

Transcriptomic and Physiological Analysis Reveals Genes Associated with Drought Stress Responses in Populus alba × Populus glandulosa

Abstract
:1. Introduction
2. Results
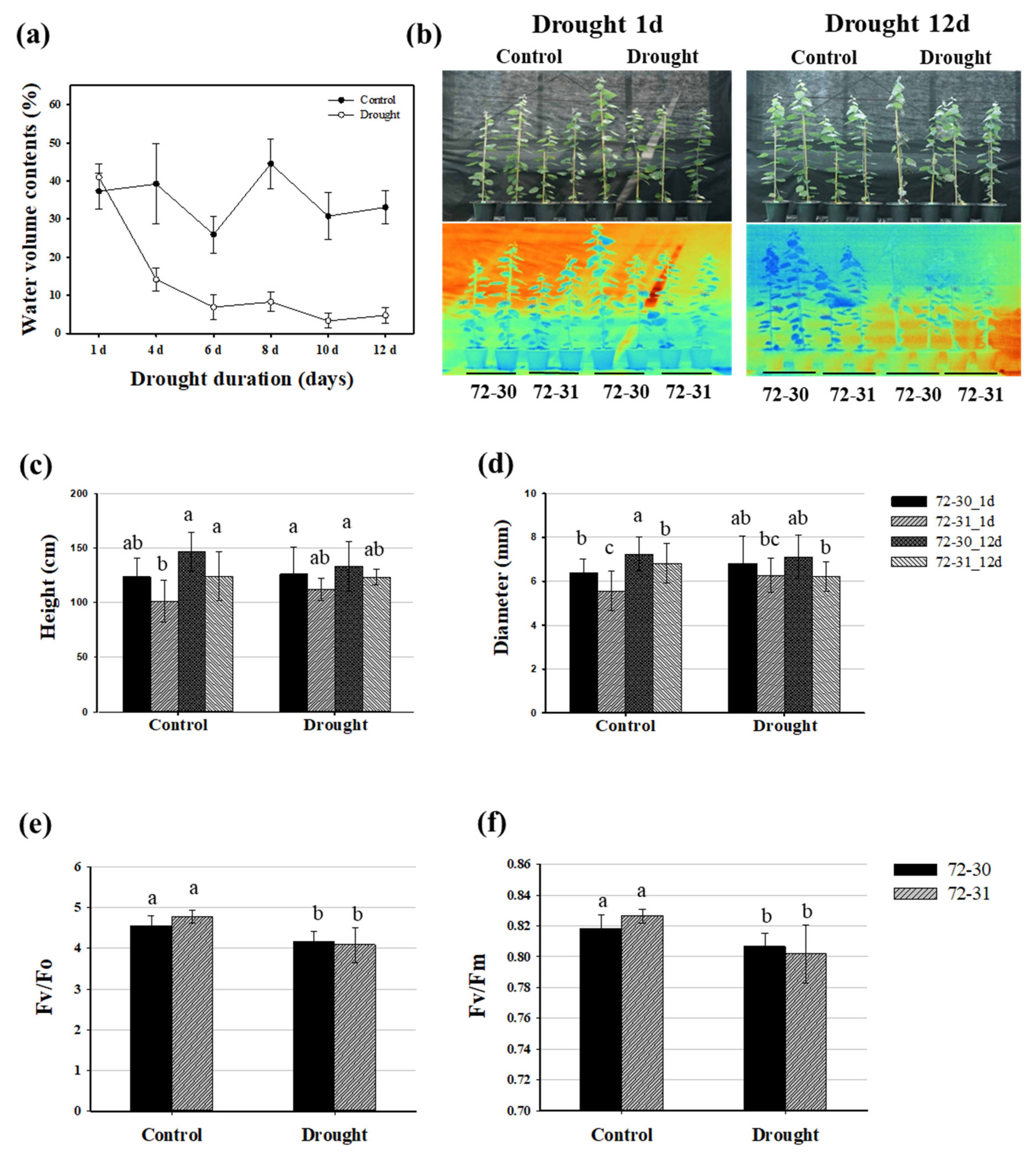
2.1. Growth and Physiological Response to Drought Stress in Poplar
2.2. Effect of Drought Stress on Soluble Sugars, Lipid Peroxidation, Proline, H2O2, and Antioxidant Activities
2.3. RNA Sequencing, Assembly, and Annotation
2.4. Analysis of Differentially Expressed Genes in the Response to Drought Stress
2.5. Functional Enrichment of DEGs in the Response to Drought Stress
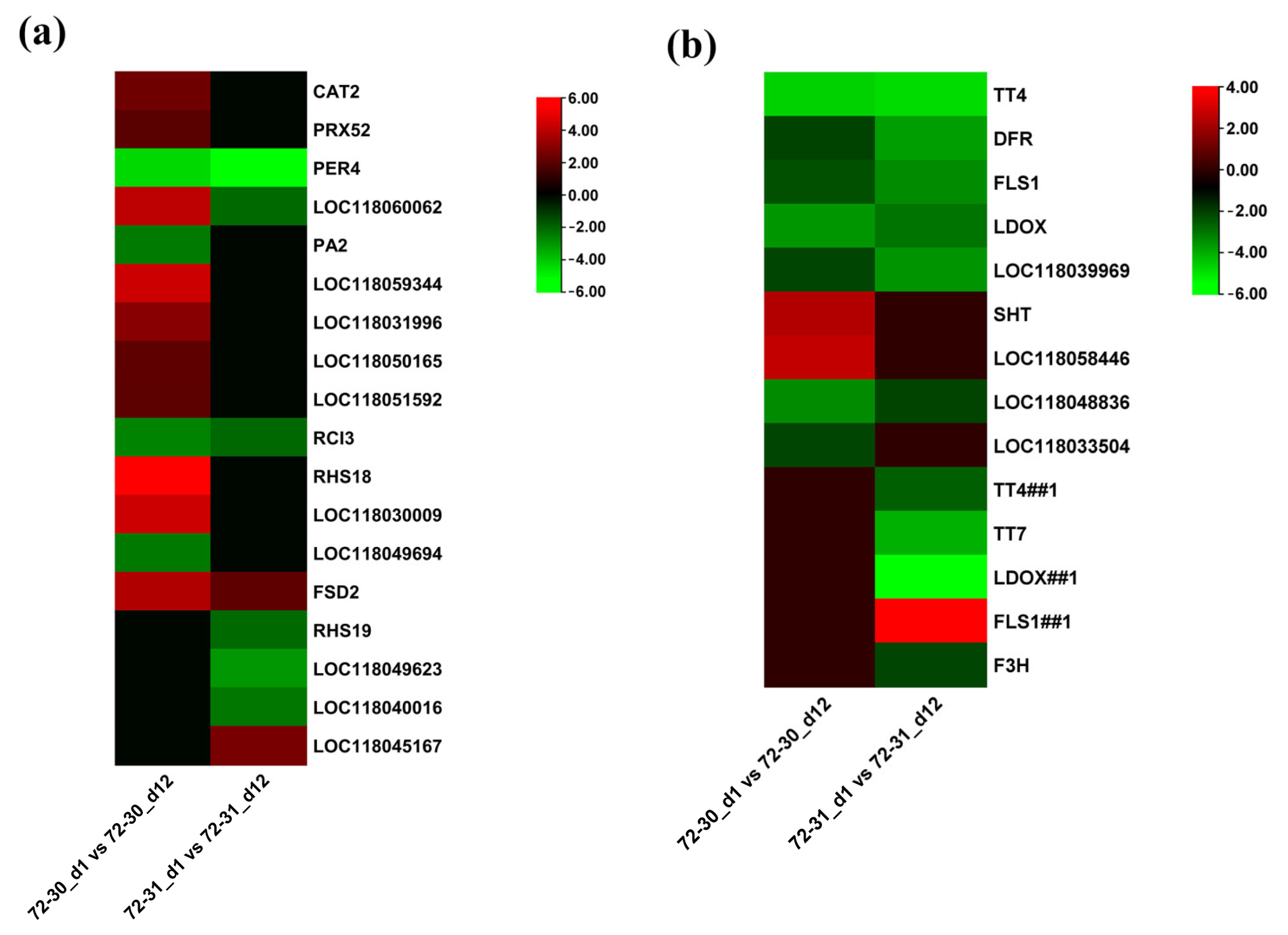
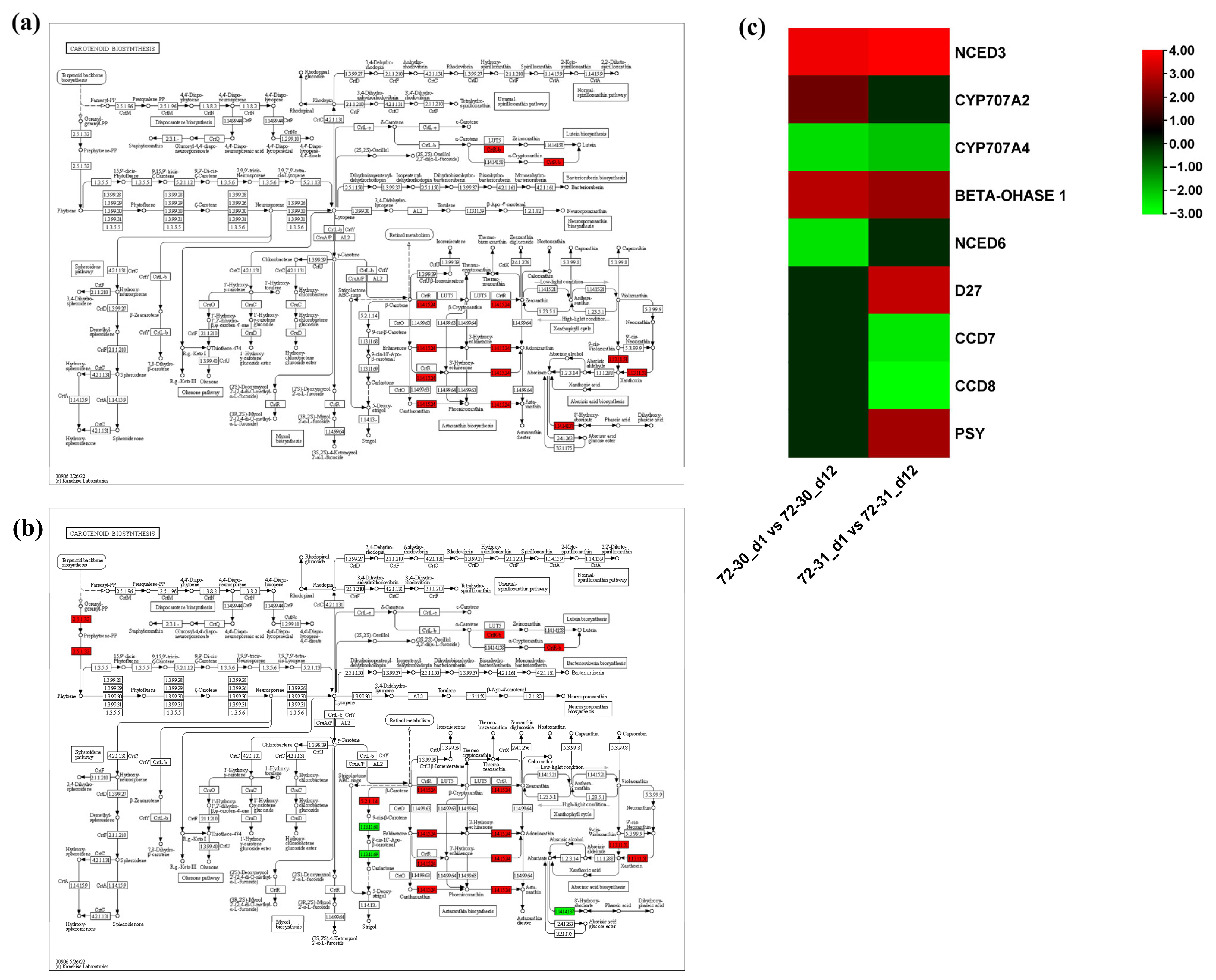
2.6. DEGs Involved in Proline, Antioxidants, and ABA
2.7. Validation of RNA-Seq Results by qPCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Drought Stress Treatment
4.2. Measurement of Chlorophyll, Soluble Sugars, Starch, Proline, MDA, H2O2, Soluble Proteins, ABA and Antioxidant Enzymes
4.3. RNA Extraction, cDNA Library Construction, Sequencing
4.4. Assembly Normalization and Quality Assessment
4.5. Differentially Expressed Genes (DEGs) Analysis
4.6. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis
4.7. Quantitative PCR (qPCR) Validation of RNA-Seq Data
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [PubMed]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought tolerance strategies in plants: A mechanistic approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [PubMed]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar]
- Todaka, D.; Zhao, Y.; Yoshida, T.; Kudo, M.; Kidokoro, S.; Mizoi, J.; Kodaira, K.S.; Takebayashi, Y.; Kojima, M.; Sakakibara, H. Temporal and spatial changes in gene expression, metabolite accumulation and phytohormone content in rice seedlings grown under drought stress conditions. Plant J. 2017, 90, 61–78. [Google Scholar]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar]
