Volume 62, pp. 285–303. [Google Scholar]
- Pauli, A.; Halloy, S.R.P. High mountain ecosystems under climate change. In Oxford Research Encyclopedia fo Climate Science; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Fragnière, Y.; Pittet, L.; Clément, B.; Bétrisey, S.; Gerber, E.; Ronikier, M.; Parisod, C.; Kozlowski, G. Climate change and alpine screes: No future for glacial relict Papaver occidentale (Papaveraceae) in Western Prealps. Diversity 2020, 12, 346. [Google Scholar] [CrossRef]
- Didukh, Y.P.; Chorney, I.I.; Budzhak, V.V.; Tokaryuk, A.I.; Kish, R.Y.; Protopopova, V.V.; Shevera, M.V.; Kozak, O.M.; Kontar, I.S.; Rozenblit, Y.V.; et al. Climatogenic Changes of Plant Life of the Ukrainian Carpathians; DrukArt: Chernivtsi, Ukraine, 2016. [Google Scholar]
- Trivedi, M.R.; Morecroft, M.D.; Berry, P.M.; Dawson, T.P. Potential effects of climate change on plant communities in three montane nature reserves in Scotland, UK. Biol. Conserv. 2008, 141, 1665–1675. [Google Scholar] [CrossRef]
- Kobiv, Y. Response of rare alpine plant species to climate change in the Ukrainian Carpathians. Folia Geobot. 2017, 52, 217–226. [Google Scholar] [CrossRef]
- Kobiv, Y. Trends in population size of rare plant species in the alpine habitats of the Ukrainian Carpathians under climate change. Diversity 2018, 10, 62. [Google Scholar] [CrossRef]
- Leunda, M.; González-Sampériz, P.; Gil-Romera, G.; Bartolomé, M.; Belmonte-Ribas, Á.; Gómez-García, D.; Kaltenrieder, P.; Rubiales, J.M.; Schwörer, C.; Tinner, W.; et al. Ice cave reveals environmental forcing of long-term Pyrenean tree line dynamics. J. Ecol. 2018, 107, 814–828. [Google Scholar] [CrossRef]
- Löffler, J.; Pape, R. Thermal niche predictors of alpine plant species. Ecology 2020, 101, e02891. [Google Scholar] [CrossRef]
- Hultén, E. The Amphi-Atlantic Plants and Their Phytogeographic Connections; Koeltz: Königstein, Germany, 1973. [Google Scholar]
- Meusel, H.; Jäger, E.; Rauschert, S.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora; Fischer: Jena, Germany, 1978; Volume 2. [Google Scholar]
- Hultén, E.; Fries, M. Atlas of North European Vascular Plants; Koelz: Königstein, Germany, 1986; Volume 1–3. [Google Scholar]
- Gómez García, D.; Ferrández Palacio, J.V.; Bernal Gálvez, M.; Campo González, A.; López Retamero, J.R.; Ezquerra Rivas, V. Plantas de las Cumbres del Pirineo; Instituto Pirenaicode Ecologia: Jaca, Spain; Prames: Zaragoza, Spain, 2020. [Google Scholar]
- Abeli, T.; Vamosi, J.C.; Orsenigo, S. The importance of marginal population hotspots of cold-adapted species for research on climate change and conservation. J. Biogeogr. 2018, 45, 977–985. [Google Scholar] [CrossRef]
- Dítě, D.; Hájek, M.; Svitková, I.; Košuthová, A.; Šoltés, R.; Kliment, J. Glacial-relict symptoms in the Western Carpathian flora. Folia Geobot. 2018, 53, 277–300. [Google Scholar] [CrossRef]
- Buchner, O.; Karadar, M.; Bauer, I.; Neuner, G. A novel system for in situ determination of heat tolerance of plants: First results on alpine dwarf shrubs. Plant Methods 2013, 9, 7. [Google Scholar] [CrossRef]
- Chopik, V.I. Vysokohirna Flora Ukrains’kykh Karpat; Naukova Dumka: Kyiv, Ukraine, 1976. [Google Scholar]
- Boratyński, A.; Didukh, Y.P. Loiseleuria procumbens (Ericaceae) in the Ukrainian Carpathians. Dendrobiology 2002, 47, 3–8. [Google Scholar]
- Boratyński, A.; Romo, A. Loiseleuria procumbens (Ericaceae) in the Spanish Pyrenees. Acta Soc. Bot. Pol. 2003, 72, 125–133. [Google Scholar] [CrossRef]
- Graebherr, G.; Gottfried, M.; Pauli, H. Climate effects on mountain plants. Nature 1994, 369, 448. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Körner, C. The green cover of mountains in a changing environment. In Global Change and Mountain Regions; Huber, U.M., Bugmann, K.M., Reasoner, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 23, pp. 367–376. [Google Scholar]
- Cherepanyn, R.M. Effect of climate changes on the habitat of rare arctic-alpine plant species in the high mountain part of the Ukrainian Carpathians. Biol. Stud. 2018, 12, 73–86. [Google Scholar] [CrossRef]
- Grime, J.P.; Price, S. The Evolutionary Strategies that Shape Ecosystems; John Wiley & Sons: West Sussex, UK, 2012. [Google Scholar]
