
Allelopathic Activity of a Novel Compound, Two Known Sesquiterpenes, and a C13 Nor-Isopenoid from the Leave of Croton oblongifolius Roxb. for Weed Control

Abstract
:1. Introduction
2. Results
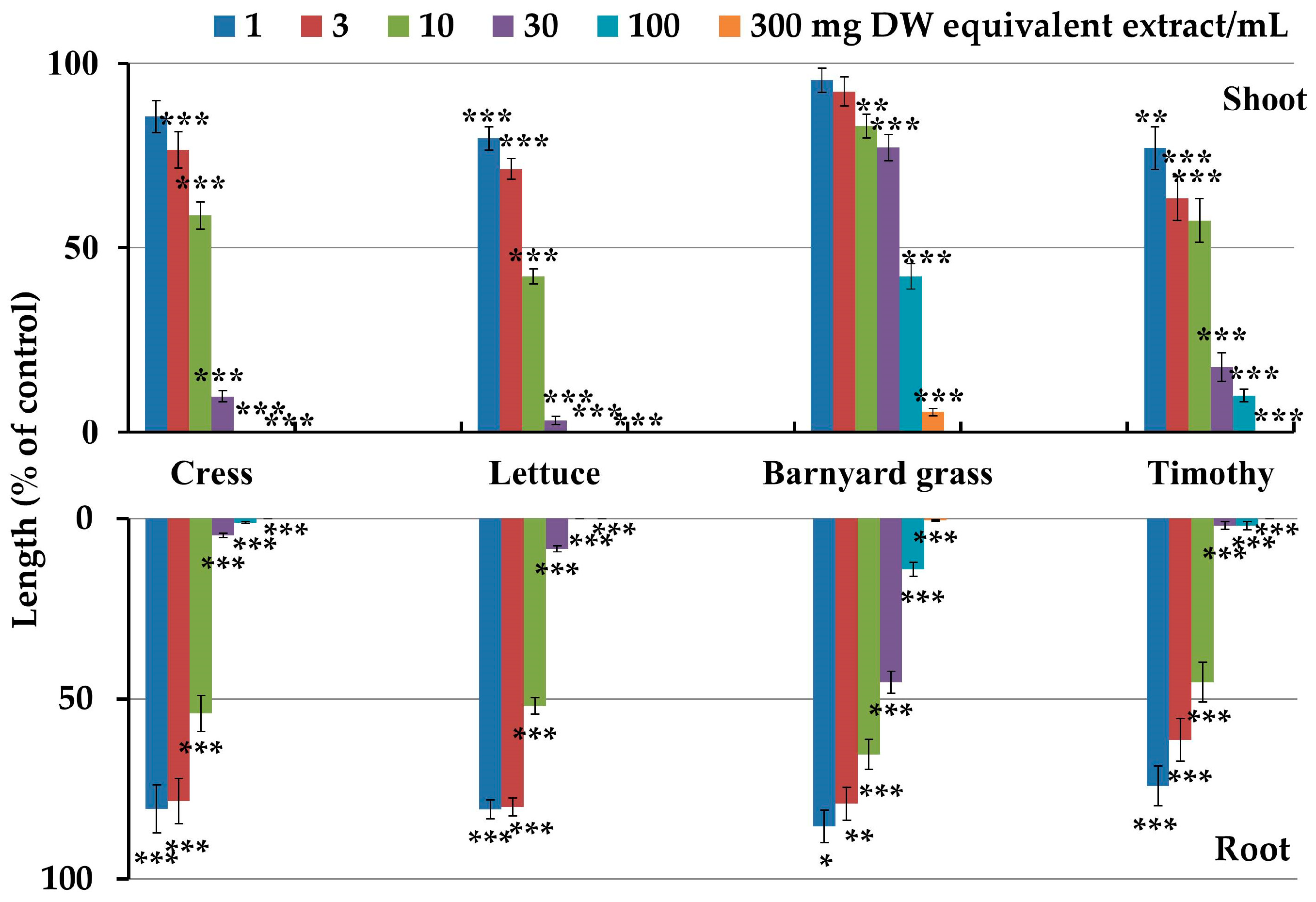
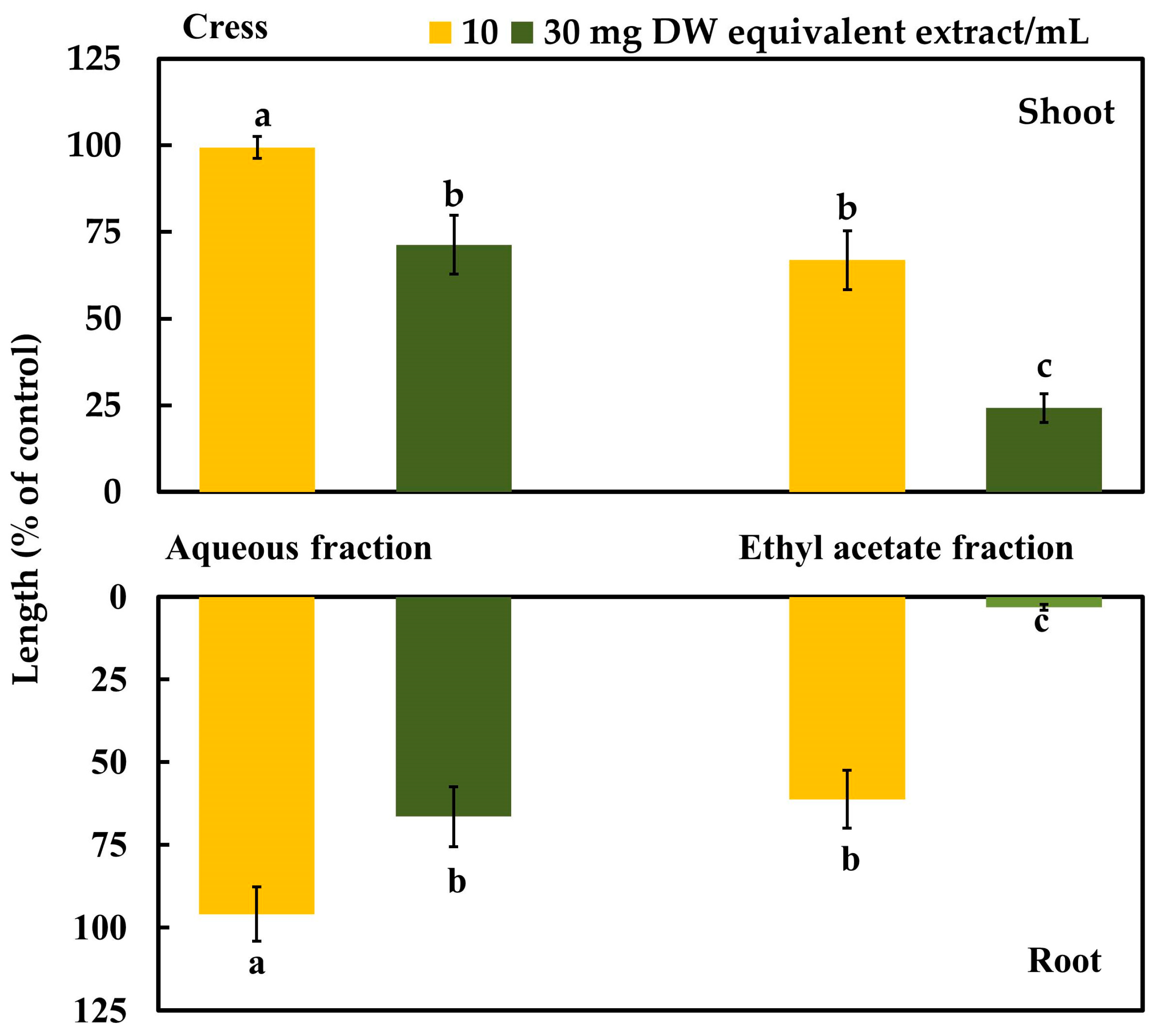
2.1. Inhibitory Activity of Croton oblongifolius
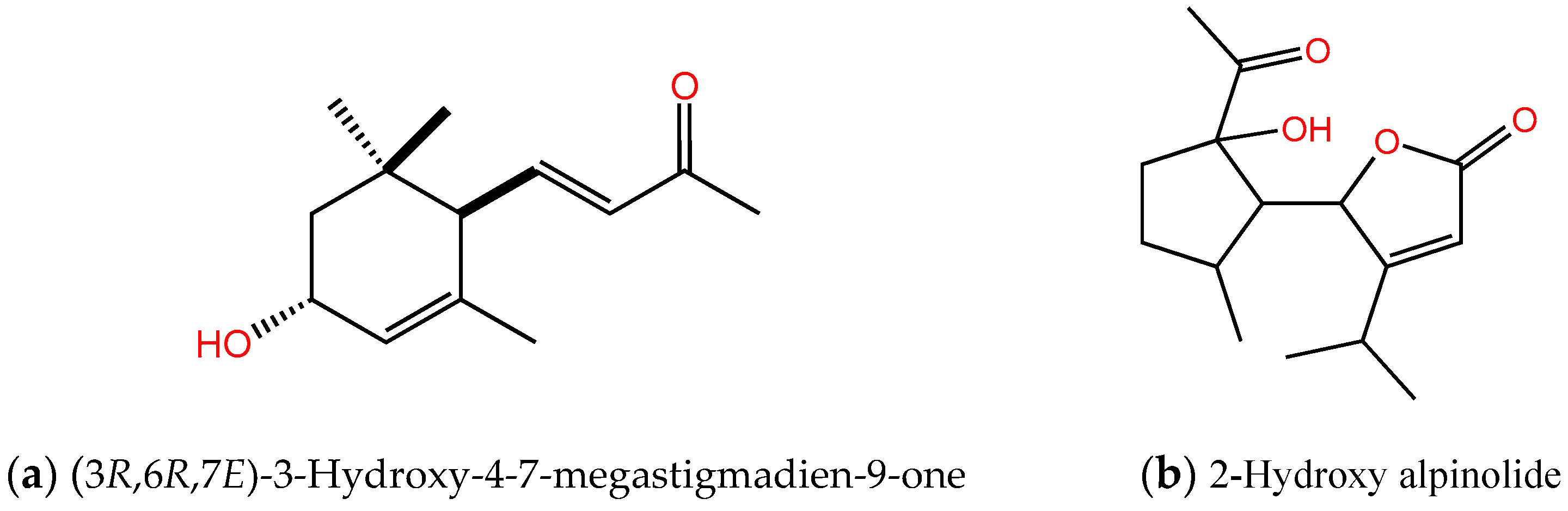
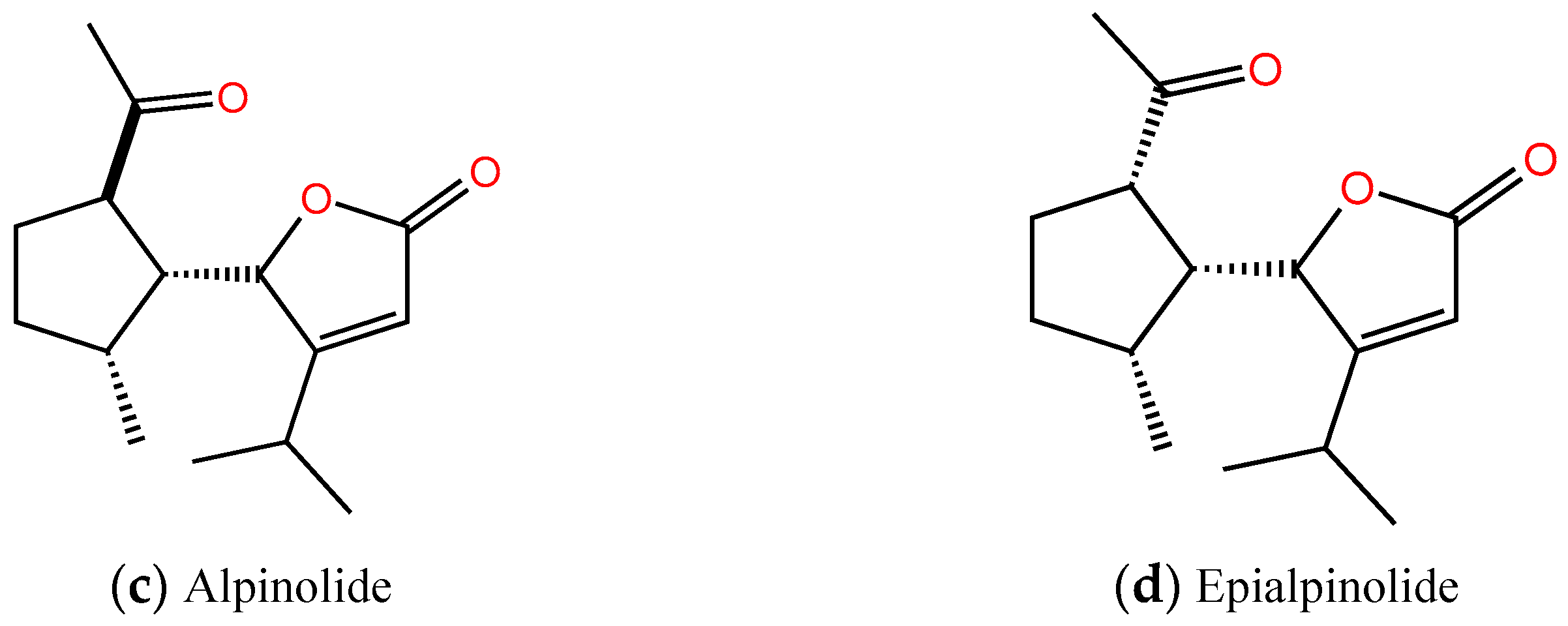
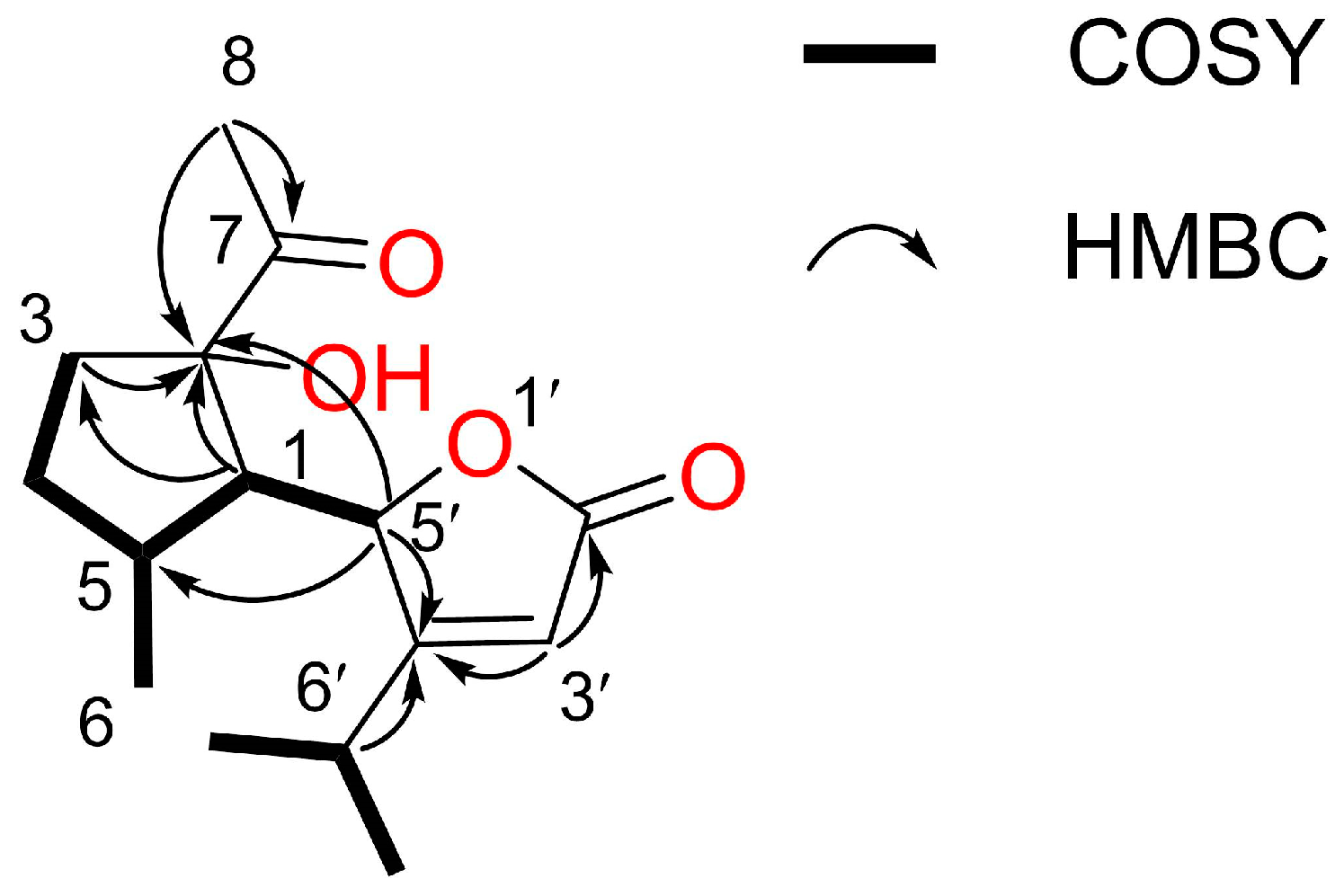
2.2. Identification of the Allelopathic Substances
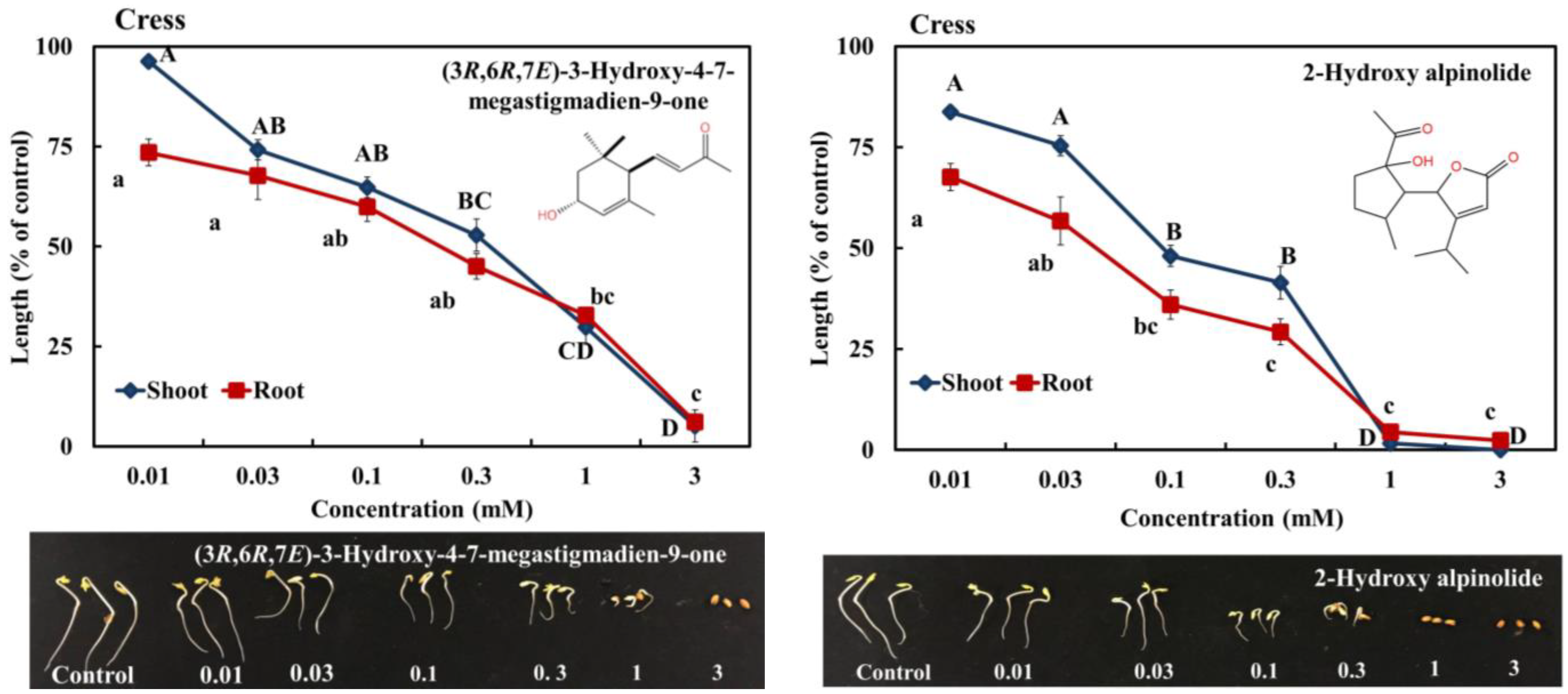
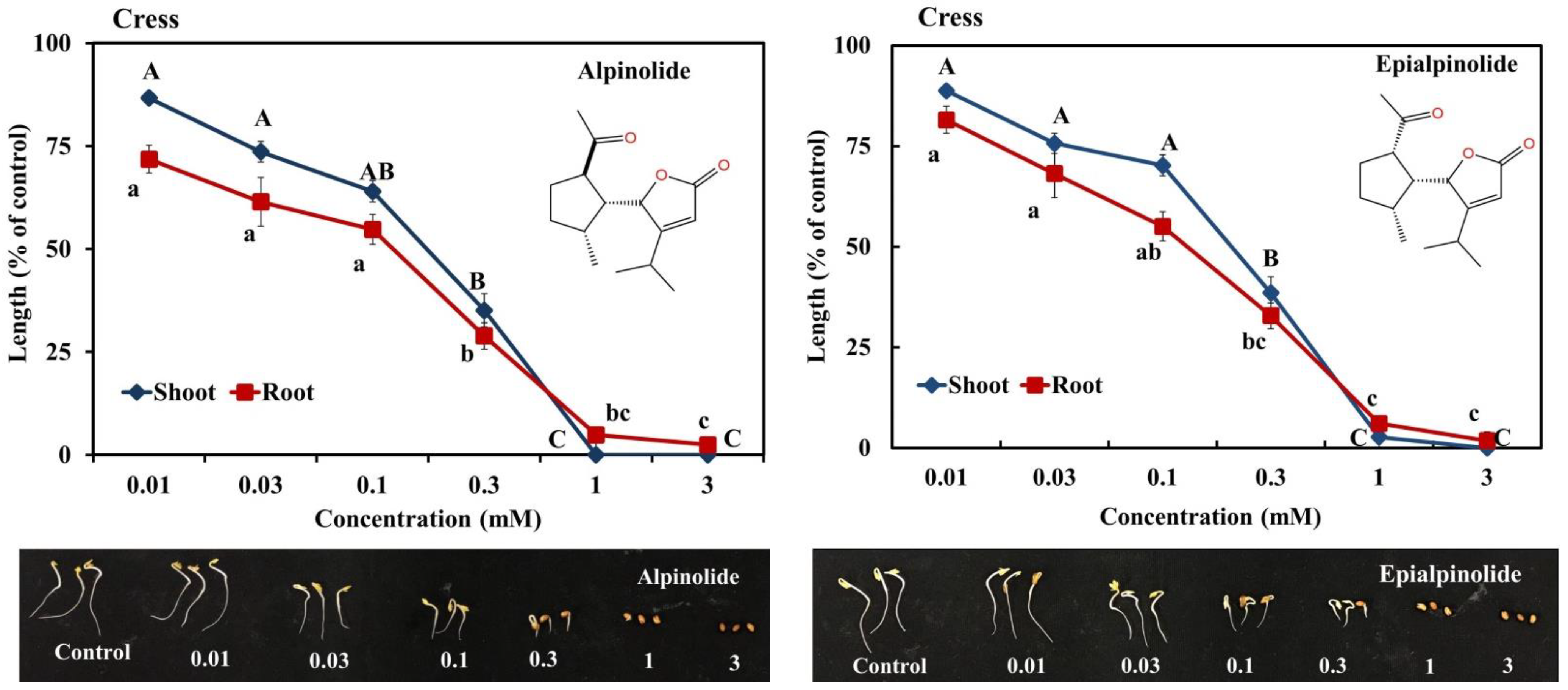
2.3. Allelopathic Activity of Compounds I, II, III, and IV
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Extraction and Bioassay
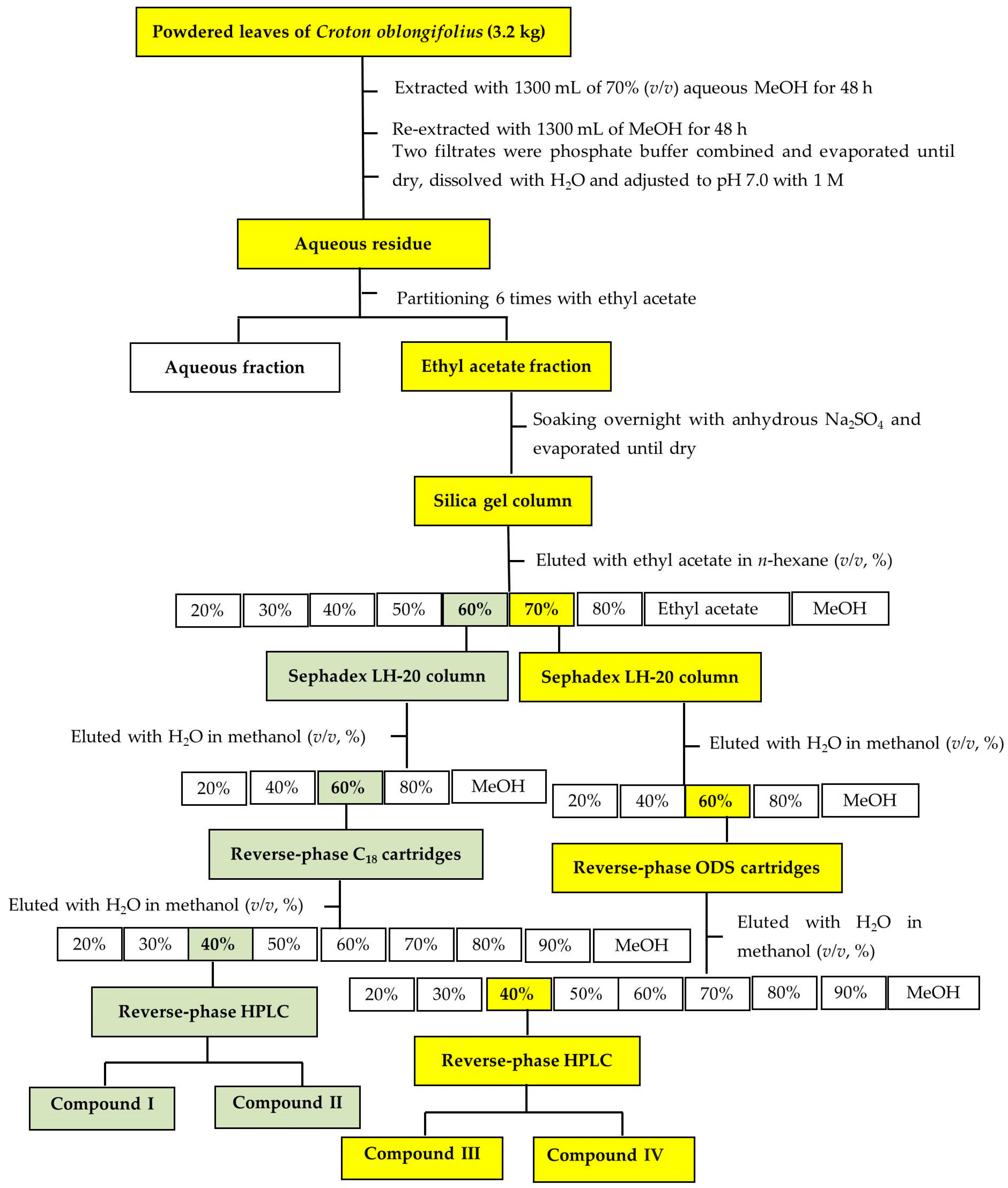
4.3. Isolation and Purification of the Bioactive Substances
4.4. Biological Activity of Compounds I, II, III, and IV
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.G.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, W.; Siddiqui, M.B.; Hakeem, K.R. Allelopathic control of native weeds. In Allelopathy: Springer Briefs in Agriculture; Springer: Cham, Switzerland, 2020; pp. 53–59. [Google Scholar]
- Harker, K.N.; O’Donovan, J.T. Recent weed control, weeds management, and integrated weed management. Weed Technol. 2013, 27, 1–11. [Google Scholar] [CrossRef]
- Wato, T. The role of allelopathy in pest management and crop production-a review. Food Sci. Qual. Manag. 2020, 93, 13–21. [Google Scholar]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Protect. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Abbas, T.; Ahmad, A.; Ahmad Kamal, A.; Nawaz, M.Y.; Jamil, M.A.; Saeed, T.; Abid, M.A.; Ali, H.H.; Ateeq, M. Ways to use allelopathic potential for weed management: A review. Int. J. Food Sci. Agric. 2021, 5, 492–498. [Google Scholar] [CrossRef]
- Iqbal, A.; Shah, F.; Hamayun, M.; Khan, Z.H.; Islam, B.; Rehman, G. Plants are the possible source of allelochemicals that can be useful in promoting sustainable agriculture. Fresenius Environ. Bull. 2019, 28, 1052. [Google Scholar]
- Hussain, M.I.; Reigosa, M.J. Evaluation of herbicide potential of sesquiterpene lactone and flavonoid: Impact on germination, seedling growth indices and root length in Arabidopsis thaliana. Pak. J. Bot. 2014, 46, 995–1000. [Google Scholar]
- Hussain, M.I.; Reigosa, M.J. Evaluation of photosynthetic performance and carbon isotope discrimination in perennial ryegrass (Lolium perenne L.) under allelochemicals stress. Ecotoxicology 2017, 26, 613–624. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ota, K. Biological activities of rice allelochemicals momilactone A and B. Rice Res. 2013, 1, 2. [Google Scholar] [CrossRef]
- Kyaw, E.H.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Assessment of the phytotoxic potential of Dregea volubilis (L.f.) Benth. ex Hook.f. and identification of its phytotoxic substances for weed control. Agriculture 2022, 12, 1826. [Google Scholar] [CrossRef]
- Moh, S.M.; Kurisawa, N.; Suenaga, K.; Kato-Noguchi, H. Allelopathic potential of Marsdenia tenacissima (Roxb.) Moon against four test plants and the biological activity of its allelopathic novel compound, 8-dehydroxy-11β-O-acetyl-12β-O-tigloyl-17β-marsdenin. Plants 2023, 12, 1663. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Salatino, M.L.F.; Negri, G. Traditional uses, chemistry and pharmacology of Croton species (Euphorbiaceae). J. Braz. Chem. Soc. 2007, 18, 11–33. [Google Scholar] [CrossRef]
- Júnior, S.F.P.; Conserva, L.M.; Filho, J.M.B. Clerodane diterpenes from Croton species: Distribution and a compilation of their 13C-NMR spectral data. Nat. Prod. Commun. 2006, 1, 319–344. [Google Scholar] [CrossRef]
- Wu, X.A.; Zhao, Y.M. Advance on chemical composition and pharmacological action of Croton L. Nat. Prod. Res. Dev. 2004, 16, 467–472. [Google Scholar]
- Chakrabarty, T.; Balakrishnan, N.P.; Gupta, D. Trichomes in Indian species of Croton L. (Euphorbiaceae) and their systematic applications. J. Plant Anat. Morphol. 1997, 7, 94–106. [Google Scholar]
- Sommit, D.; Petsom, A.; Ishikawa, T.; Roengsumran, S. Cytotoxic activity of natural labdanes and their semi-synthetic modified derivatives from Croton oblongifolius. Planta Med. 2003, 69, 167–170. [Google Scholar] [CrossRef]
- Ngamrojnavanich, N.; Sirimongkon, S.; Roengsumran, S.; Petsom, A.; Kamimura, H. Inhibition of Na+,K+-ATPase activity by (-)-ent-Kaur-16-en-19-oic acid and its derivatives. Planta Med. 2003, 69, 555–556. [Google Scholar]
- Athikomkulchai, S.; Tadtong, S.; Ruangrungsi, N.; Hongratanaworakit, T. Chemical composition of the essential oils from Croton oblongifolius and its antibacterial activity against Propionibacterium acnes. Nat. Prod. Commun. 2015, 10, 1459–1460. [Google Scholar] [CrossRef]
- Takeshige, Y.; Kawakami, S.; Matsunami, K.; Otsuka, H.; Lhieochaiphant, D.; Lhieochaiphant, S. Oblongionosides A-F, megastigmane glycosides from the leaves of Croton oblongifolius Roxburgh. Phytochemistry 2012, 80, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Pudhom, K.; Vilaivan, T.; Ngamrojanavanich, N.; Dechangvipart, S.; Sommit, D.; Petsom, A.; Roengsumran, S. Furanocembranoids from the stem bark of Croton oblongifolius. J. Nat. Prod. 2007, 70, 659–661. [Google Scholar] [CrossRef]
- Youngsa-ad, W.; Ngamrojanavanich, N.; Mahidol, C.; Ruchirawat, S.; Prawat, H.; Kittakoop, P. Diterpenoids from the roots of Croton oblongifolius. Planta Med. 2007, 73, 1491–1494. [Google Scholar] [CrossRef] [PubMed]
- Roengsumran, S.; Musikul, K.; Petsom, A.; Vilaivan, T.; Sangvanich, P.; Pornpakakul, S.; Puthong, S.; Chaichantipyuth, C.; Jaiboon, N.; Chaichit, N. Croblongifolin, a new anticancer clerodane from Croton oblongifolius. Planta Medica 2002, 68, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Wijesekera, K. A bioactive diterpene; Nasimalun A from Croton oblongifolius Roxb. Prayog. Ras. 2017, 1, 41–44. [Google Scholar] [CrossRef]
- Roengsumran, S.; Petsom, A.; Kuptiyanuwat, N.; Vilaivan, T.; Ngamrojnavanich, N.; Chaichantipyuth, C.; Phuthong, S. Cytotoxic labdane diterpenoids from Croton oblongifolius. Phytochemistry 2001, 56, 103–107. [Google Scholar] [CrossRef]
- Pudhom, K.; Sommit, D. Clerodane diterpenoids and a trisubstituted furan from Croton oblongifolius. Phytochem. Lett. 2011, 4, 147–150. [Google Scholar] [CrossRef]
- Ahmed, B.; Alam, T.; Varshney, M.; Khan, S.A. Hepatoprotective activity of two plants belonging to the Apiaceae and the Euphorbiaceae family. J. Ethnopharmacol. 2002, 79, 313–316. [Google Scholar] [CrossRef]
- Sothearith, Y.; Appiah, K.S.; Mardani, H.; Motobayashi, T.; Suzuki, Y.; Eang Hourt, K.; Sugiyama, A.; Fujii, Y. Determination of the allelopathic potential of Cambodia’s medicinal plants using the dish pack method. Sustainability 2021, 13, 9062. [Google Scholar] [CrossRef]
- D’Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Monaco, P.; Oriano, P.; Temussi, F. Structure elucidation and phytotoxicity of C13 nor-isoprenoids from Cestrum parqui. Phytochemistry 2004, 65, 497–505. [Google Scholar] [CrossRef]
- Itokawa, H.; Morita, H.; Kobayashi, T.; Watanabe, K.; Takase, A.; Iitaka, Y. Two new sesquiterpenoids (alpinolide and hanamyol) from Alpinia Japonica (THUNB.) Miq. Chem. Lett. 1984, 13, 1687–1690. [Google Scholar] [CrossRef]
- Itokawa, H.; Morita, H.; Kobayashi, T.; Watanabe, K.; Iitaka, Y. Novel sesquiterpenes from Alpinia intermedia. Chem. Pharm. Bull. 1987, 35, 2860–2868. [Google Scholar] [CrossRef]
- Moh, S.M.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathic activity of a novel compound, 5,6-dihydrogen-11α-O-acetyl-12β-O-tigloyl-17β-marsdenin, and a known steroidal glycoside from the leaves of Marsdenia tenacissima (Roxb.) Moon. Agronomy 2022, 12, 1536. [Google Scholar] [CrossRef]
- Rob, M.M.; Iwasaki, A.; Suzuki, R.; Suenaga, K.; Kato-Noguchi, H. Garcienone, a novel compound involved in allelopathic activity of Garcinia Xanthochymus Hook. Plants 2019, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Lun, T.L.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of plant-growth inhibitory constituents from Polygonum chinense Linn and evaluation of their bioherbicidal potential. Plants 2023, 12, 1577. [Google Scholar] [CrossRef] [PubMed]
- Bari, I.N.; Kato-Noguchi, H.; Iwasaki, A.; Suenaga, K. Allelopathic potency and an active substance from Anredera cordifolia (Tenore) Steenis. Plants 2019, 8, 134. [Google Scholar] [CrossRef]
- Sun, G.; Putkaradze, N.; Bohnacker, S.; Jonczyk, R.; Fida, T.; Hoffmann, T.; Bernhardt, R.; Härtl, K.; Schwab, W. Six uridine-diphosphate glycosyltransferases catalyze the glycosylation of bioactive C13-apocarotenols. Plant Physiol. 2020, 184, 1744–1761. [Google Scholar] [CrossRef]
- Qi, J.; Shi, R.f.; Yu, J.M.; Li, Y.; Yuan, S.T.; Yang, J.Z.; Hu, J.M.; Ji, A.Q. Chemical constituents from leaves of Camellia nitidissima and their potential cytotoxicity on SGC7901 cells. Chin. Herb. Med. 2016, 8, 80–84. [Google Scholar] [CrossRef]
- Macías, F.A.; Lacret, R.; Varela, R.M.; Nogueiras, C.; Molinillo, J.M.G. Bioactive apocarotenoids from Tectona grandis. Phytochemistry 2008, 69, 2708–2715. [Google Scholar] [CrossRef]
- Hossen, K.; Asato, Y.; Teruya, T.; Kato-Noguchi, H. Identification of four allelopathic compounds including a novel compound from Elaeocarpus floribundus Blume and determination of their allelopathic activity. J. Environ. Manag. 2023, 326, 116728. [Google Scholar] [CrossRef]
- Machida, K.; Kikuchi, M. Norisoprenoids from Viburnum dilatatum. Phytochemistry 1996, 41, 1333–1336. [Google Scholar] [CrossRef]
- Yang, W.; Chen, X.; Li, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in pharmacological activities of terpenoids. Nat. Prod. Commun. 2020, 15, 1–13. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Berchová-Bímová, K.; Šudomová, M.; Malaník, M.; Šmejkal, K.; Rengasamy, K.R.R. In vitro study of multi-therapeutic properties of Thymus bovei Benth. essential oil and its main component for promoting their use in clinical practice. J. Clin Med. 2018, 7, 283. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Bao, R.; Malhi, M. Induction of apoptosis by costunolide in bladder cancer cells is mediated through ROS generation and mitochondrial dysfunction. Molecules 2013, 18, 1418–1433. [Google Scholar] [CrossRef]
- Hsu, J.L.; Pan, S.L.; Ho, Y.F.; Hwang, T.L.; Kung, F.L.; Guh, J.H. Costunolide induces apoptosis through nuclear calcium2+ overload and DNA damage response in human prostate cancer. J. Urol. 2011, 185, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Ortiz, M.P.; Wei, M.Q. Antitumor activity of artemisinin and its derivatives: From a well-known antimalarial agent to a potential anticancer drug. J. Biomed. Biotechnol. 2012, 2012, 247597. [Google Scholar] [CrossRef]
- Macías, F.A.; Galino, J.C.G.; Molinillo, J.M.G.; Castellano, D. Dehydrozaluzanin C: A potent plant growth regulator with potential use as a natural herbicide template. Phytochemistry 2000, 54, 165–171. [Google Scholar] [CrossRef]
- Wiart, C. Medicinal Plants of China, Korea, and Japan: Bioresources for Tomorrow’s Drugs and Cosmetics; Taylor & Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2012; p. 454. ISBN 1439899118. [Google Scholar]
- Kuroyanagi, M.; Ueno, A.; Ujiie, K.; Sato, S. Structures of sesquiterpenes from Curcuma aromatica SALISB. Chem. Pharm. Bull. 1987, 35, 53–59. [Google Scholar] [CrossRef]
- DellaGreca, M.; Fiorentino, A.; Monaco, P.; Previtera, L.; Temussi, F.; Zarrelli, A. New dimeric phenanthrenoids from the rhizomes of Juncus acutus. Structure determination and antialgal activity. Tetrahedron 2003, 59, 2317–2324. [Google Scholar] [CrossRef]
- Okada, Y.; Okajima, H.; Konishi, H.; Terauchi, M.; Ishii, K.; Liu, I.M.; Watanabe, H. Antioxidant effect of naturally occurring furan fatty acids on oxidation of linoleic acid in aqueous dispersion. J Am Oil Chem Soc. 1990, 67, 858–862. [Google Scholar] [CrossRef]
- Wang, X.; Gao, W.; Yao, Z. Immunosuppressive sesquiterpenes from Tripterygium wilfordii. Chem. Pharm. Bull. 2005, 53, 607–610. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | I50 Value (mg DW Equivalent Extract/mL) | Correlation Coefficient (r) | |||
|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | ||
| Dicots | Cress | 9.4 c | 8.3 c | −0.83 *** | −0.81 *** |
| Lettuce | 5.8 de | 8.2 c | −0.89 *** | −0.91 *** | |
| Monocots | Barnyard grass | 65.7 a | 18.2 b | −0.72 *** | −0.77 *** |
| Timothy | 7.6 cd | 4.8 e | −0.71 *** | −0.74 *** | |
| Position | δH Mult (J in Hz) a | δC b |
|---|---|---|
| 1 | 2.36, dd (7.5, 3.8) | 56.5 |
| 2 | 88.7 | |
| 3a | 2.47, m | 36.9 |
| 3b | 1.66, ddd (13.7, 9.0, 4.9) | |
| 4a | 2.03, m | 32.6 |
| 4b | 1.51, m | |
| 5 | 2.48, m | 36.1 |
| 6 | 0.85, d (7.3) | 16.0 |
| 7 | 214.1 | |
| 8 | 2.35, s | 26.9 |
| 2’ | 175.7 | |
| 3’ | 5.87, t (1.6) | 115.1 |
| 4’ | 182.4 | |
| 5’ | 5.41, dd (3.8, 1.8) | 83.1 |
| 6’ | 2.71, m | 28.6 |
| 7’ | 1.16, d (7.0) | 22.2 |
| 8’ | 1.28, d (6.6) | 20.3 |
| Tested Plant | I50 Value (mM) | ||||
|---|---|---|---|---|---|
| Compound I | Compound II | Compound III | Compound IV | ||
| Cress | Shoot | 0.24 a | 0.11 c | 0.12 c | 0.16 b |
| Root | 0.15 b | 0.04 e | 0.07 d | 0.09 cd | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moh, S.M.; Tojo, S.; Teruya, T.; Kato-Noguchi, H. Allelopathic Activity of a Novel Compound, Two Known Sesquiterpenes, and a C13 Nor-Isopenoid from the Leave of Croton oblongifolius Roxb. for Weed Control. Plants 2023, 12, 3384. https://doi.org/10.3390/plants12193384
Moh SM, Tojo S, Teruya T, Kato-Noguchi H. Allelopathic Activity of a Novel Compound, Two Known Sesquiterpenes, and a C13 Nor-Isopenoid from the Leave of Croton oblongifolius Roxb. for Weed Control. Plants. 2023; 12(19):3384. https://doi.org/10.3390/plants12193384
Chicago/Turabian StyleMoh, Seinn Moh, Shunya Tojo, Toshiaki Teruya, and Hisashi Kato-Noguchi. 2023. "Allelopathic Activity of a Novel Compound, Two Known Sesquiterpenes, and a C13 Nor-Isopenoid from the Leave of Croton oblongifolius Roxb. for Weed Control" Plants 12, no. 19: 3384. https://doi.org/10.3390/plants12193384