- Praba, M.L.; Cairns, J.; Babu, R.; Lafitte, H. Identification of physiological traits underlying cultivar differences in drought tolerance in rice and wheat. J. Agron. Crop Sci. 2009, 195, 30–46. [Google Scholar]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Liu, H.; Su, C.; Qian, Y.; Jiao, F. MaCDSP32 from mulberry enhances resilience post-drought by regulating antioxidant activity and the osmotic content in transgenic tobacco. Front. Plant Sci. 2020, 11, 419. [Google Scholar]
- Zhao, J.; Wang, X.; Pan, X.; Jiang, Q.; Xi, Z. Exogenous putrescine alleviates drought stress by altering reactive oxygen species scavenging and biosynthesis of polyamines in the seedlings of Cabernet Sauvignon. Front. Plant Sci. 2021, 12, 767992. [Google Scholar] [PubMed]
- Challabathula, D.; Analin, B.; Mohanan, A.; Bakka, K. Differential modulation of photosynthesis, ROS and antioxidant enzyme activities in stress-sensitive and-tolerant rice cultivars during salinity and drought upon restriction of COX and AOX pathways of mitochondrial oxidative electron transport. J. Plant Physiol. 2022, 268, 153583. [Google Scholar] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [PubMed]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [PubMed]
- Wang, P.; Liu, H.; Hua, H.; Wang, L.; Song, C.-P. A vacuole localized β-glucosidase contributes to drought tolerance in Arabidopsis. Chin. Sci. Bull. 2011, 56, 3538–3546. [Google Scholar]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar]
- Zhang, C.; Zhang, D.-W.; Sun, Y.-N.; Arfan, M.; Li, D.-X.; Yan, J.-J.; You, M.-H.; Bai, S.-Q.; Lin, H.-H. Photo-protective mechanisms in reed canary grass to alleviate photo-inhibition of PSII on the Qinghai-Tibet Plateau. J. Plant Physiol. 2017, 215, 11–19. [Google Scholar]
- Liu, Y.; Subhash, C.; Yan, J.; Song, C.; Zhao, J.; Li, J. Maize leaf temperature responses to drought: Thermal imaging and quantitative trait loci (QTL) mapping. Environ. Exp. Bot. 2011, 71, 158–165. [Google Scholar]
- Brunner, A.M.; Busov, V.B.; Strauss, S.H. Poplar genome sequence: Functional genomics in an ecologically dominant plant species. Trends Plant Sci. 2004, 9, 49–56. [Google Scholar]