- Didukh, Y.P. World of Plants of Ukraine in Aspect of the Climate Change; Naukova Dumka: Kyiv, Ukraine, 2023. [Google Scholar]
- Pellissier, L.; Eidesen, P.B.; Ehrich, D.; Descombes, P.; Schönswetter, P.; Tribsch, A.; Westergaard, K.B.; Alvarez, N.; Guisan, A.; Zimmermann, N.E.; et al. Past climate-driven range shifts and population genetic diversity in arctic plants. J. Biogeogr. 2015, 43, 461–470. [Google Scholar] [CrossRef]
- Wada, N.; Shimmono, M.; Miyamoto, M.; Kojima, S. Warming effects on shoot developmental growth production in sympatric evergreen alpine dwarf shrubs Empetrum nigrum and Loiseleuria procumbens. Ecol. Res. 2002, 17, 125–132. [Google Scholar] [CrossRef]
- Rodríguez-Sanchéz, F.; Arroyo, J. Reconstructing the demise of Tethyan plants: Climate-driven range dynamics of Laurus since the Pliocene. Glob. Ecol. Biogeogr. 2008, 17, 685–695. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Fløjgaard, C.; Marske, K.A.; Nógues-Bravo, D.; Normand, S. Applications of species distribution modelling to paleobiology. Quat. Sci. Rev. 2011, 30, 2930–2947. [Google Scholar] [CrossRef]
- Walas, Ł.; Sobierajska, K.; Ok, T.; Dönmez, A.A.; Kanoğlu, S.S.; Bou Dagher-Kharrat, M.; Douaihy, B.; Romo, A.; Stephan, J.; Jasińska, A.K.; et al. Past, present and future geographic range of the oro-Mediterranean Tertiary relict: Juniperus drupacea case study. Reg. Environ. Change 2019, 19, 1507–1520. [Google Scholar] [CrossRef]
- Zapałowicz, H. Roślinna Szata Gór Pokucko-Marmaroskich; Sprawozdanie Komisyi Fizyjograficznej 24; Akademia Umiejętności: Kraków, Poland, 1889. [Google Scholar]
- Mráz, P.; Ronikier, M. Biogeography of the Carpathians: Evolutionary and spatial facets of biodiversity. Biol. J. Linn. Soc. 2016, 119, 528–559. [Google Scholar] [CrossRef]
- Tasenkevich, L.; Boratyński, A.; Skrypec, K.; Seniv, M.; Khmil, T.; Walas, Ł. Biodiversity of high-mountain woody plants in the East Carpathians in Ukraine. Dendrobiology 2023, 89, 1–19. [Google Scholar] [CrossRef]
- Braun-Blanquet, L. La Végétation Alpine des Pyrénées Olrientales, Étude de phytosociologie Compare; Consejo Superior de Investigaciones Científicas: Barcelona, Spain, 1948. [Google Scholar]
- Villar, L.; Sesé, J.A.; Ferrández, J.V. Atlas de la Flora del Pirineo Aragonés; Consejo de Protección de la Naturaleza de Aragón e Instituto de Estudios Altoaragoneses: Huesca, Spain, 2001; Volume 2. [Google Scholar]
- Gómez García, D.; Ferrández Palacio, J.V.; Tejero, P.; Font, X. Spatial distribution and environmental analysis of the alpine flora in the Pyrenees. Pirineos 2017, 172, e027. [Google Scholar]
- Larcher, W.; Wagner, J. High mountain bioclimate: Temperatures near the ground recorded from the timberline to the nival zone in the Central Alps. Contrib. Nat. Hist. Mus. Bern 2009, 12, 857–874. [Google Scholar]
- Larcher, W.; Wagner, J. Temperatures in the life zones of the Tyrolean Alps. Sitzungsberichte Abteilung 2011, 213, 31–51. [Google Scholar] [CrossRef]
- Wipf, S.; Stoeckli, V.; Bebi, P. Winter climate change in alpine tundra: Plant responses to changes in snow depth and snowmelt timing. Clim. Change 2009, 94, 105–121. [Google Scholar] [CrossRef]
- Ladinig, U.; Hacker, J.; Neuner, G.; Wagner, J. How endangered is sexual reproduction of high-mountain plants by summer frosts? Frost resistance, frequency of frost events and risk assessment. Oecologia 2013, 171, 743–760. [Google Scholar] [CrossRef]
- Tasenkevich, L. Flora of the Carpathians: Checklist of the Native Vascular Plant Species; State Museum of Natural History National Academy of Sciences of Ukraine: Lviv, Ukraine, 1998. [Google Scholar]
- Ninot, J.M.; Carrillo, E.; Font, X.; Carreras, J.; Ferré, A.; Masalles, R.M.; Soriano, I.; Vigo, J. Altitude zonation in the Pyrenees. A geobotanic interpretation. Phytocoenologia 2007, 37, 371–398. [Google Scholar] [CrossRef]
- Ninot, J.M.; Carrillo, E.; Ferré., A. The Pyrenees. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer: Cham, Switzerland, 2017; Volume 12, pp. 323–366. [Google Scholar]
- HabRef v7. Available online: https://inpn.mnhn.fr/habitat/cd_hab/23233/tab/description (accessed on 17 July 2023).
- Baudière, A.; Serve, L. Les Landes rases à Loiseleuria procumbens en Pyrénées Orientales et leur intérêt phytogéographique. Coll. Phytosoc. 1975, 337–347. [Google Scholar]
- Biţă-Nicolae, C.D. The Natural Priority Habitats in the Alpine zone of Bucegi Massif (Romanian Southern Carpathians). Bot. Serbica 2011, 35, 79–85. [Google Scholar]
- Stancu, D.I. The main alpine and subalpine habitats in Râiosu & Buda mountains, Fǎgǎraş massif. Muzeul Olten. Craiova. Oltenia. Stud. Şi Comunicǎri. Ştiinţele Nat. 2013, 29, 118–126. [Google Scholar]
- Puşcaş, M.; Gafta, D.; Cristea, V. Analyse éco-coenotique des prairies édifiées par Carex curvala All. des Carpates Roumaines. Acta Bot. Gall. 2005, 152, 497–505. [Google Scholar] [CrossRef]
- Kricsfalusy, V.V. Mountain grasslands of high conservation value in the Eastern Carpathians: Syntaxonomy, biodiversity, protection and management. Thaiszia 2013, 23, 67–112. [Google Scholar]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. World-wide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634 + maps. [Google Scholar]
- Rivas-Martínez, S.; Penas, Á.; del Rió, S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer: Cham, Switzerland, 2017; Volume 12, pp. 29–80. [Google Scholar]
- Dobrowski, S.Z. A climatic basis for micro-refugia: The influence of terrain on climate. Glob. Change Biol. 2011, 17, 1022–1035. [Google Scholar] [CrossRef]
- Bär, A.; Pape, R.; Bräuning, A.; Löffler, J. Growth-ring variations of dwarf shrubs reflect regional climate signals in alpine environments rather than topoclimatic differences. J. Biogeogr. 2008, 35, 625–636. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.B.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G.; Körner, C.; Thompson, D.B.A. Alpine biodiversity in space and time. In Alpine Biodiversity in Europe; Nagy, L., Grabherr, G., Körner, C., Thompson, D.B.A., Eds.; Springer: Berlin, Germany, 2003; pp. 453–464. [Google Scholar]
- Skrede, I.; Eidesen, P.B.; Piñeiro Portela, R.; Brochmann, C. Refugia, differentiation and postglacial migration in arctic-alpine Eurasia, exemplified by the mountain avens (Dryas octopetala L.). Mol. Ecol. 2006, 15, 1827–1840. [Google Scholar] [CrossRef]
- Stephenson, C.M.; Kohn, D.D.; Park, K.J.; Atkinson, R.; Edwards, C.; Travis, J.M. Testing mechanistic models of seed dispersal for the invasive Rhododendron ponticum (L.). Persp. Pl. Ecol., Evol. Syst. 2007, 9, 15–28. [Google Scholar] [CrossRef]
- Escaravage, N.; Wagner, J. Pollination effectiveness and pollen dispersal in a Rhododendron ferrugineum (Ericaceae) population. Pl. Biol. 2004, 6, 606–615. [Google Scholar] [CrossRef]
- Alsos, I.G.; Alm, T.; Normand, S.; Brochmann, C. Past and future range shifts and loss of diversity in dwarf willow (Salix herbacea L.) inferred from genetics, fossils and modelling. Glob. Ecol. Biogeogr. 2009, 18, 223–239. [Google Scholar] [CrossRef]
- Ronikier, M.; Schneeweiss, G.M.; Schönswetter, P. The extreme disjunction between Beringia and Europe in Ranunculus glacialis s. l. (Ranunculaceae) does not coincide with the deepest genetic split–a story of the importance of temperate mountain ranges in arctic–alpine phylogeography. Mol. Ecol. 2012, 21, 5561–5578. [Google Scholar] [CrossRef]
- Winkler, M.; Tribsch, A.; Schneeweiss, G.M.; Brodbeck, S.; Gugerli, F.; Holderegger, R.; Abbott, R.J.; Schönswetter, P. Tales of the unexpected: Phylogeography of the arctic-alpine model plant Saxifraga oppositifolia (Saxifragaceae) revisited. Mol. Ecol. 2012, 21, 4618–4630. [Google Scholar] [CrossRef]
- Kropf, M.; Comes, H.P.; Kadereit, J.W. Past, present and future of mountain species of the French Massif Central–the case of Soldanella alpina L. subsp. alpina (Primulaceae) and a review of other plant and animal studies. J. Biogeogr. 2012, 39, 799–812. [Google Scholar] [CrossRef]
- Suchan, T.; Malicki, M.; Ronikier, M. Relict populations and Central European glacial refugia: The case of Rhododendron ferrugineum (Ericaceae). J. Biogeogr. 2019, 46, 392–404. [Google Scholar] [CrossRef]
- Lewandowska, A.; Boratyńska, K.; Marcysiak, K.; Gómez, D.; Romo, A.; Malicki, M.; Iszkuło, G.; Boratyński, A. Phenotypic differentiation of Rhododendron ferrugineum populations in European mountains. Dendrobiology 2022, 87, 1–12. [Google Scholar] [CrossRef]
- Lewandowska, A.; Marcysiak, K.; Gómez, D.; Jasińska, A.K.; Romo, A.; Didukh, Y.; Sękiewicz, K.; Boratyńska, K.; Boratyński, A. Inference of taxonomic relationships between Rhododendron ferrugineum and R. myrtifolium (Ericaceae) from leaf and fruit morphologies. Bot. J. Linn. Soc. 2023, 201, 483–497. [Google Scholar] [CrossRef]
- Ikeda, H.; Senni, K.; Fujii, N.; Setoguchi, H. High mountains of the Japanese archipelago as refugia for arctic–alpine plants: Phylogeography of Loiseleuria procumbens (L.) Desvaux (Ericaceae). Biol. J. Linn. Soc. 2009, 97, 403–412. [Google Scholar] [CrossRef]
- Ikeda, H.; Eidesen, P.B.; Yakubov, V.; Barkalov, V.; Brochmann, C.; Setoguchi, H. Late Pleistocene origin of the entire circumarctic range of the arctic-alpine plant Kalmia procumbens. Mol. Ecol. 2017, 26, 5773–5783. [Google Scholar] [CrossRef]
- Ikeda, H.; Carlsen, T.; Fujii, N.; Brochmann, C.; Setoguchi, H. Pleistocene climatic oscillations and the speciation history of an alpine endemic and a widespread arctic-alpine plant. New Phytol. 2012, 194, 583–594. [Google Scholar] [CrossRef]
- Eidesen, P.B.; Ehrich, D.; Bakkestuen, V.; Alsos, I.G.; Gilg, O.; Taberlet, P.; Brochmann, C. Genetic roadmap of the Arctic: Plant dispersal highways, traffic barriers and capitals of diversity. New Phytol. 2013, 200, 898–910. [Google Scholar] [CrossRef]
- Pykälä, J. Relation between extinction and assisted colonization of plants in the arctic-alpine and boreal regions. Conserv. Biol. 2016, 31, 524–530. [Google Scholar] [CrossRef]
- Ronikier, M. Biogeography of high-mountain plants in the Carpathians: An emerging phylogeographical perspective. Taxon 2011, 60, 373–389. [Google Scholar] [CrossRef]
- Hülber, K.; Wessely, J.; Gattringer, A.; Moser, D.; Kuttner, M.; Essl, F.; Leitner, M.; Winkler, M.; Ertl, S.; Willner, W.; et al. Uncertainty in predicting range dynamics of endemic alpine plants under climate warming. Glob. Change Biol. 2016, 22, 2608–2619. [Google Scholar] [CrossRef]
- Maclean, I.M.D.; Hopkins, J.J.; Bennie, J.; Lawson, C.R.; Wilson, R.J. Microclimates buffer plant community response to climate change. Glob. Ecol. Biogeogr. 2015, 24, 1340–1350. [Google Scholar] [CrossRef]
- Treu, R.; Laursen, G.A.; Stephenson, S.L.; Landolt, J.C.; Densmore, R. Mycorrhizae from Denali National Park and Preserve, Alaska. Mycorrhiza 1996, 6, 21–29. [Google Scholar] [CrossRef]
- Koizumi, T.; Nara, K. Communities of putative Ericoid Mycorrhizal Fungi isolated from alpine dwarf shrubs in Japan: Effects of host identity and microhabitat. Microbes Envir. 2017, 32, 147–153. [Google Scholar] [CrossRef]
- Haselwandter, K. Mycorrhizal status of ericaceous plants in alpine and subalpine areas. New Phytol. 1979, 83, 427–431. [Google Scholar] [CrossRef]
- Cripps, C.L.; Eddington, L.H. Distribution of mycorrhizal types among alpine vascular plant families on the Beartooth Plateau, Rocky Mountains, U.S.A., in reference to large-scale patterns in Arctic–Alpine habitats. Arctic Antarct. Alp. Res. 2005, 37, 177–188. [Google Scholar] [CrossRef]
- Schweingruber, F.; Poschlod, P. Growth rings in herbs and shrubs: Lifespan, age determination and stem anatomy. For. Snow Landsc. Res. 2005, 79, 195–415. [Google Scholar]
- Scherrer, D.; Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Zeidler, M.; Banaš, M. Vybrané Kapitoly z Ekologie Horských Ekosystémů; Univerzita Palackého v Olomouci, Přírodovědecká Fakulta: Olomouc, Slovakia, 2013. [Google Scholar]
- García, M.B.; Domingo, D.; Pizarro, M.; Font, X.; Gómez-García, D.; Ehrlén, J. Rocky habitats as microclimatic refuges for biodiversity. A close-up thermal approach. Env. Exp. Bot. 2020, 170, 103886. [Google Scholar] [CrossRef]
- Kuprian, E.; Briceño, V.F.; Wagner, J.; Neuner, G. Ice barriers promote supercooling and prevent frost injury in reproductive buds, flowers and fruits of alpine dwarf shrubs throughout the summer. Environ. Exper. Bot. 2014, 106, 4–12. [Google Scholar] [CrossRef]
- Palacio, S.; Lenz, A.; Wipf, S.; Hoch, G.; Rixen, C. Bud freezing resistance in alpine shrubs across snow depth gradients. Envir. Exper. Bot. 2015, 118, 95–101. [Google Scholar] [CrossRef]
- Tasenkevich, L. Endemism in the Carpathian flora–a chorological aspect. In Geobotanist and Taxonomist. A Volume Dedicated to Professor Adam Zając on the 70th Anniversary of His Birth; Zemanek, B., Ed.; Istitute of Botany of Jagellonian University: Kraków, Poland, 2011; pp. 157–168. [Google Scholar]
- Breman, E.; Hurdu, B.I.; Kliment, J.; Kobiv, Y.; Kučera, J.; Mráz, P.; Pușcaș, M.; Renaud, J.; Ronikier, M.; Šibík, J.; et al. Conserving the endemic flora of the Carpathian Region: An international project to increase and share knowledge of the distribution, evolution and taxonomy of Carpathian endemics and to conserve endangered species. Pl. Syst. Evol. 2020, 306, 15–59. [Google Scholar] [CrossRef]
- Margielewski, W. Geological and geomorphological settings. In Postglacial History of Vegetation in the Polish Part of the Western Carpathians Based on Isopollen Map; Obidowicz, A., Madeyska, E., Turner, C., Eds.; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2013; pp. 11–14. [Google Scholar]
- Mirek, Z. Altitudinal vegetation belts of the Western Carpathians. In Postglacial History of Vegetation in the Polish Part of the Western Carpathians Based on Isopollen Maps; Obidowicz, A., Madeyska, E., Turner, C., Eds.; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2013; pp. 15–21. [Google Scholar]
- Ţopa, E. Ericaceae. In Flora Republicii Popularae Romîne; Sãvulescu, T., Ed.; Academia Republicii Populare Romîne: Bucureşti, Romania, 1960; Volume 7, pp. 119–140. [Google Scholar]
- Jasičová, M. Ericaceae Juss. In Flóra Slovenska; Futák, J., Bertová, L., Eds.; Veda: Bratislava, Slovakia, 1982; Volume 3, pp. 337–359. [Google Scholar]
- Turis, P.; Kliment, J.; Feráková, V.; Dítě, D.; Eliáš, P.; Hrivnák, R.; Košťál, J.; Šuvada, R.; Mráz, P.; Bernátová, D. Red List of vascular plants of the Carpathian part of Slovakia. Thaiszia 2014, 24, 35–87. [Google Scholar]
- Mirek, Z. High Mountain Vascular Plants of the Carpathians. Atlas of Distribution; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2020. [Google Scholar]
- QGIS.org. QGIS Geographic Information System. QGIS Association. 2022. Available online: http://www.qgis.org (accessed on 1 April 2023).
- Bradie, L.; Leung, B. A quantitative synthesis of the importance of variables used in Maxent species distribution models. J. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic predictors for supporting ecological applications in the conterminous United States. U.S. Geol. Surv. Data Ser. 2012, 691, 10. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Intern. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Brown, J.L.; Hill, D.J.; Dolan, A.M.; Carnaval, A.C.; Haywood, A.M. PaleoClim, high spatial resolution paleoclimate surfaces for global land areas. Nat. Sci. Data 2018, 5, 180–254. [Google Scholar] [CrossRef]
- Karger, D.N.; Nobis, M.P.; Normand, S.; Graham, C.H.; Zimmermann, N.E. CHELSA-TraCE21k v1. 0. Downscaled transient temperature and precipitation data since the last glacial maximum. Clim. Past. 2021, 19, 439–456. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; et al. Long-term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 1029–1136. [Google Scholar]
- Otto-Bliesner, B.L.; Marshall, S.J.; Overpeck, J.T.; Miller, G.H.; Hu, A. Simulating arctic climate warmth and ice field retreat in the Last Interglaciation. Science 2006, 311, 1751–1753. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 20 March 2022).
- Phillips, S.J. A Brief Tutor. Maxent. 2017. Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/Maxent_tutorial2017.pdf (accessed on 15 March 2020).
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 15 March 2020).
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Salvà-Catarineu, M.; Romo, A.; Mazur, M.; Zielińska, M.; Minissale, P.; Dönmez, A.A.; Boratyńska, K.; Boratyński, A. Past, present and future geographic range of the relict Mediterranean and Macaronesian Juniperus phoenicea complex. Ecol. Evol. 2021, 11, 5075–5095. [Google Scholar] [CrossRef]
- Wang, Z.; Chang, Y.C.I.; Ying, Z.; Zhu, L.; Yang, Y. A parsimonious threshold-independent protein feature selection method through the area under receiver operating characteristic curve. Bioinformatics 2007, 23, 2788–2794. [Google Scholar] [CrossRef]
- Mas, J.-F.; Soares Filho, B.; Pontius, R.G.; Farfán Gutiérrez, M.; Rodrigues, H. A suite of tools for ROC analysis of spatial models. ISPRS Intern. J. Geo-Inform. 2013, 2, 869–887. [Google Scholar] [CrossRef]
- Didukh, Y. Climate change assessment based on synphytoindication method. In Handbook of Climate Change Mitigation and Adaptation, 3rd ed.; Lackner, M., Sajjadi, B., Chen, W.-Y., Eds.; Springer: Heidelberg, Switzerland, 2022; pp. 2759–2814. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Factor | CPYR | EPYR | PYR | ECARP | SCARP | CARP | |
|---|---|---|---|---|---|---|---|
| AUC | 0.994 | 0.991 | 0.993 | 0.999 | 0.997 | 0.998 | |
| Bio1 | Annual Mean Temperature | 0.1 | 0.1 | 0.1 | 2.0 | 0.3 | 1.3 |
| Bio2 | Mean Diurnal Range | 4.1 | 0.9 | 2.5 | 0.1 | 0.0 | 0.1 |
| Bio3 | Isothermality | 0.6 | 0.2 | 0.4 | 1.7 | 0.0 | 0.9 |
| Bio4 | Temperature Seasonality | 0.3 | 0.2 | 0.3 | 4.7 | 0.0 | 2.4 |
| Bio5 | Max Temperature of Warmest Month | 0.7 | 0.3 | 0.5 | 0.1 | 0.1 | 0.1 |
| Bio6 | Min Temperature of Coldest Month | 0.3 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 |
| Bio7 | Temperature Annual Range | 1.0 | 1.0 | 1.0 | 0.1 | 0.0 | 0.1 |
| Bio8 | Mean Temperature of Wettest Quarter | 0.4 | 2.0 | 1.2 | 0.2 | 0.2 | 0.2 |
| Bio9 | Mean Temperature of Driest Quarter | 3.8 | 6.0 | 4.9 | 5.5 | 8.2 | 6.9 |
| Bio10 | Mean Temperature of Warmest Quarter | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 |
| Bio11 | Mean Temperature of Coldest Quarter | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 |
| Bio12 | Annual Precipitation | 0.6 | 0.2 | 0.4 | 0.1 | 0.0 | 0.1 |
| Bio13 | Precipitation of Wettest Month | 0.1 | 0.3 | 0.2 | 0.0 | 0.0 | 0.0 |
| Bio14 | Precipitation of Driest Month | 1.1 | 0.4 | 0.8 | 0.0 | 0.1 | 0.1 |
| Bio15 | Precipitation Seasonality | 0.3 | 0.1 | 0.2 | 3.4 | 10.0 | 6.7 |
| Bio16 | Precipitation of Wettest Quarter | 0.3 | 1.2 | 0.8 | 0.0 | 0.0 | 0.0 |
| Bio17 | Precipitation of Driest Quarter | 7.8 | 6.2 | 7.0 | 0.1 | 0.0 | 0.1 |
| Bio18 | Precipitation of Warmest Quarter | 0.1 | 0.1 | 0.1 | 31.1 | 9.9 | 20.5 |
| Bio19 | Precipitation of Coldest Quarter | 0.5 | 0.5 | 0.5 | 0.2 | 0.1 | 0.2 |
| Elevation | 77.6 | 80.2 | 78.9 | 50.1 | 70.4 | 60.3 |
| Bioclimatic Factor | CPYR | EPYR | Pyrenees | ECARP | SCARP | Carpathians | |
|---|---|---|---|---|---|---|---|
| bio1 | Annual Mean Temperature | 2.37 | 2.40 | 2.39 | 1.07 | −0.25 | 0.30 |
| bio2 | Mean Diurnal Range | 9.27 | 8.67 | 8.90 | 6.91 | 7.16 | 7.06 |
| bio3 | Isothermality | 33.79 | 32.62 | 33.07 | 27.73 | 29.06 | 28.52 |
| bio4 | Temperature Seasonality | 649.33 | 646.56 | 647.61 | 684.48 | 657.93 | 668.81 |
| bio5 | Max Temperature of Warmest Month | 18.69 | 18.14 | 18.35 | 13.92 | 12.52 | 13.10 |
| bio6 | Min Temperature of Coldest Month | −8.73 | −8.44 | −8.55 | −10.99 | −12.08 | −11.62 |
| bio7 | Temperature Annual Range | 27.43 | 26.58 | 26.90 | 24.91 | 24.60 | 24.73 |
| bio8 | Mean Temperature of Wettest Quarter | −0.62 | −0.30 | −0.42 | 8.98 | 6.84 | 7.73 |
| bio9 | Mean Temperature of Driest Quarter | 10.80 | 10.78 | 10.79 | −6.81 | −8.02 | −7.51 |
| bio10 | Mean Temperature of Warmest Quarter | 10.93 | 10.89 | 10.90 | 9.36 | 7.83 | 8.46 |
| bio11 | Mean Temperature of Coldest Quarter | −4.71 | −4.60 | −4.64 | −7.19 | −8.14 | −7.74 |
| bio12 | Annual Precipitation | 1434.57 | 1447.54 | 1442.62 | 1274.90 | 934.12 | 1075.62 |
| bio13 | Precipitation of Wettest Month | 154.04 | 161.53 | 158.69 | 167.76 | 132.44 | 147.06 |
| bio14 | Precipitation of Driest Month | 75.71 | 68.17 | 71.03 | 65.56 | 46.75 | 54.58 |
| bio15 | Precipitation Seasonality | 20.19 | 23.53 | 22.27 | 32.36 | 39.56 | 36.57 |
| bio16 | Precipitation of Wettest Quarter | 430.82 | 457.02 | 447.08 | 462.96 | 363.02 | 404.52 |
| bio17 | Precipitation of Driest Quarter | 263.48 | 251.75 | 256.20 | 221.51 | 149.49 | 179.33 |
| bio18 | Precipitation of Warmest Quarter | 263.57 | 251.91 | 256.33 | 462.67 | 358.05 | 401.49 |
| bio19 | Precipitation of Coldest Quarter | 388.06 | 399.96 | 395.45 | 228.93 | 149.75 | 182.63 |
| Bioclimatic Factor | CPYR | EPYR | ECARP | SCARP | |||||
|---|---|---|---|---|---|---|---|---|---|
| RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | RCP 2.6 | RCP 8.5 | ||
| AUC | 0.994 | 0.994 | 0.991 | 0.991 | 0.999 | 0.999 | 0.997 | 0.997 | |
| Bio1 | Annual Mean Temperature | 0.0 | 0.1 | 0.0 | 0.0 | 1.5 | 1.1 | 0.6 | 0.4 |
| Bio2 | Mean Diurnal Range | 3.9 | 3.1 | 0.9 | 0.9 | 0.1 | 0.1 | 0.0 | 0.0 |
| Bio3 | Isothermality | 0.8 | 1.1 | 1.7 | 0.1 | 2.1 | 2.7 | 0.0 | 0.0 |
| Bio4 | Temperature Seasonality | 0.3 | 0.7 | 0.2 | 0.2 | 4.2 | 4.9 | 0.0 | 0.1 |
| Bio5 | Max Temperature of Warmest Month | 0.8 | 0.3 | 0.2 | 0.5 | 0.1 | 0.1 | 0.1 | 0.1 |
| Bio6 | Min Temperature of Coldest Month | 0.4 | 0.1 | 0.1 | 0.9 | 0.1 | 0.1 | 0.1 | 0.2 |
| Bio7 | Temperature Annual Range | 0.9 | 0.5 | 0.4 | 1.0 | 0.1 | 0.1 | 0.0 | 0.0 |
| Bio8 | Mean Temperature of Wettest Quarter | 1.2 | 0.7 | 0.4 | 4.0 | 0.0 | 0.1 | 0.1 | 0.2 |
| Bio9 | Mean Temperature of Driest Quarter | 4.3 | 5.5 | 6.4 | 4.0 | 9.5 | 3.8 | 6.6 | 5.9 |
| Bio10 | Mean Temperature of Warmest Quarter | 0.2 | 0.1 | 0.1 | 0.2 | 0.1 | 0.0 | 0.1 | 0.1 |
| Bio11 | Mean Temperature of Coldest Quarter | 0.1 | 0.0 | 0.1 | 0.1 | 0.3 | 0.3 | 0.3 | 0.2 |
| Bio12 | Annual Precipitation | 0.8 | 0.7 | 0.4 | 0.4 | 0.0 | 0.1 | 0.0 | 0.0 |
| Bio13 | Precipitation of Wettest Month | 0.3 | 0.2 | 0.1 | 1.1 | 0.0 | 0.0 | 0.1 | 0.0 |
| Bio14 | Precipitation of Driest Month | 1.0 | 1.0 | 0.4 | 0.5 | 0.0 | 0.1 | 0.0 | 0.1 |
| Bio15 | Precipitation Seasonality | 0.3 | 0.3 | 0.0 | 0.1 | 2.0 | 5.4 | 12.1 | 9.8 |
| Bio16 | Precipitation of Wettest Quarter | 0.5 | 0.3 | 0.2 | 0.3 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bio17 | Precipitation of Driest Quarter | 7.5 | 8.7 | 7.3 | 5.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| Bio18 | Precipitation of Warmest Quarter | 0.1 | 0.1 | 0.2 | 0.2 | 32.0 | 30.3 | 9.6 | 10.2 |
| Bio19 | Precipitation of Coldest Quarter | 0.4 | 0.2 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Elevation | 76.1 | 76.4 | 80.5 | 79.9 | 47.5 | 50.5 | 70.0 | 72.4 | |
| Region | Model | Area in the Probability Levels (Hectares) | Total | |||
|---|---|---|---|---|---|---|
| Low (0.1–0.25) | Medium (0.25–0.50) | High (0.5–0.75) | Very High (>0.75) | |||
| CPYR | Current | 6099.48 | 6545.66 | 4920.93 | 1288.34 | 18,854.41 |
| RCP 2.6 | 6344.43 | 10,857.13 | 420.55 | 16.35 | 17,638.46 | |
| RCP 8.5 | 6698.12 | 711.52 | 4.67 | 0.00 | 7414.31 | |
| EPYR | Current | 4825.15 | 7280.36 | 8192.59 | 1338.57 | 21,636.67 |
| RCP 2.6 | 5756.46 | 5986.58 | 6773.90 | 5156.05 | 23,672.99 | |
| RCP 8.5 | 4482.04 | 4812.61 | 4664.27 | 4470.36 | 18,429.28 | |
| ECARP | Current | 7406.57 | 2796.66 | 1059.53 | 686.48 | 11,949.24 |
| RCP 2.6 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| RCP 8.5 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| SCARP | Current | 9808.72 | 4446.85 | 2123.17 | 708.15 | 17,086.89 |
| RCP 2.6 | 771.79 | 0.00 | 0.00 | 0.00 | 771.79 | |
| RCP 8.5 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walas, Ł.; Pietras, M.; Mazur, M.; Romo, Á.; Tasenkevich, L.; Didukh, Y.; Boratyński, A. The Perspective of Arctic–Alpine Species in Southernmost Localities: The Example of Kalmia procumbens in the Pyrenees and Carpathians. Plants 2023, 12, 3399. https://doi.org/10.3390/plants12193399
Walas Ł, Pietras M, Mazur M, Romo Á, Tasenkevich L, Didukh Y, Boratyński A. The Perspective of Arctic–Alpine Species in Southernmost Localities: The Example of Kalmia procumbens in the Pyrenees and Carpathians. Plants. 2023; 12(19):3399. https://doi.org/10.3390/plants12193399
Chicago/Turabian StyleWalas, Łukasz, Marcin Pietras, Małgorzata Mazur, Ángel Romo, Lydia Tasenkevich, Yakiv Didukh, and Adam Boratyński. 2023. "The Perspective of Arctic–Alpine Species in Southernmost Localities: The Example of Kalmia procumbens in the Pyrenees and Carpathians" Plants 12, no. 19: 3399. https://doi.org/10.3390/plants12193399