
Adaptive Relationships in Hemi-Boreal Forests: Tree Species Responses to Competition, Stress, and Disturbance



Abstract
:1. Introduction
2. Methods
2.1. Study Area, Climate, and Forest Zone
2.2. Reviewing Hemi-Boreal Forest Characteristics
3. Results and Discussion
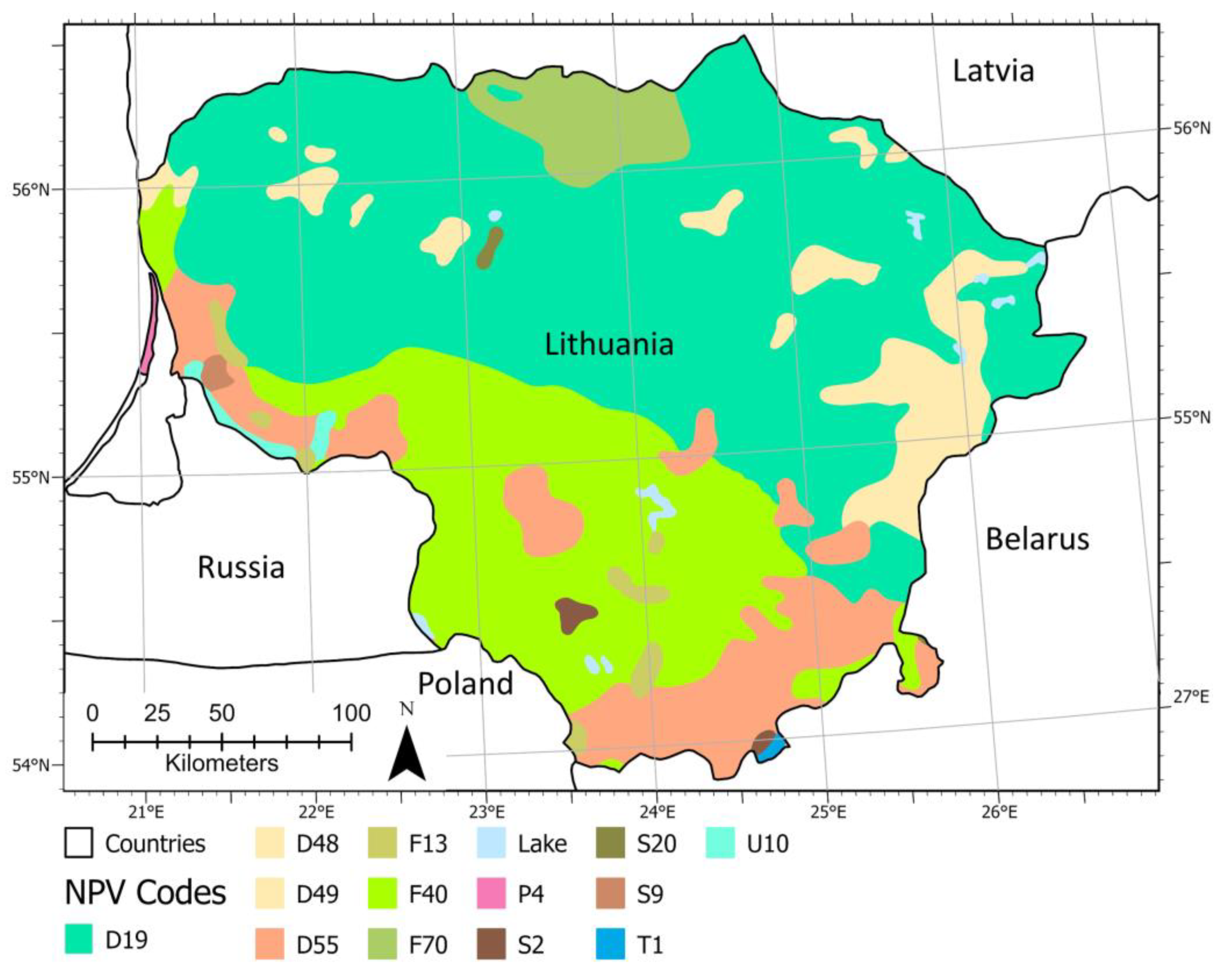
3.1. Potential Natural Forest Communities
3.2. Successional Characteristics of Hemi-Boreal Forest Communities: A Background
3.3. Adaptive Strategies of Forest Tree Species: A Conceptualization
3.4. A Synthesis: Adaptive Relationships in Hemi-Boreal Tree Communities
4. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filotas, E.; Parrott, L.; Burton, P.J.; Chazdon, R.L.; Coates, K.D.; Coll, L.; Haeussler, S.; Martin, K.; Nocentini, S.; Puettmann, K.J.; et al. Viewing Forests through the Lens of Complex Systems Science. Ecosphere 2014, 5, art1. [Google Scholar] [CrossRef]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-Boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, J.; Angelstam, P.; Elmqvist, T.; Emanuelsson, U.; Folke, C.; Ihse, M.; Moberg, F.; Nyström, M. Reserves, Resilience and Dynamic Landscapes. AMBIO J. Hum. Environ. 2003, 32, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Kotar, J. Approaches to Ecologically Based Forest Management on Private Lands; USDA Forest Service, Minnesota Extension Service, University of Minnesota: St. Paul, MM, USA, 1997. [Google Scholar]
- Kuuluvainen, T. Conceptual Models of Forest Dynamics in Environmental Education and Management: Keep It as Simple as Possible, but No Simpler. For. Ecosyst. 2016, 3, 18. [Google Scholar] [CrossRef]
- Whisenant, S. Managing and Directing Natural Succession. In Forest Restoration in Landscapes: Beyond Planting Trees; Mansourian, S., Vallauri, D., Dudley, N., Eds.; Springer: New York, NY, USA, 2005; pp. 257–261. ISBN 978-0-387-29112-3. [Google Scholar]
- Bukantis, A.; Ignatavičius, G.; Satkūnas, J.; Sinkevičius, S.; Šulijienė, G.; Vasarevičius, S.; Veteikis, D. Lithuania’s Environment: State, Processes and Trends; Kopa: Vilnius, Lietuva, 2013; ISBN 978-9955-772-58-3. [Google Scholar]
- Naumov, V.; Manton, M.; Elbakidze, M.; Rendenieks, Z.; Priednieks, J.; Uhlianets, S.; Yamelynets, T.; Zhivotov, A.; Angelstam, P. How to Reconcile Wood Production and Biodiversity Conservation? The Pan-European Boreal Forest History Gradient as an “Experiment”. J. Environ. Manag. 2018, 218, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Roberge, J.-M.; Angelstam, P.; Villard, M.-A. Specialised Woodpeckers and Naturalness in Hemiboreal Forests—Deriving Quantitative Targets for Conservation Planning. Biol. Conserv. 2008, 141, 997–1012. [Google Scholar] [CrossRef]
- Petrokas, R. Appropriate Measures for Retention Forestry. Balt. For. 2016, 22, 382–389. [Google Scholar]
- Richards, P.W. The Tropical Rain Forest: An Ecological Study; University Press: Cambridge, UK, 1952. [Google Scholar]
- Lefèvre, F.; Boivin, T.; Bontemps, A.; Courbet, F.; Davi, H.; Durand-Gillmann, M.; Fady, B.; Gauzere, J.; Gidoin, C.; Karam, M.-J.; et al. Considering Evolutionary Processes in Adaptive Forestry. Ann. For. Sci. 2014, 71, 723–739. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance Legacies Increase the Resilience of Forest Ecosystem Structure, Composition, and Functioning. Ecol. Appl. 2014, 24, 2063–2077. [Google Scholar] [CrossRef]
- Angelstam, P.; Kuuluvainen, T. Boreal Forest Disturbance Regimes, Successional Dynamics and Landscape Structures: A European Perspective. Ecol. Bull. 2004, 51, 117–136. [Google Scholar] [CrossRef]
- Petrokas, R.; Kavaliauskas, D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land 2022, 11, 1249. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Angelstam, P.; Frelich, L.; Jõgiste, K.; Koivula, M.; Kubota, Y.; Lafleur, B.; Macdonald, E. Natural Disturbance-Based Forest Management: Moving Beyond Retention and Continuous-Cover Forestry. Front. For. Glob. Chang. 2021, 4, 629020. [Google Scholar] [CrossRef]
- Ibanga, D.-A. Is Deep Ecology Inapplicable in African Context: A Conversation with Fainos Mangena. Filos. Theor. J. Afr. Philos. Cult. Relig. 2017, 6, 101–119. [Google Scholar] [CrossRef]
- European Commission Guidelines on Closer-to-Nature Forest Management 2023. Available online: https://environment.ec.europa.eu/publications/guidelines-closer-nature-forest-management_en (accessed on 17 August 2023).
- Spies, T.A.; Hessburg, P.F.; Skinner, C.N.; Puettmann, K.J.; Reilly, M.J.; Davis, R.J.; Kertis, J.A.; Long, J.W.; Shaw, D.C. Chapter 3: Old Growth, Disturbance, Forest Succession, and Management in the Area of the Northwest Forest Plan. In Synthesis of Science to Inform Land Management within the Northwest Forest Plan Area; Tech. Coords. Gen. Tech. Rep. PNW-GTR-966; Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., Eds.; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2018; Volume 966, pp. 95–243. [Google Scholar]
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Angelstam, P.; Asplund, B.; Bastian, O.; Engelmark, O.; Fedoriak, M.; Grunewald, K.; Ibisch, P.L.; Lindvall, P.; Manton, M.; Nilsson, M.; et al. Tradition as Asset or Burden for Transitions from Forests as Cropping Systems to Multifunctional Forest Landscapes: Sweden as a Case Study. For. Ecol. Manag. 2022, 505, 119895. [Google Scholar] [CrossRef]
- Jõgiste, K.; Korjus, H.; Stanturf, J.A.; Frelich, L.E.; Baders, E.; Donis, J.; Jansons, A.; Kangur, A.; Köster, K.; Laarmann, D.; et al. Hemiboreal Forest: Natural Disturbances and the Importance of Ecosystem Legacies to Management. Ecosphere 2017, 8, e01706. [Google Scholar] [CrossRef]
- Bohn, U.; Gollub, G.; Hettwer, C.; Weber, H.; Neuhäuslová, Z.; Raus, T.; Schlüter, H. Karte Der Natürlichen Vegetation Europas/Map of the Natural Vegetation of—Maßstab/Scale 1:2,500,000; Federal Agency for Nature Conservation: Bonn, Germany, 2000. [Google Scholar]
- Mozgeris, G.; Kazanavičiūtė, V.; Juknelienė, D. Does Aiming for Long-Term Non-Decreasing Flow of Timber Secure Carbon Accumulation: A Lithuanian Forestry Case. Sustainability 2021, 13, 2778. [Google Scholar] [CrossRef]
- Manton, M.; Ruffner, C.; Kibirkštis, G.; Brazaitis, G.; Marozas, V.; Pukienė, R.; Makrickiene, E.; Angelstam, P. Fire Occurrence in Hemi-Boreal Forests: Exploring Natural and Cultural Scots Pine Fire Regimes Using Dendrochronology in Lithuania. Land 2022, 11, 260. [Google Scholar] [CrossRef]
- Bartašius, L. A Message from the Forest—“We Each Own the Forest”. Live Stories—Labanoras. Available online: https://www.youtube.com/watch?v=LH-3I9D3J6M (accessed on 26 March 2023).
- Kuliešis, A.; Kulbokas, G.; Kasperavičius, A.; Kazanavičiūtė, V.; Kvalkauskienė, M. Lithuanian National Forest Inventory, 1998–2017. From Measurements to Decision Making; Lututė: Kaunas, Lithuania, 2021; ISBN 978-9955-37-234-9. [Google Scholar]
- Karazija, S. Forest Types of Lithuania; Mokslas: Vilnius, Lithuania, 1988; ISBN 978-5-420-00421-0. [Google Scholar]
- Vaičys, M.; Labanauskas, B. Dirvožemio tipologinės grupės [Groups of site classification]. In Miškininko Taksasinis Žinynas [Manual of Forest Inventory]; Mintis: Vilnius, Lithuania, 1968; pp. 182–211. [Google Scholar]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Barbati, A.; Corona, P.; Marchetti, M. European Forest Types. Categories and Types for Sustainable Forest Management and Reporting; EEA Technical report; European Environment Agency: Copenhagen, Denmark, 2006; ISBN 92-9167-886-4. [Google Scholar]
- Vaičys, M.; Mažvila, J. The Influence of Soil Characteristics on Plant Productivity and Ecological Stability. Ekologija 2009, 55, 99–106. [Google Scholar] [CrossRef]
- Buivydaitė, V. Classification of Soils of Lithuania Based on FAO-Unesco Soil Classification System and WRB. In Proceedings of the 17 World Congress of Soil Science, Bangkok, Thailand, 14–20 August 2002; pp. 2189-1–2189-13. [Google Scholar]
- Armolaitis, K.; Varnagirytė-Kabašinskienė, I.; Žemaitis, P.; Stakėnas, V.; Beniušis, R.; Kulbokas, G.; Urbaitis, G. Evaluation of Organic Carbon Stocks in Mineral and Organic Soils in Lithuania. Soil Use Manag. 2022, 38, 355–368. [Google Scholar] [CrossRef]
- IUSS Working Group. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; ISBN 979-8-9862451-1-9. [Google Scholar]
- Petrokas, R.; Ibanga, D.-A.; Manton, M. Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests. Diversity 2022, 14, 892. [Google Scholar] [CrossRef]
- Anyomi, K.A.; Neary, B.; Chen, J.; Mayor, S.J. A Critical Review of Successional Dynamics in Boreal Forests of North America. Environ. Rev. 2022, 30, 563–594. [Google Scholar] [CrossRef]
- Cequinel, A.; Capellesso, E.S.; Marcilio-Silva, V.; Cardoso, F.C.G.; Marques, M.C.M. Determinism in Tree Turnover during the Succession of a Tropical Forest. Perspect. Plant Ecol. Evol. Syst. 2018, 34, 120–128. [Google Scholar] [CrossRef]
- Taylor, A.R.J. Concepts, Theories and Models of Succession in the Boreal Forest of Central Canada. Ph.D. Thesis, Lakehead University, Thunder Bay, ON, Canada, 2009. [Google Scholar]
- Chang, C.; HilleRisLambers, J. Integrating Succession and Community Assembly Perspectives. F1000Research 2016, 5, 2294. [Google Scholar] [CrossRef] [PubMed]
- Petrokas, R. Forest Climax Phenomenon: An Invariance of Scale. Forests 2020, 11, 56. [Google Scholar] [CrossRef]
- Putz, F.E.; Redford, K.H.; Robinson, J.G.; Fimbel, R.; Blate, G.M. Biodiversity Conservation in the Context of Tropical Forest Management; World Bank: Washington, DC, USA, 2000. [Google Scholar]
- Stewart, O.C. Fire as the First Great Force Employed by Man. In Man’s Role in Changing the Face of the Earth; Thomas, W.L., Jr., Ed.; University of Chicago Press: Chicago, IL, USA, 1956; Volume 2, pp. 115–133. [Google Scholar]
- Whittaker, R.H. A Consideration of Climax Theory: The Climax as a Population and Pattern. Ecol. Monogr. 1953, 23, 41–78. [Google Scholar] [CrossRef]
- Stern, K.; Roche, L. Genetics of Forest Ecosystems; Chapman and Hall: London, UK, 1974; ISBN 978-0-387-06095-8. [Google Scholar]
- Anand, M. Towards a Unifying Theory of Vegetation Dynamics; Faculty of Graduate Studies, University of Western Ontario: London, ON, Canada, 1997. [Google Scholar]
- Lamb, D. Restoring Tropical Moist Broad-Leaf Forests. In Forest Restoration in Landscapes: Beyond Planting Trees; Mansourian, S., Vallauri, D., Dudley, N., Eds.; Springer: New York, NY, USA, 2005; pp. 291–297. ISBN 978-0-387-29112-3. [Google Scholar]
- Whitmore, T.C. Changes Over Twenty-One Years in the Kolombangara Rain Forests. J. Ecol. 1989, 77, 469–483. [Google Scholar] [CrossRef]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R.; Wan, P. Plant Functional Traits Suggest a Change in Novel Ecological Strategies for Dominant Species in the Stages of Forest Succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef]
- Yu, R.; Huang, J.; Xu, Y.; Ding, Y.; Zang, R. Plant Functional Niches in Forests Across Four Climatic Zones: Exploring the Periodic Table of Niches Based on Plant Functional Traits. Front. Plant Sci. 2020, 11, 841. [Google Scholar] [CrossRef]
- Evstigneev, O.I. Features of Undergrowth Development in Eastern European Forests. Russ. J. Ecosyst. Ecol. 2019, 4, 1–23. [Google Scholar] [CrossRef]
- Král, K.; Shue, J.; Vrška, T.; Gonzalez-Akre, E.B.; Parker, G.G.; McShea, W.J.; McMahon, S.M. Fine-Scale Patch Mosaic of Developmental Stages in Northeast American Secondary Temperate Forests: The European Perspective. Eur. J. For. Res. 2016, 135, 981–996. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; ISBN 978-0-470-85040-4. [Google Scholar]
- Holmberg, T.J. 4.2.3: Life Histories and Natural Selection. Available online: https://bio.libretexts.org/Sandboxes/tholmberg_at_nwcc.edu/BIOL_1213/04%3A_Unit_4%3A__Ecology/4.02%3A_Population_Ecology/4.2.03%3A_Life_Histories_and_Natural_Selection (accessed on 2 May 2023).
- Chuine, I. Why Does Phenology Drive Species Distribution? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies That Shape Ecosystems; John Wiley & Sons: Chichester, UK, 2012; ISBN 978-0-470-67481-9. [Google Scholar]
- Metz, J.A.J.; Nisbet, R.M.; Geritz, S.A.H. How Should We Define ‘Fitness’ for General Ecological Scenarios? Trends Ecol. Evol. 1992, 7, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Tinner, W.; Ammann, B. Long-Term Responses of Mountain Ecosystems to Environmental Changes: Resilience, Adjustment, and Vulnerability. In Global Change and Mountain Regions: An Overview of Current Knowledge; Huber, U.M., Bugmann, H.K.M., Reasoner, M.A., Eds.; Advances in Global Change Research; Springer: Dordrecht, The Netherlands, 2005; pp. 133–143. ISBN 978-1-4020-3508-1. [Google Scholar]
- Yamamoto, S. Gap Regeneration of Major Tree Species in Different Forest Types of Japan. Vegetatio 1996, 127, 203–213. [Google Scholar] [CrossRef]
- Evstigneev, O.I. Ontogenetic Scales of Relation of Trees to Light (on the Example of Eastern European Forests). Russ. J. Ecosyst. Ecol. 2018, 3, 1–18. [Google Scholar] [CrossRef]
- McVean, D.N. Alnus glutinosa (L.) Gaertn. J. Ecol. 1953, 41, 447–466. [Google Scholar] [CrossRef]
- Dickson, J.H. Pleistocene History of Betula with Special Reference to the British Isles. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1984, 85, 1–11. [Google Scholar] [CrossRef]
- Steijlen, I.; Zackrisson, O. Long-Term Regeneration Dynamics and Successional Trends in a Northern Swedish Coniferous Forest Stand. Can. J. Bot. 1987, 65, 839–848. [Google Scholar] [CrossRef]
- Rogers, P.C.; Pinno, B.D.; Šebesta, J.; Albrectsen, B.R.; Li, G.; Ivanova, N.; Kusbach, A.; Kuuluvainen, T.; Landhäusser, S.M.; Liu, H.; et al. A Global View of Aspen: Conservation Science for Widespread Keystone Systems. Glob. Ecol. Conserv. 2020, 21, e00828. [Google Scholar] [CrossRef]
- Savill, P.S. The Silviculture of Trees Used in British Forestry, 3rd ed.; CABI: Wallingford, UK, 2019; ISBN 978-1-78639-392-0. [Google Scholar]
- Kuehne, C.; Nosko, P.; Horwath, T.; Bauhus, J. A Comparative Study of Physiological and Morphological Seedling Traits Associated with Shade Tolerance in Introduced Red Oak (Quercus rubra) and Native Hardwood Tree Species in Southwestern Germany. Tree Physiol. 2014, 34, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Kuliesis, A.; Kasperavicius, A.; Kulbokas, G.; Kvalkauskiene, M. Lithuanian National Forest Inventory, 2003–2007. Forest Resources and Their Dynamic; Lututė: Kaunas, Lithuania, 2009. [Google Scholar]
- Boratyńska, K.; Dolatowski, J. Systematics and geographical distribution. In Limes–Tilia cordata Mill., Tilia platyphyllos Scop.; Białobok, S., Ed.; Monografie popularnonaukowe; PAN, Instytut Dendrologii: Poznań, Poland, 1991; Volume 5. [Google Scholar]
- Walter, H. Vegetation of the Earth and Ecological Systems of the Geo-Biosphere, 2nd ed.; Heidelberg Science Library; Springer: New York, NY, USA, 2012; ISBN 978-1-4684-0468-5. [Google Scholar]
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech Regeneration Research: From Ecological to Silvicultural Aspects. For. Ecol. Manag. 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Hemery, G.E.; Clark, J.R.; Aldinger, E.; Claessens, H.; Malvolti, M.E.; O’connor, E.; Raftoyannis, Y.; Savill, P.S.; Brus, R. Growing Scattered Broadleaved Tree Species in Europe in a Changing Climate: A Review of Risks and Opportunities. For. Int. J. For. Res. 2010, 83, 65–81. [Google Scholar] [CrossRef]
- Franklin, J. Regeneration and Growth of Pioneer and Shade-tolerant Rain Forest Trees in Tonga. N. Z. J. Bot. 2003, 41, 669–684. [Google Scholar] [CrossRef]
- Hickler, T.; Vohland, K.; Feehan, J.; Miller, P.A.; Smith, B.; Costa, L.; Giesecke, T.; Fronzek, S.; Carter, T.R.; Cramer, W.; et al. Projecting the Future Distribution of European Potential Natural Vegetation Zones with a Generalized, Tree Species-Based Dynamic Vegetation Model. Glob. Ecol. Biogeogr. 2012, 21, 50–63. [Google Scholar] [CrossRef]
- WorldData.Info Climate and Temperature Development in Lithuania 2023. Available online: https://www.worlddata.info/europe/lithuania/climate.php (accessed on 17 August 2023).
- Gedminas, A.; Lynikienè, J.; Marciulynas, A.; Povilaitiene, A. Effect of Physokermes Piceae Schrank. on Shoot and Needle Growth in Norway Spruce Stands in Lithuania. Balt. For. 2015, 21, 162–169. [Google Scholar]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Myking, T.; Bøhler, F.; Austrheim, G.; Solberg, E.J. Life History Strategies of Aspen (Populus tremula L.) and Browsing Effects: A Literature Review. For. Int. J. For. Res. 2011, 84, 61–71. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; Wallenius, T.; Kauhanen, H. Tree Mortality Episodes in the Intact Picea Abies-Dominated Taiga in the Arkhangelsk Region of Northern European Russia. J. Veg. Sci. 2011, 22, 322–333. [Google Scholar] [CrossRef]
- Stobbe, A.; Gumnior, M. Palaeoecology as a Tool for the Future Management of Forest Ecosystems in Hesse (Central Germany): Beech (Fagus sylvatica L.) versus Lime (Tilia cordata Mill.). Forests 2021, 12, 924. [Google Scholar] [CrossRef]
- Plesa, I.M.; González-Orenga, S.; Al Hassan, M.; Sestras, A.F.; Vicente, O.; Prohens, J.; Sestras, R.E.; Boscaiu, M. Effects of Drought and Salinity on European Larch (Larix decidua Mill.) Seedlings. Forests 2018, 9, 320. [Google Scholar] [CrossRef]
- Gabrilavičius, R.; Petrokas, R.; Danusevičius, J. Rare Tree Species in Lithuanian Forests; Baltic Printing House: Klaipėda, Lithuania, 2013; ISBN 978-609-460-072-2. [Google Scholar]
- Makrickas, E.; Manton, M.; Angelstam, P.; Grygoruk, M. Trading Wood for Water and Carbon in Peatland Forests? Rewetting Is Worth More than Wood Production. J. Environ. Manag. 2023, 341, 117952. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.; Janson, C.H. The Relationship between Seed Size and Establishment Conditions in Tropical Woody Plants. Ecology 1985, 66, 773–780. [Google Scholar] [CrossRef]
- Walters, M.B.; Reich, P.B. Are Shade Tolerance, Survival, and Growth Linked? Low Light and Nitrogen Effects on Hardwood Seedlings. Ecology 1996, 77, 841–853. [Google Scholar] [CrossRef]
- Hillebrand, H.; Kunze, C. Meta-Analysis on Pulse Disturbances Reveals Differences in Functional and Compositional Recovery across Ecosystems. Ecol. Lett. 2020, 23, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Akça, E.; de Alba, S.; Álvarez, A.G.; Bialousz, S.; Berger, B.; Bielek, P.; Blum, W.; Breuning-Madsen, H.; Buivydaite, V.V.; Cangir, C.; et al. Soil Atlas of Europe; European Soil Bureau Network: Luxembourg, 2005; ISBN 978-92-894-8120-5. [Google Scholar]
- Świtoniak, M.; Mroczek, P.; Bednarek, R. Luvisols or Cambisols? Micromorphological Study of Soil Truncation in Young Morainic Landscapes—Case Study: Brodnica and Chełmno Lake Districts (North Poland). CATENA 2016, 137, 583–595. [Google Scholar] [CrossRef]
- Hugh, M.F. The Periglacial Environment, 3rd ed.; John Wiley & Sons: Chichester, UK, 2017; ISBN 978-0-470-86588-0. [Google Scholar]
- Wilson, D.S.; Sober, E. Reviving the Superorganism. J. Theor. Biol. 1989, 136, 337–356. [Google Scholar] [CrossRef]
- Dercole, F.; Ferriere, R.; Rinaldi, S. Chaotic Red Queen Coevolution in Three-Species Food Chains. Proc. Biol. Sci. 2010, 277, 2321–2330. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species, 1859, 1st ed.; Routledge: London, UK, 2004; ISBN 978-0-203-50910-4. [Google Scholar]
- Bader, M.K.-F.; Leuzinger, S. Hydraulic Coupling of a Leafless Kauri Tree Remnant to Conspecific Hosts. iScience 2019, 19, 1238–1247. [Google Scholar] [CrossRef]
- Simard, S.W.; Perry, D.A.; Jones, M.D.; Myrold, D.D.; Durall, D.M.; Molina, R. Net Transfer of Carbon between Ectomycorrhizal Tree Species in the Field. Nature 1997, 388, 579–582. [Google Scholar] [CrossRef]
- Klein, T.; Siegwolf, R.T.W.; Körner, C. Belowground Carbon Trade among Tall Trees in a Temperate Forest. Science 2016, 352, 342–344. [Google Scholar] [CrossRef]
- Rosenau, J. Natural Selection/Survival of the Fittest. Available online: https://ncse.ngo/natural-selection-survival-fittest (accessed on 6 April 2023).
- Hall, B.K.; Strickberger, M.W.; Hallgrímsson, B. Strickberger’s Evolution, 4th ed.; Jones & Bartlett Learning: Sudbury, MA, USA, 2008; ISBN 978-0-7637-0066-9. [Google Scholar]
- Heskel, M.; Pengra, J.; Kruper, A.; Anderson, M.D.; Dosch, J.J.; Goldstein, L.; Hahn, S.; Hoffman, S. Age and Phenology Control Photosynthesis and Leaf Traits in the Understory Woody Species, Rhamnus Cathartica and Prunus Serotina. AoB Plants 2022, 14, plac044. [Google Scholar] [CrossRef]
- Poulter, B.; MacBean, N.; Hartley, A.; Khlystova, I.; Arino, O.; Betts, R.; Bontemps, S.; Boettcher, M.; Brockmann, C.; Defourny, P. Plant Functional Type Classification for Earth System Models: Results from the European Space Agency’s Land Cover Climate Change Initiative. Geosci. Model Dev. 2015, 8, 2315–2328. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Forest Site Types | Dominant Ground Vegetation Types | Forest Stand Types |
|---|---|---|
| Temporarily over moist eutrophic | Oxalido-nemorosa | Piceetum, Quercetum, Fraxinetum, Populetum, Betuletum pendulae, Alnetum |
| Normally moist mesotrophic | Oxalidosa | Piceetum, Pinetum, Populetum, Betuletum pendulae, Quercetum |
| Temporarily over moist mesotrophic | Myrtillo-oxalidosa | Piceetum, Betuletum pendulae, Populetum, Pinetum |
| Temporarily over moist oligotrophic | Myrtillosa | Pinetum, Piceetum, Betuletum pendulae, Populetum |
| Normally moist oligotrophic | Vaccinio-myrtillosa | Pinetum, Betuletum pendulae, Populetum, Piceetum |
| Normally moist (very) oligotrophic | Vacciniosa | Pinetum, Betuletum pendulae |
| Normally moist very oligotrophic | Cladoniosa | Pinetum |
| Over moist oligotrophic | Myrtillo-sphagnosa | Pinetum, Betuletum pubescentis, Piceetum |
| Peatland oligotrophic | Carico-sphagnosa | Pinetum, Betuletum pubescentis |
| Peatland very oligotrophic | Ledo-sphagnosa | Pinetum |
| Normally moist eutrophic | Hepatico-oxalidosa | Quercetum, Piceetum, Carpinetum, Fagetum, Populetum, Betuletum pendulae |
| Normally moist very eutrophic | Aegopodiosa | Quercetum, Fraxinetum, Tilietum, Ulmetum, Populetum, Betuletum |
| Temporarily over moist very eutrophic | Carico-mixtoherbosa | Fraxinetum, Quercetum, Populetum, Betuletum, Alnetum |
| Over moist very eutrophic | Urticosa | Alnetum glutinosae, Fraxinetum, Betuletum |
| Over moist eutrophic | Filipendulo-mixtoherbosa | Alnetum glutinosae, Fraxinetum, Betuletum |
| Peatland eutrophic | Carico-iridosa | Alnetum glutinosae, Betuletum pubescentis |
| Peatland mesotrophic | Caricosa | Betuletum pubescentis, Alnetum glutinosae |
| Over moist mesotrophic | Calamagrostidosa | Betuletum pubescentis, Alnetum glutinosae |
| End Communities | Forest Disturbance Regimes | Plant Functional Groups |
|---|---|---|
| Biotic climax | Multi-cohort succession | Ruderals |
| Edaphic climax | Successional development | Stress tolerators |
| Climatic climax | Gap dynamics | Competitors |
| Development | Establishment | |
|---|---|---|
| Forest | Gaps | |
| Forest | Stress-resistant competitors: Tilia cordata, Fagus sylvatica *. | Competitive stress-sensitive ruderals: Acer platanoides, Carpinus betulus, Picea abies, Ulmus glabra, Ulmus laevis. |
| Gaps | Ruderal stress-sensitive competitors: Fraxinus excelsior, Quercus robur. | Stress-resistant ruderals: Alnus glutinosa, Alnus incana, Betula pendula, Betula pubescens, Pinus sylvestris, Populus tremula. |
| Major Soil Groups ** | Dominant Ground Vegetation Types *** | The Four Modes of Tree Establishment and Phenological Development in the Forest * | Forest Disturbance Regimes | Potential End Community | |||
|---|---|---|---|---|---|---|---|
| Stress-Resistant Ruderals | Competitive Stress-Sensitive Ruderals | Ruderal Stress-Sensitive Competitors | Stress-Resistant Competitors | ||||
| Hemi-boreal spruce forests with broadleaved trees (D19, including U10) | |||||||
| LV, CM, FL | oxn | Be Bu Pt Ai Ag | Pa Ug Ap | Fe Qr | Tc | Successional development | Climatic climax |
| AR, LV, AB, PL, CM, FL | ox, mox | Ps Be Pt | Pa | Qr | - | Successional development | Climatic climax |
| Boreal and hemi-boreal pine forests, partly with birch and spruce (D48 and D49) | |||||||
| AR, PZ | vm, m | Ps Be Pt | Pa | - | - | Multi-cohort succession | Climatic climax |
| Hemi-boreal pine forests, partly with birch (D55) | |||||||
| AR, PZ, RG | v | Ps Be | - | - | - | Multi-cohort succession | Edaphic climax |
| AR, RG | cl | Ps | - | - | - | Multi-cohort succession | Edaphic climax |
| Pine bog forests (S9) | |||||||
| GL | msp | Ps Bu | Pa | - | - | Multi-cohort succession | Edaphic climax |
| HSf-s | csp | Ps Bu | - | - | - | Multi-cohort succession | Fire climax |
| HSf | lsp | Ps | - | - | - | Multi-cohort succession | Fire climax |
| Oak-hornbeam forests (F40, including U10) | |||||||
| AR, LV, AB, PL, CM, FL | hox | Be Pt Ai | Pa Cb Ug Ul Ap | Qr | Tc Fs | Gap dynamics | Climatic climax |
| Lime-oak forests (F70, including U10) | |||||||
| LV, CM | aeg, cmh | Pt Be Bu Ag Ai | Ug Ul Ap | Qr Fe | Tc | Gap dynamics | Climatic climax |
| Swamp and fen forests (T1) | |||||||
| GL | fil, ur | Ag Bu Be Ai | Pa | Fe | - | Gap dynamics | Edaphic climax |
| HSs-ph-ef | cir | Ag Bu | Pa | - | - | Gap dynamics | Biotic climax |
| HSs-ph-mf | c | Bu Ag | Pa | - | - | Gap dynamics | Biotic climax |
| GL | cal | Bu Ag Be | Pa | - | - | Gap dynamics | Edaphic climax |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrokas, R.; Manton, M. Adaptive Relationships in Hemi-Boreal Forests: Tree Species Responses to Competition, Stress, and Disturbance. Plants 2023, 12, 3256. https://doi.org/10.3390/plants12183256
Petrokas R, Manton M. Adaptive Relationships in Hemi-Boreal Forests: Tree Species Responses to Competition, Stress, and Disturbance. Plants. 2023; 12(18):3256. https://doi.org/10.3390/plants12183256
Chicago/Turabian StylePetrokas, Raimundas, and Michael Manton. 2023. "Adaptive Relationships in Hemi-Boreal Forests: Tree Species Responses to Competition, Stress, and Disturbance" Plants 12, no. 18: 3256. https://doi.org/10.3390/plants12183256