- Caruso, A.; Chefdor, F.; Carpin, S.; Depierreux, C.; Delmotte, F.M.; Kahlem, G.; Morabito, D. Physiological characterization and identification of genes differentially expressed in response to drought induced by PEG 6000 in Populus canadensis leaves. J. Plant Physiol. 2008, 165, 932–941. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Z.; Bo, W.; Ren, Y.; Zhang, Z.; Zhang, D. Transcriptional profiling by cDNA-AFLP analysis showed differential transcript abundance in response to water stress in Populus hopeiensis. BMC Genom. 2012, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.-B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Jia, J.; Zhang, C.; Li, H.; Liu, T.; Jiang, X.; Polle, A.; Peng, C.; Luo, Z.B. Anatomical, physiological and transcriptional responses of two contrasting poplar genotypes to drought and re-watering. Physiol. Plant. 2014, 151, 480–494. [Google Scholar] [CrossRef]
- Cao, X.; Jia, J.; Li, H.; Li, M.; Luo, J.; Liang, Z.; Liu, T.; Liu, W.; Peng, C.; Luo, Z. Photosynthesis, water use efficiency and stable carbon isotope composition are associated with anatomical properties of leaf and xylem in six poplar species. Plant Biol. 2012, 14, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, B.T.; Marguerat, S.; Watt, S.; Schubert, F.; Wood, V.; Goodhead, I.; Penkett, C.J.; Rogers, J.; Bähler, J. Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nature 2008, 453, 1239–1243. [Google Scholar] [CrossRef]
- Han, Y.; Gao, S.; Muegge, K.; Zhang, W.; Zhou, B. Advanced applications of RNA sequencing and challenges. Bioinform. Biol. Insights 2015, 9, BBI-S28991. [Google Scholar] [CrossRef]
- Shanker, A.K.; Maheswari, M.; Yadav, S.; Desai, S.; Bhanu, D.; Attal, N.B.; Venkateswarlu, B. Drought stress responses in crops. Funct. Integr. Genom. 2014, 14, 11–22. [Google Scholar] [CrossRef]
- Matsui, A.; Ishida, J.; Morosawa, T.; Mochizuki, Y.; Kaminuma, E.; Endo, T.A.; Okamoto, M.; Nambara, E.; Nakajima, M.; Kawashima, M. Arabidopsis transcriptome analysis under drought, cold, high-salinity and ABA treatment conditions using a tiling array. Plant Cell Physiol. 2008, 49, 1135–1149. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; AbadÞa, J. Photoinhibition and photoprotection under nutrient deficiencies, drought and salinity. In Photoprotection, Photoinhibition, Gene Regulation, and Environment; Springer: Dordrecht, The Netherlands, 2006; pp. 65–85. [Google Scholar]
- Wahid, A.; Close, T. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Afzal, S.; Chaudhary, N.; Singh, N.K. Role of soluble sugars in metabolism and sensing under abiotic stress. In Plant Growth Regulators: Signalling under Stress Conditions; Springer Nature: Berlin, Germany, 2021; pp. 305–334. [Google Scholar]
- Bano, Q.; Ilyas, N.; Bano, A.; Zafar, N.; Akram, A.; Hassan, F. Effect of Azospirillum inoculation on maize (Zea mays L.) under drought stress. Pak. J. Bot. 2013, 45, 13–20. [Google Scholar]
- Azzeme, A.M.; Abdullah, S.N.A.; Aziz, M.A.; Wahab, P.E.M. Oil palm leaves and roots differ in physiological response, antioxidant enzyme activities and expression of stress-responsive genes upon exposure to drought stress. Acta Physiol. Plant. 2016, 38, 52. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, X.-Q.; Peng, Y.; Liang, X.-Y.; Huang, L.-K.; Ma, X.; Ma, Y.-M. Effects of drought stress on lipid peroxidation, osmotic adjustment and activities of protective enzymes in the roots and leaves of orchardgrass. Acta Pratacult. Sin. 2014, 23, 144. [Google Scholar]
- Dziedziński, M.; Kobus-Cisowska, J.; Stachowiak, B. Pinus species as prospective reserves of bioactive compounds with potential use in functional food—Current state of knowledge. Plants 2021, 10, 1306. [Google Scholar] [CrossRef]
- Peskin, A.V.; Winterbourn, C.C. Assay of superoxide dismutase activity in a plate assay using WST-1. Free Radic. Biol. Med. 2017, 103, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef]
- Hilal, M.; Parrado, M.F.; Rosa, M.; Gallardo, M.; Orce, L.; Massa, E.M.; González, J.A.; Prado, F.E. Epidermal Lignin Deposition in Quinoa Cotyledons in Response to UV-B Radiation. Photochem. Photobiol. 2004, 79, 205–210. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline accumulation influenced by osmotic stress in arbuscular mycorrhizal symbiotic plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar] [CrossRef]
- Shao, H.-B.; Chu, L.-Y.; Jaleel, C.A.; Manivannan, P.; Panneerselvam, R.; Shao, M.-A. Understanding water deficit stress-induced changes in the basic metabolism of higher plants–biotechnologically and sustainably improving agriculture and the ecoenvironment in arid regions of the globe. Crit. Rev. Biotechnol. 2009, 29, 131–151. [Google Scholar] [CrossRef]
- Tuteja, R. Malaria− an overview. FEBS J. 2007, 274, 4670–4679. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Schumaker, K.; Zhu, J.K. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J. Exp. Bot. 2004, 55, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Deltoro, V.I.; Calatayud, A.; Gimeno, C.; Abadía, A.; Barreno, E. Changes in chlorophyll a fluorescence, photosynthetic CO2 assimilation and xanthophyll cycle interconversions during dehydration in desiccation-tolerant and intolerant liverworts. Planta 1998, 207, 224–228. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; López-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biol. 2009, 11, 339–350. [Google Scholar] [CrossRef]
- Berger, K.; Machwitz, M.; Kycko, M.; Kefauver, S.C.; Van Wittenberghe, S.; Gerhards, M.; Verrelst, J.; Atzberger, C.; van der Tol, C.; Damm, A. Multi-sensor spectral synergies for crop stress detection and monitoring in the optical domain: A review. Remote Sens. Environ. 2022, 280, 113198. [Google Scholar] [CrossRef]
- Yang, R.; Yang, T.; Zhang, H.; Qi, Y.; Xing, Y.; Zhang, N.; Li, R.; Weeda, S.; Ren, S.; Ouyang, B. Hormone profiling and transcription analysis reveal a major role of ABA in tomato salt tolerance. Plant Physiol. Biochem. 2014, 77, 23–34. [Google Scholar] [CrossRef]
- Lu, C.-M.; Zhang, J.-H. Heat-induced multiple effects on PSII in wheat plants. J. Plant Physiol. 2000, 156, 259–265. [Google Scholar] [CrossRef]
- Ruban, A.V.; Saccon, F. Chlorophyll a de-excitation pathways in the LHCII antenna. J. Chem. Phys. 2022, 156, 070902. [Google Scholar] [CrossRef]
- Berwal, M.; Ram, C. Superoxide dismutase: A stable biochemical marker for abiotic stress tolerance in higher plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019; Available online: http://dx.doi.org/10.5772/intechopen.82079 (accessed on 30 June 2023).
- Hikosaka, K.; Terashima, I. A model of the acclimation of photosynthesis in the leaves of C3 plants to sun and shade with respect to nitrogen use. Plant Cell Environ. 1995, 18, 605–618. [Google Scholar] [CrossRef]
- Zhang, W.; Chu, Y.; Ding, C.; Zhang, B.; Huang, Q.; Hu, Z.; Huang, R.; Tian, Y.; Su, X. Transcriptome sequencing of transgenic poplar (Populus× euramericana’Guariento’) expressing multiple resistance genes. BMC Genet. 2014, 15, S7. [Google Scholar] [CrossRef] [PubMed]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity. PLoS Pathog. 2012, 8, e1002684. [Google Scholar] [CrossRef] [PubMed]
- Hessini, K.; Cruz, C.; Gandour, M.; Soltani, A.; Abdelly, C. Do reactive oxygen species (ROS) induced by NaCl contribute to ammonium accumulation in Spartina alterniflora? J. Plant Nutr. Soil Sci. 2009, 172, 851–860. [Google Scholar] [CrossRef]
- Kim, T.-L.; Lee, K.; Hwang, H.-S.; Oh, C.; Lee, I.H.; Lim, H. Comparison of Physiological and Biochemical Responses of Two Poplar Species under Drought Stress. Plant Breed. Biotechnol. 2022, 10, 145–162. [Google Scholar] [CrossRef]
- Kim, T.-L.; Lim, H.; Chung, H.; Veerappan, K.; Oh, C. Elevated CO2 Alters the Physiological and Transcriptome Responses of Pinus densiflora to Long-Term CO2 Exposure. Plants 2022, 11, 3530. [Google Scholar] [CrossRef]
- Shibaeva, T.; Mamaev, A.; Sherudilo, E. Evaluation of a SPAD-502 plus chlorophyll meter to estimate chlorophyll content in leaves with interveinal chlorosis. Russ. J. Plant Physiol. 2020, 67, 690–696. [Google Scholar] [CrossRef]
- Stitt, M.; Lilley, R.M.; Gerhardt, R.; Heldt, H.W. [32]Metabolite levels in specific cells and subcellular compartments of plant leaves. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1989; Volume 174, pp. 518–552. [Google Scholar]
- Bates, L.; Waldren, R.a.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: II. Role of electron transfer. Arch. Biochem. Biophys. 1968, 125, 850–857. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pettengill, E.A.; Parmentier-Line, C.; Coleman, G.D. Evaluation of qPCR reference genes in two genotypes of Populus for use in photoperiod and low-temperature studies. BMC Res. Notes 2012, 5, 366. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Treatment | mg/g FW | Chl a/b | Chl/Car | |||
|---|---|---|---|---|---|---|---|
| Chl a | Chl b | Total Chl | Carotenoids | ||||
| 72-30 | Control | 1.76 ± 0.09 ab | 0.73 ± 0.04 a | 2.49 ± 0.31 a | 0.53 ± 0.02 a | 2.42 ± 0.07 b | 4.70 ± 0.11 n.s |
| Drought | 1.92 ± 0.07 a | 0.72 ± 0.04 ab | 2.68 ± 0.20 a | 0.55 ± 0.02 a | 2.68 ± 0.16 a | 4.84 ± 0.01 n.s | |
| 72-31 | Control | 1.51 ± 0.10 b | 0.64 ± 0.05 b | 2.14 ± 0.15 b | 0.45 ± 0.02 b | 2.38 ± 0.06 b | 4.81 ± 0.11 n.s |
| Drought | 1.91 ± 0.15 a | 0.76 ± 0.05 a | 2.68 ± 0.20 a | 0.57 ± 0.05 a | 2.51 ± 0.07 ab | 4.68 ± 0.13 n.s | |
| Clone | Treatment | mg/g FW | |||
|---|---|---|---|---|---|
| Glucose | Fructose | Sucrose | Starch | ||
| 72-30 | Control | 1.18 ± 0.10 d | 1.00 ± 0.11 c | 54.70 ± 1.20 b | 0.28 ± 0.11 ab |
| Drought | 2.46 ± 0.25 b | 2.69 ± 0.04 a | 82.38 ± 2.45 a | 0.28 ± 0.08 ab | |
| 72-31 | Control | 2.10 ± 0.13 c | 2.03 ± 0.25 b | 51.51 ± 3.97 b | 0.37 ± 0.12 a |
| Drought | 2.92 ± 0.22 a | 3.01 ± 0.42 a | 49.83 ± 5.30 b | 0.19 ± 0.05 b | |
| Sample | Raw Reads | Clean Reads | Mapped (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|
| 72-30_d1_1 | 21,322,525 | 19,423,209 | 91.94% | 95 | 47 |
| 72-30_d1_2 | 25,534,053 | 23,613,885 | 92.22% | 95 | 48 |
| 72-30_d1_3 | 27,750,144 | 24,773,305 | 84.14% | 95 | 49 |
| 72-30_d12_1 | 29,270,661 | 26,337,948 | 90.26% | 95 | 47 |
| 72-30_d12_2 | 29,362,659 | 26,669,548 | 89.28% | 95 | 48 |
| 72-30_d12_3 | 32,321,191 | 29,446,757 | 88.63% | 95 | 46 |
| 72-31_d1_1 | 28,614,343 | 26,106,663 | 86.77% | 95 | 47 |
| 72-31_d1_2 | 32,949,652 | 29,442,606 | 89.40% | 95 | 46 |
| 72-31_d1_3 | 32,479,330 | 30,465,003 | 88.08% | 95 | 48 |
| 72-31_d12_1 | 30,944,699 | 29,702,711 | 85.40% | 95 | 48 |
| 72-31_d12_2 | 29,104,362 | 27,656,786 | 81.95% | 95 | 49 |
| 72-31_d12_3 | 27,334,799 | 26,081,336 | 80.33% | 95 | 50 |
| Sum | 346,988,418 | 319,719,757 | |||
| Mean | 28,915,701.5 | 26,643,313.08 | 87.37% | 95 | 47.7 |
| Gene ID | Gene Description | Log2 FC | p-Value | TAIR ID | Gene Symbol | Regulation |
|---|---|---|---|---|---|---|
| LogFC > 0 | ||||||
| LOC118043229 | galactinol synthase 2-like | 12.27314 | 1.13 × 10−13 | AT2G47180 | GolS1 | Up |
| LOC118043085 | bidirectional sugar transporter SWEET15-like | 11.40774 | 2.21 × 10−21 | AT5G13170 | SAG29 | Up |
| LOC118053155 | zeatin O-xylosyltransferase-like | 10.90792 | 2.54 × 10−18 | AT2G15490 | UGT73B4 | Up |
| LOC118057602 | uncharacterized protein | 10.08275 | 5.60 × 10−11 | AT5G50360 | Up | |
| LOC118033421 | embryonic protein DC-8 | 9.94831 | 1.14 × 10−14 | AT2G36640 | ECP63 | Up |
| LOC118062093 | dehydration-responsive element-binding protein 2D-like | 9.40484 | 5.60 × 10−13 | AT1G75490 | Up | |
| LOC118052105 | umecyanin-like | 9.22683 | 2.11 × 10−11 | AT2G25060 | ENODL14 | Up |
| LOC118061594 | non-specific lipid-transfer protein-like protein | 9.08816 | 6.39 × 10−6 | AT3G22600 | Up | |
| LOC118034863 | late embryogenesis abundant protein D-7 | 9.00364 | 2.88 × 10−13 | AT3G15670 | Up | |
| LOC118055598 | glucose and ribitol dehydrogenase-like | 8.91445 | 9.97 × 10−28 | AT1G54870 | Up | |
| LogFC < 0 | ||||||
| LOC118040537 | auxin-induced protein 15A-like | −5.69993 | 0.0013 | AT4G34770 | Down | |
| LOC118059218 | uncharacterized protein | −5.49169 | 0.0040 | AT4G04745 | Down | |
| LOC118047630 | acetyl-CoA-benzylalcohol acetyltransferase-like | −5.44358 | 0.0007 | AT3G26040 | Down | |
| LOC118039754 | protein RADIALIS-like 2 | −5.42122 | 0.0039 | AT1G75250 | RL6 | Down |
| LOC118044774 | uncharacterized protein | −5.39230 | 0.0065 | AT4G12690 | Down | |
| LOC118043148 | protein HEADING DATE 3A-like | −5.24861 | 0.0306 | AT1G65480 | FT | Down |
| LOC118063338 | syntaxin-112-like | −5.18643 | 0.0395 | AT2G18260 | SYP112 | Down |
| LOC118053660 | uncharacterized protein | −5.17855 | 0.0099 | AT3G01410 | Down | |
| LOC118042489 | cyclin-U4-1-like | −5.16138 | 0.0385 | AT2G44740 | CYCP4;1 | Down |
| LOC118058270 | RNA exonuclease 4-like | −5.13317 | 8.59 × 10−5 | AT3G27970 | Down |
| Gene ID | Gene Description | Log 2FC | p-Value | TAIR ID | Gene Symbol | Regulation |
|---|---|---|---|---|---|---|
| LogFC > 0 | ||||||
| LOC118057602 | uncharacterized protein | 8.97992 | 7.9 × 10−19 | AT5G50360 | Up | |
| LOC118056377 | probable protein phosphatase 2C 75 isoform X1 | 7.09109 | 4.6 × 10−6 | AT5G51760 | AHG1 | Up |
| LOC118028052 | WUSCHEL-related homeobox 2-like | 6.87533 | 2.2 × 10−5 | AT5G59340 | WOX2 | Up |
| LOC118043229 | galactinol synthase 2-like | 6.48803 | 3.5 × 10−61 | AT2G47180 | GolS1 | Up |
| LOC118047047 | uncharacterized protein | 6.37664 | 4.0 × 10−9 | AT1G07985 | Up | |
| LOC118044732 | uncharacterized protein | 6.14501 | 5.2 × 10−31 | AT5G50360 | Up | |
| LOC118032224 | uncharacterized protein | 6.08772 | 6.9 × 10−4 | AT4G33467 | Up | |
| LOC118046914 | translocator protein homolog | 6.07722 | 6.3 × 10−6 | AT2G47770 | TSPO | Up |
| LOC118053986 | putative disease resistance RPP13-like protein 1 | 5.97234 | 9.5 × 10−4 | AT3G14470 | Up | |
| LOC118029876 | heavy metal-associated isoprenylated plant protein 27 | 5.91057 | 4.6 × 10−3 | AT5G66110 | HIPP27 | Up |
| LogFC < 0 | ||||||
| LOC118058995 | auxin-responsive protein SAUR24-like | −6.84085 | 2.3 × 10−6 | AT5G18020 | SAUR20 | Down |
| LOC118061976 | ornithine decarboxylase-like | −6.12173 | 1.2 × 10−4 | AT5G11880 | Down | |
| LOC118056925 | uncharacterized protein | −5.77476 | 8.6 × 10−4 | AT3G15115 | Down | |
| LOC118045856 | major allergen Pru ar 1-like | −5.69965 | 1.3 × 10−3 | AT1G24020 | MLP423 | Down |
| LOC118039754 | protein RADIALIS-like 2 | −5.61201 | 1.1 × 10−3 | AT1G75250 | RL6 | Down |
| LOC118044774 | uncharacterized protein | −5.61180 | 1.1 × 10−4 | AT4G12690 | Down | |
| LOC118035984 | axial regulator YABBY 1-like | −5.56260 | 1.4 × 10−3 | AT2G45190 | AFO | Down |
| LOC118051349 | probable serine/threonine-protein kinase PBL7 | −5.48302 | 4.1 × 10−4 | AT1G54820 | Down | |
| LOC118028545 | leucoanthocyanidin dioxygenase-like | −5.46618 | 2.9 × 10−5 | AT4G22880 | LDOX | Down |
| LOC118055368 | terpene synthase 10-like | −5.45600 | 3.3 × 10−2 | AT3G25830 | TPS-CIN | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-L.; Lim, H.; Denison, M.I.J.; Oh, C. Transcriptomic and Physiological Analysis Reveals Genes Associated with Drought Stress Responses in Populus alba × