

Physiological and Cellular Ultrastructural Responses of Sesuvium portulacastrum under Cd Stress Grown Hydroponically


Abstract
:1. Introduction
2. Results
2.1. Effect of Cd Stress on Chlorophyll Content of Leaves
2.2. Effect of Cd Stress on Lipid Peroxidation of Tissues
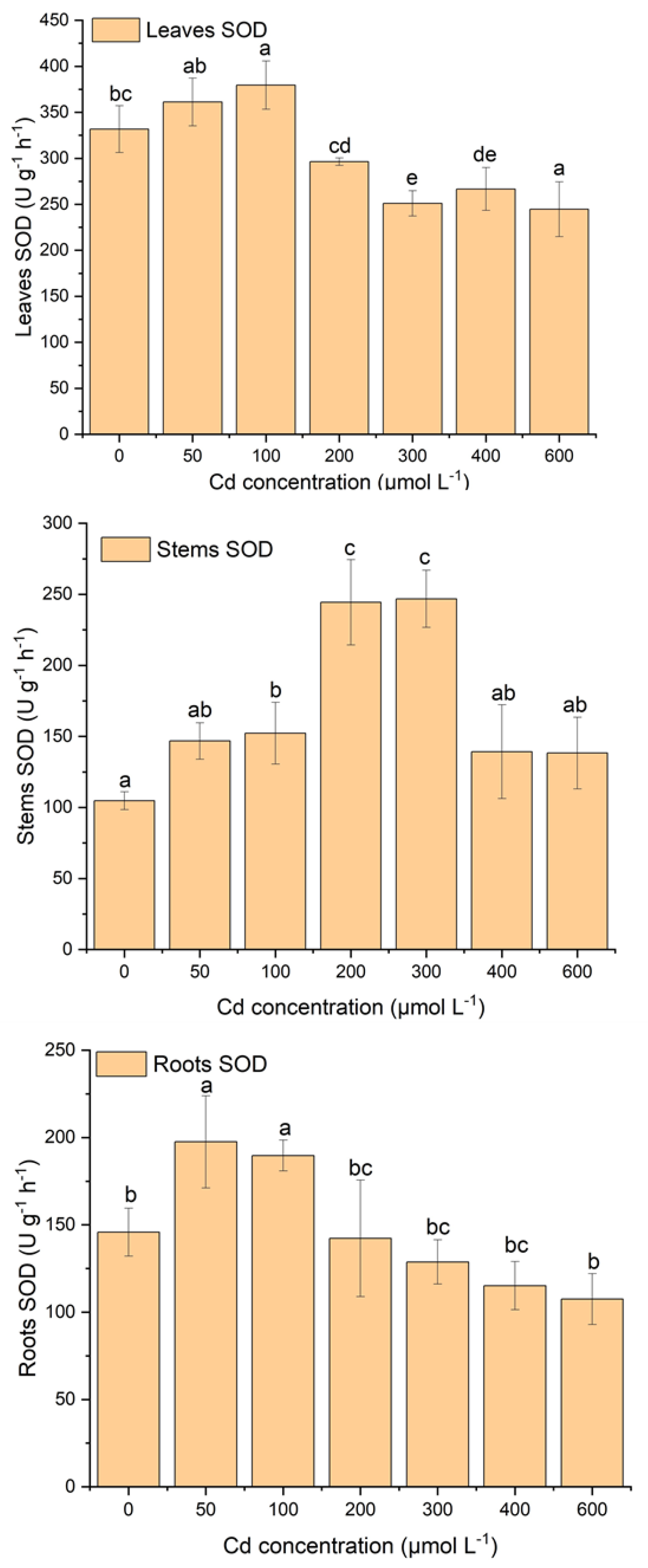
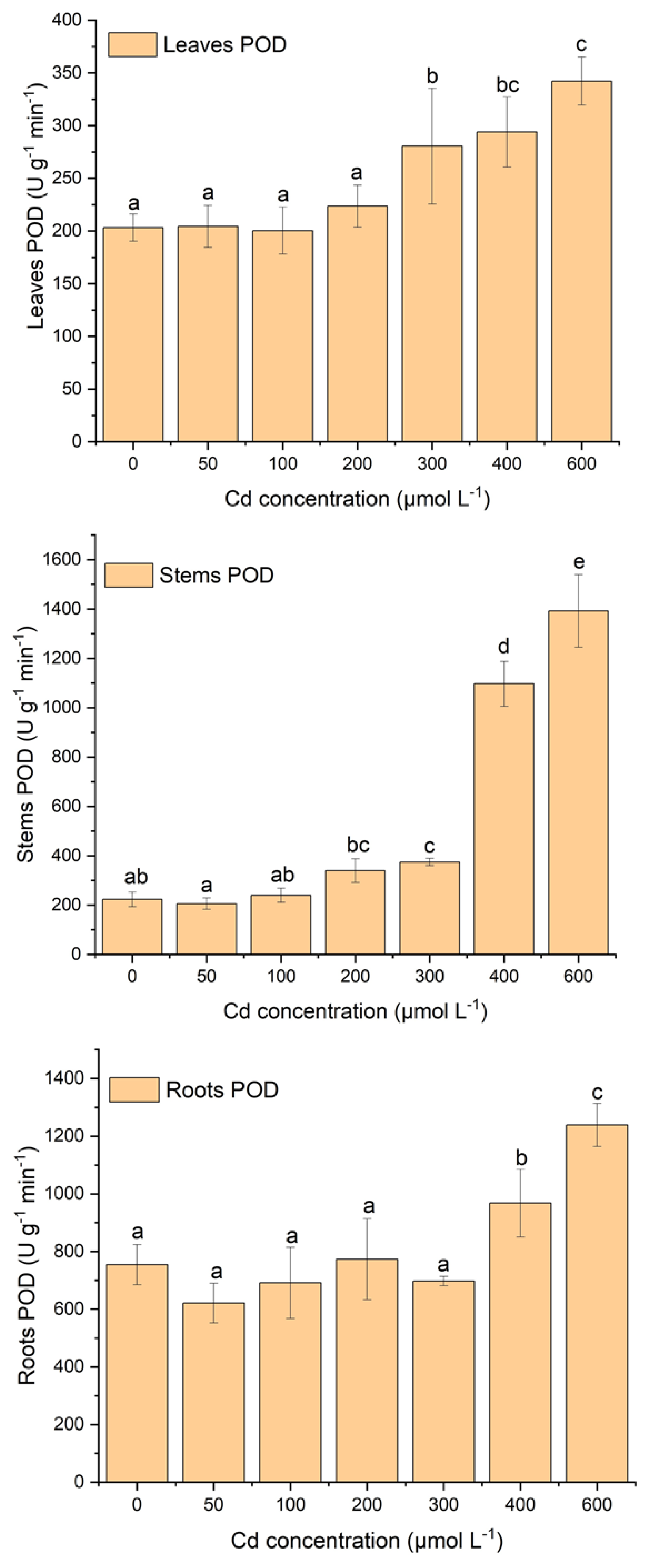
2.3. Effect of Cd Stress on Antioxidant System of S. portulacastrum
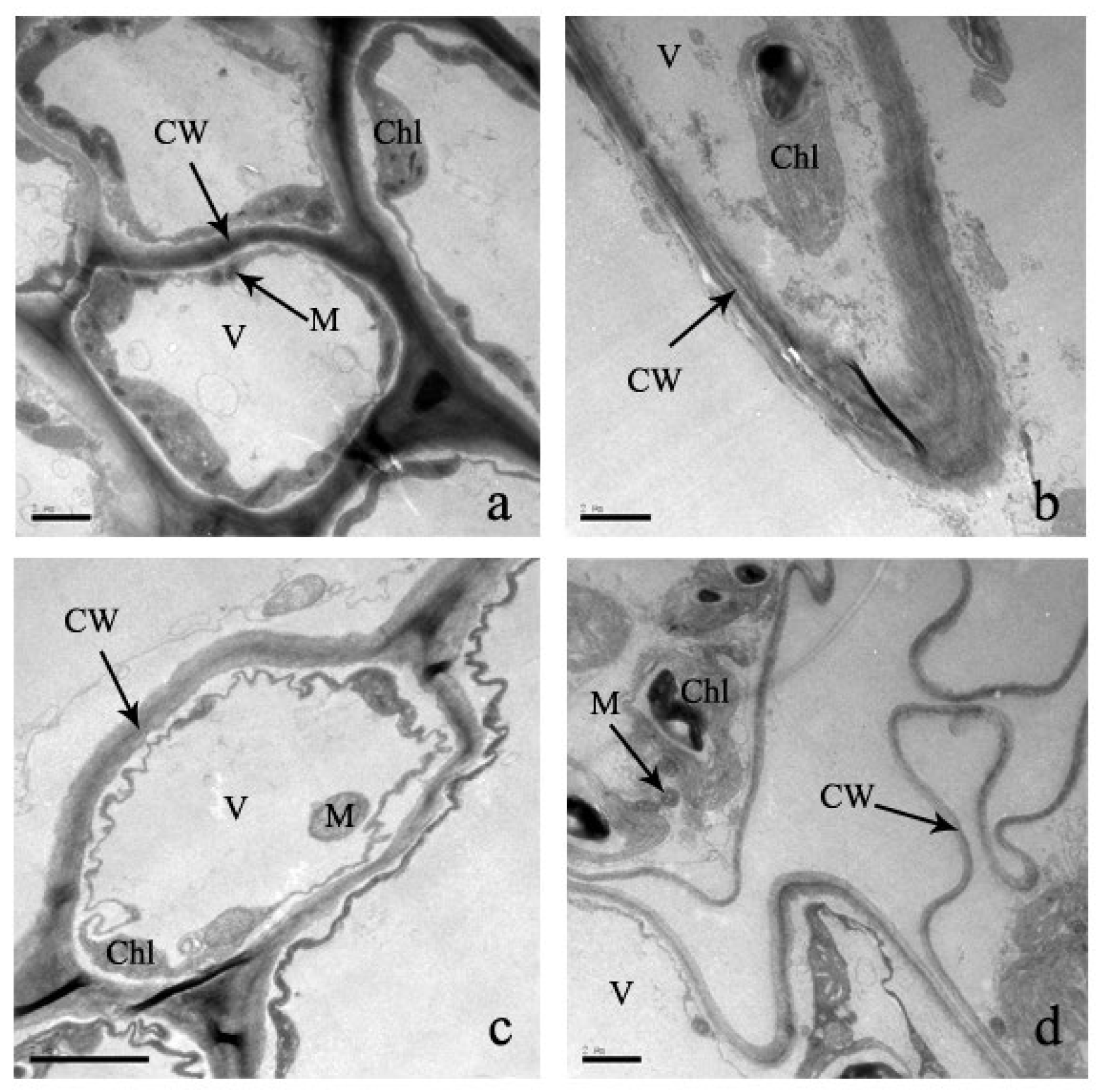
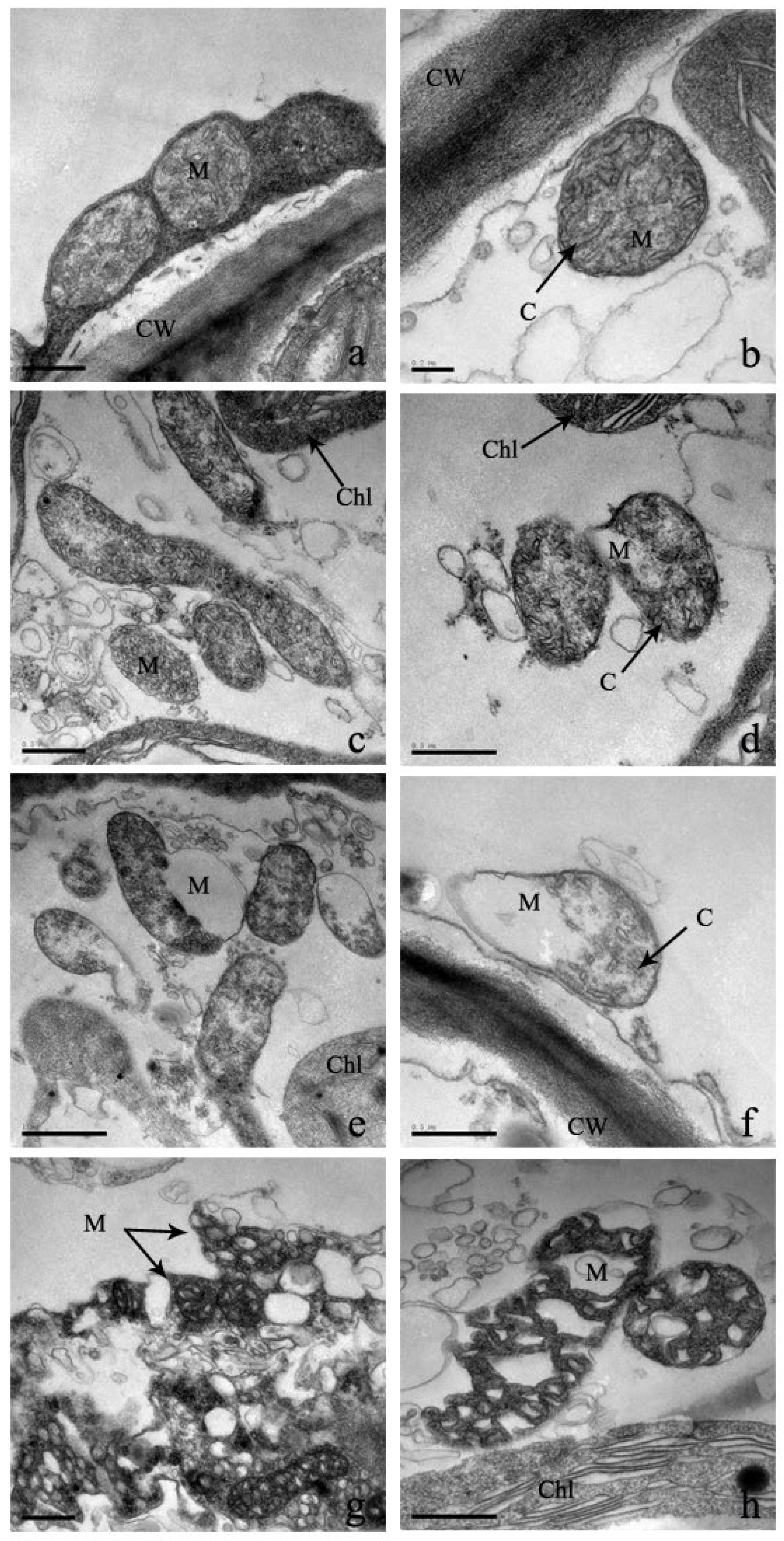
2.4. Effect of Cd Stress on Ultrastructure of S. portulacastrum Cells
3. Discussion
3.1. Effect of Cadmium on Chlorophyll Content
3.2. Effect of Cadmium Stress on Lipid Peroxidation and Antioxidant Enzymes
3.3. Effect of Cadmium Stress on Ultrastructure of Cells
4. Materials and Methods
4.1. Experimental Plant and Treatments Protocol
4.2. Determination of Chlorophyll Content in Leaves of S. portulacastrum
4.3. Determination of MDA Content in Tissues of S. portulacastrum
4.4. Determination of the Activities of Antioxidant Enzymes in S. portulacastrum
4.5. Determination of Cell Structures and Their Deformations of S. portulacastrum
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.-L.; Xu, X.-R.; Sun, Y.-X.; Liu, J.-L.; Li, H.-B. Heavy Metal Pollution in Coastal Areas of South China: A Review. Mar. Pollut. Bull. 2013, 76, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Huang, L.; Bin, X. Spatial and Temporal Changes of Heavy Metals in Coastal Mangrove Sediment in China: Review of Present Status. J. Biol. Sci. 2019, 19, 314–322. [Google Scholar] [CrossRef]
- Uddin, M.M.; Peng, G.; Wang, Y.; Huang, J.; Huang, L. Pollution Status, Spatial Distribution and Ecological Risk of Heavy Metals in Sediments of a Drinking Water Lake in South Eastern China. Environ. Pollut. Bioavailab. 2021, 33, 19–30. [Google Scholar] [CrossRef]
- Pachés, M.; Martínez-Guijarro, R.; Romero, I.; Aguado, D. Assessment of Metal Pollution and Its Environmental Impact on Spanish Mediterranean Coastal Ecosystems. J. Mar. Sci. Eng. 2023, 11, 89. [Google Scholar] [CrossRef]
- Uddin, M.M.; Huang, L. Temporal Distribution, Accumulation, Speciation and Ecological Risk of Heavy Metals in the Sediment of an Urban Lagoon Catchment at Xiamen in China. Chem. Ecol. 2022, 38, 801–822. [Google Scholar] [CrossRef]
- Uddin, M.M.; Huang, C.; Chen, X.; Huang, L. Speciation and Mobility of Heavy Metals in the Yundang Lagoon Catchment Surface Sediments after 30 Years of Continuous Remediation and Management at Xiamen in China. Chem. Ecol. 2021, 38, 48–71. [Google Scholar] [CrossRef]
- Uddin, M.M.; Xie, B.; Peng, G.; Huang, L. Heavy Metal Pollution Status, Spatial Distribution and Associated Ecological Risks within Sediments of Yundang Lagoon Catchment in Xiamen, China, after 30 Years Continuous Ecological Rehabilitation and Management. Hum. Ecol. Risk Assess. Int. J. 2020, 27, 465–482. [Google Scholar] [CrossRef]
- Uddin, M.M.; Huang, L. Influence of Mangrove Forestation on Heavy Metals Accumulation and Speciation in Sediments and Phytoremediation Capacity of Mangrove Species of an Artificial Managed Coastal Lagoon at Xiamen in China. Chem. Ecol. 2022, 39, 1–23. [Google Scholar] [CrossRef]
- Uddin, M.M.; Peng, G.; Huang, L. Trophic Transfer, Bioaccumulation, and Potential Health Risk of Trace Elements in Water and Aquatic Organisms of Yundang Lagoon at Xiamen in China. Toxin Rev. 2022, 42, 242–256. [Google Scholar] [CrossRef]
- Sun, J.X. A brief overview of the national ecological environment quality in 2020. Environ. Econ. 2021, 7, 8–9. [Google Scholar]
- Yang, G.L.; Zheng, M.M.; Liao, H.M.; Tan, A.J.; Feng, D.; Lv, S.M. Influence of Cadmium and Microplastics on Physiological Responses, Ultrastructure and Rhizosphere Microbial Community of Duckweed. Ecotoxicol. Environ. Saf. 2022, 243, 114011. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, X.; Hou, L.; Ji, S.; Zhang, Y.; Fan, W.; Li, T.; Zhang, L.; Liu, P.; Yang, L. Effects of Cadmium on Transcription, Physiology, and Ultrastructure of Two Tobacco Cultivars. Sci. Total Environ. 2023, 869, 161751. [Google Scholar] [CrossRef]
- Appenroth, K.-J. What Are “Heavy Metals” in Plant Sciences? Acta Physiol. Plant. 2010, 32, 615–619. [Google Scholar] [CrossRef]
- Daud, M.K.; Ali, S.; Variath, M.T.; Zhu, S.J. Differential Physiological, Ultramorphological and Metabolic Responses of Cotton Cultivars under Cadmium Stress. Chemosphere 2013, 93, 2593–2602. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.d.R.H.; Nowroz, F.; Nahar, K. Insight into the Physiological and Biochemical Mechanisms of Biostimulating Effect of Ascophyllum nodosum and Moringa oleifera Extracts to Minimize Cadmium-Induced Oxidative Stress in Rice. Environ. Sci. Pollut. Res. 2023, 30, 55298–55313. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Chen, L. Growth and Physiological Responses of Pennisetum Sp. to Cadmium Stress under Three Different Soils. Environ. Sci. Pollut. Res. 2020, 28, 14867–14881. [Google Scholar] [CrossRef]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of Cadmium Stress on Growth and Physiological Characteristics of Sassafras Seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef]
- Shackira, A.M.; Puthur, J.T. Enhanced Phytostabilization of Cadmium by a Halophyte—Acanthus ilicifolius L. Int. J. Phytoremediation 2016, 19, 319–326. [Google Scholar] [CrossRef]
- Bankaji, I.; Caçador, I.; Sleimi, N. Physiological and Biochemical Responses of Suaeda fruticosa to Cadmium and Copper Stresses: Growth, Nutrient Uptake, Antioxidant Enzymes, Phytochelatin, and Glutathione Levels. Environ. Sci. Pollut. Res. 2015, 22, 13058–13069. [Google Scholar] [CrossRef]
- Jiang, N.; Li, Z.; Yang, J.; Zu, Y. Responses of Antioxidant Enzymes and Key Resistant Substances in Perennial Ryegrass (Lolium perenne L.) to Cadmium and Arsenic Stresses. BMC Plant Biol. 2022, 22, 145. [Google Scholar] [CrossRef]
- Muradoglu, F.; Gundogdu, M.; Ercisli, S.; Encu, T.; Balta, F.; Jaafar, H.Z.; Zia-Ul-Haq, M. Cadmium Toxicity Affects Chlorophyll a and b Content, Antioxidant Enzyme Activities and Mineral Nutrient Accumulation in Strawberry. Biol. Res. 2015, 48, 11. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, A.; Conti, G.O.; Jho, E.H.; Zuccarello, P.; Grasso, A.; Copat, C.; Ferrante, M. Phytoremediation of Contaminated Soils by Heavy Metals and PAHs. A Brief Review. Environ. Technol. Innov. 2017, 8, 309–326. [Google Scholar] [CrossRef]
- Uddin, M.M.; Chen, Z.; Huang, L. Cadmium Accumulation, Subcellular Distribution and Chemical Fractionation in Hydroponically Grown Sesuvium portulacastrum [Aizoaceae]. PLoS ONE 2020, 15, e0244085. [Google Scholar] [CrossRef] [PubMed]
- Ghnaya, T.; Nouairi, I.; Slama, I.; Messedi, D.; Grignon, C.; Abdelly, C.; Ghorbel, M.H. Cadmium Effects on Growth and Mineral Nutrition of Two Halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef]
- Ghnaya, T.; Slama, I.; Messedi, D.; Grignon, C.; Ghorbel, M.H.; Abdelly, C. Effects of Cd2+ on K+, Ca2+ and N Uptake in Two Halophytes Sesuvium portulacastrum and Mesembryanthemum crystallinum: Consequences on Growth. Chemosphere 2007, 67, 72–79. [Google Scholar] [CrossRef]
- Wali, M.; Fourati, E.; Hmaeid, N.; Ghabriche, R.; Poschenrieder, C.; Abdelly, C.; Ghnaya, T. NaCl Alleviates Cd Toxicity by Changing Its Chemical Forms of Accumulation in the Halophyte Sesuvium portulacastrum. Environ. Sci. Pollut. Res. 2015, 22, 10769–10777. [Google Scholar] [CrossRef]
- Mariem, W.; Kilani, B.R.; Benet, G.; Abdelbasset, L.; Stanley, L.; Charlotte, P.; Chedly, A.; Tahar, G. How Does NaCl Improve Tolerance to Cadmium in the Halophyte Sesuvium portulacastrum? Chemosphere 2014, 117, 243–250. [Google Scholar] [CrossRef]
- Fourati, E.; Vogel-Mikuš, K.; Bettaieb, T.; Kavčič, A.; Kelemen, M.; Vavpetič, P.; Pelicon, P.; Abdelly, C.; Ghnaya, T. Physiological Response and Mineral Elements Accumulation Pattern in Sesuvium portulacastrum L. Subjected in Vitro to Nickel. Chemosphere 2019, 219, 463–471. [Google Scholar] [CrossRef]
- Wali, M.; Gunsè, B.; Llugany, M.; Corrales, I.; Abdelly, C.; Poschenrieder, C.; Ghnaya, T. High Salinity Helps the Halophyte Sesuvium portulacastrum in Defense against Cd Toxicity by Maintaining Redox Balance and Photosynthesis. Planta 2016, 244, 333–346. [Google Scholar] [CrossRef]
- Bouzidi, A.; Chaieb, M.; Ellouzi, H.; Krouma, A. Physiological Studies on Sulla Carnosa Growth, Ionic Compartmentation and Oxidative Stress under Salt Stress. Russ. J. Plant Physiol. 2022, 69, 1–10. [Google Scholar] [CrossRef]
- Hou, Q.; Ye, G.; Wang, R.; Jia, L.; Liang, J.; Feng, H.; Wen, J.; Shi, D.; Wang, Q. Changes by Cadmium Stress in Lipid Peroxidation and Activities of Lipoxygenase and Antioxidant Enzymes in Arabidopsis Are Associated with Extracellular ATP. Biologia 2017, 72, 1467–1474. [Google Scholar] [CrossRef]
- Wang, J.; Ge, J.; Yang, X.; Cheng, D.; Yuan, C.; Liu, Z.; Yang, S.; Guo, Y.; Gu, Y. Distribution and Ecological Risk Assessment of Heavy Metals in Sediments of Dajiuhu Lake Wetland in Shennongjia, China. Environ. Sci. Pollut. Res. 2022, 30, 25999–26011. [Google Scholar] [CrossRef]
- Xue, Z.-C.; Gao, H.-Y.; Zhang, L.-T. Effects of Cadmium on Growth, Photosynthetic Rate and Chlorophyll Content in Leaves of Soybean Seedlings. Biol. Plant. 2013, 57, 587–590. [Google Scholar] [CrossRef]
- Yang, H.Y.; Shi, G.X.; Xu, Q.S.; Wang, H.X. Cadmium Effects on Mineral Nutrition and Stress in Potamogeton Crispus. Russ. J. Plant Physiol. 2011, 58, 253–260. [Google Scholar] [CrossRef]
- Gill, S.S.; Khan, N.A.; Tuteja, N. Cadmium at High Dose Perturbs Growth, Photosynthesis and Nitrogen Metabolism While at Low Dose It up Regulates Sulfur Assimilation and antioxidant machinery in garden cress (Lepidium sativum L.). Plant Sci. 2012, 182, 112–120. [Google Scholar] [CrossRef]
- Tunçtürk, M.; Rezaee Danesh, Y.; Tunçtürk, R.; Oral, E.; Najafi, S.; Nohutçu, L.; Jalal, A.; da Silva Oliveira, C.E.; Filho, M.C.M.T. Safflower (Carthamus tinctorius L.) Response to Cadmium Stress: Morpho-Physiological Traits and Mineral Concentrations. Life 2023, 13, 135. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Keutgen, A.J.; Keutgen, N.; Sierka, E.; Baczewska-Dąbrowska, A.H.; Mojski, J.; Pawluśkiewicz, B.; Sieczko, L.; Kalaji, H.M. Photosynthetic Efficiency of Perennial Ryegrass (Lolium perenne L.) Seedlings in Response to Ni and Cd Stress. Sci. Rep. 2023, 13, 5357. [Google Scholar] [CrossRef]
- Liu, C.; Xiao, R.; Dai, W.; Huang, F.; Yang, X. Cadmium Accumulation and Physiological Response of Amaranthus tricolor L. under Soil and Atmospheric Stresses. Environ. Sci. Pollut. Res. 2020, 28, 14041–14053. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of Enzymatic and Nonenzymatic Antioxidants in Plants during Abiotic Stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Mishra, S.; Srivastava, S.; Tripathi, R.D.; Dwivedi, S.; Shukla, M.K. Response of Antioxidant Enzymes in Coontail (Ceratophyllum demersum L.) Plants under Cadmium Stress. Environ. Toxicol. 2008, 23, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Farheen, J.; Mansoor, S. Morpho-Biochemical Response of Vigna radiata to Salinity Generated Hydrogen Peroxide Stress. Pak. J. Bot. 2020, 52, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Nali, C.; Lorenzini, G.; Massai, R.; Landi, M. Living in a Mediterranean City in 2050: Broadleaf or Evergreen ‘Citizens’? Environ. Sci. Pollut. Res. 2017, 25, 8161–8173. [Google Scholar] [CrossRef] [PubMed]
- Marchica, A.; Loré, S.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Remorini, D. Early Detection of Sage (Salvia officinalis L) Responses to Ozone Using Reflectance Spectroscopy. Plants 2019, 8, 346. [Google Scholar] [CrossRef]
- Merey, H.A.; Ramadan, N.K.; Diab, S.S.; Moustafa, A.A. Validated UPLC Method for the Determination of Guaiphenesin, Oxeladin Citrate, Diphenhydramine, and Sodium Benzoate in Their Quaternary Mixture Used in Treatment of Cough, in the Presence of Guaiphenesin-Related Substance (Guaiacol). Chem. Pap. 2018, 72, 2247–2254. [Google Scholar] [CrossRef]
- Zheng, D.F. Effects of Plant Growth Regulators (PGRs) on Endogenous Hormone Contents and Activities of Protective Enzymes in Soybean Leaves. Acta Agron. Sin. 2008, 34, 1233–1239. [Google Scholar] [CrossRef]
- Shaw, B.P. Effects of Mercury and Cadmium on the Activities of Antioxidative Enzymes in the Seedlings of Phaseolus aureus. Biol. Plant. 1995, 37, 587–596. [Google Scholar] [CrossRef]
- Amari, T.; Souid, A.; Ghabriche, R.; Porrini, M.; Lutts, S.; Sacchi, G.A.; Abdelly, C.; Ghnaya, T. Why Does the Halophyte Mesembryanthemum crystallinum Better Tolerate Ni Toxicity than Brassica juncea: Implication of Antioxidant Defense Systems. Plants 2020, 9, 312. [Google Scholar] [CrossRef]
- Spormann, S.; Nadais, P.; Sousa, F.; Pinto, M.; Martins, M.; Sousa, B.; Fidalgo, F.; Soares, C. Accumulation of Proline in Plants under Contaminated Soils—Are We on the Same Page? Antioxidants 2023, 12, 666. [Google Scholar] [CrossRef]
- Najeeb, U.; Jilani, G.; Ali, S.; Sarwar, M.; Xu, L.; Zhou, W. Insights into Cadmium Induced Physiological and Ultra-Structural Disorders in Juncus effusus L. and Its Remediation through Exogenous Citric Acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, X.; Li, K.; Wu, M.; Zhang, R.; Zhang, L.; Chen, G. Photosynthetic Responses of Oryza sativa L. Seedlings to Cadmium Stress: Physiological, Biochemical and Ultrastructural Analyses. Biometals 2014, 27, 389–401. [Google Scholar] [CrossRef]
- Guo, H.; Hong, C.; Chen, X.; Xu, Y.; Liu, Y.; Jiang, D.; Zheng, B. Different Growth and Physiological Responses to Cadmium of the Three Miscanthus Species. PLoS ONE 2016, 11, e0153475. [Google Scholar] [CrossRef]
- Mocquot, B.; Vangronsveld, J.; Clijsters, H.; Mench, M. Copper Toxicity in Young Maize (Zea mays L.) Plants: Effects on Growth, Mineral and Chlorophyll Contents, and Enzyme Activities. Plant Soil 1996, 182, 287–300. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Bloom, P.R. Extinction Coefficients of Chlorophyll a and b in N,N-Dimethylformamide and 80% Acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, C.; Bai, S.; Sun, Z.; Zhu, T.; Zhu, X.; Grit, D.H.; Tembrock, L.R. Cadmium Accumulation and Tolerance of Lagerstroemia indica and Lagerstroemia fauriei (Lythraceae) Seedlings for Phytoremediation Applications. Int. J. Phytoremediation 2016, 18, 1104–1112. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Chai, M.W.; Shi, F.C.; Li, R.L.; Liu, F.C.; Qiu, G.Y.; Liu, L.M. Effect of NaCl on Growth and Cd Accumulation of Halophyte Spartina alterniflora under CdCl2 Stress. South Afr. J. Bot. 2013, 85, 63–69. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A Spectrophotometric Method for Measuring the Breakdown of Hydrogen Peroxide by Catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cd Concentration (μM) | Chl a (mg g−1) | Chl b (mg g−1) | Chl(a + b) (mg g−1) | Chl a/b |
|---|---|---|---|---|
| 0 | 6.52 ± 0.57 | 2.05 ± 0.17 | 8.56 ± 0.74 | 3.18 ± 0.06 |
| 50 | 5.93 ± 0.37 | 1.99 ± 0.01 | 7.92 ± 0.38 | 2.98 ± 0.16 |
| 100 | 5.07 ± 0.52 | 1.78 ± 0.13 | 6.85 ± 0.65 | 2.84 ± 0.10 |
| 200 | 4.62 ± 0.22 | 1.61 ± 0.17 | 6.23 ± 0.39 | 2.88 ± 0.16 |
| 300 | 3.94 ± 0.20 | 1.45 ± 0.00 | 5.39 ± 0.20 | 2.81 ± 0.19 |
| 400 | 2.24 ± 0.50 | 1.11 ± 0.11 | 3.35 ± 0.61 | 1.95 ± 0.53 |
| 600 | 1.19 ± 0.63 | 0.90 ± 0.15 | 2.10 ± 0.78 | 1.27 ± 0.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, M.M.; Chen, Z.; Xu, F.; Huang, L. Physiological and Cellular Ultrastructural Responses of Sesuvium portulacastrum under Cd Stress Grown Hydroponically. Plants 2023, 12, 3381. https://doi.org/10.3390/plants12193381
Uddin MM, Chen Z, Xu F, Huang L. Physiological and Cellular Ultrastructural Responses of Sesuvium portulacastrum under Cd Stress Grown Hydroponically. Plants. 2023; 12(19):3381. https://doi.org/10.3390/plants12193381
Chicago/Turabian StyleUddin, Mohammad Mazbah, Zhenfang Chen, Fuliu Xu, and Lingfeng Huang. 2023. "Physiological and Cellular Ultrastructural Responses of Sesuvium portulacastrum under Cd Stress Grown Hydroponically" Plants 12, no. 19: 3381. https://doi.org/10.3390/plants12193381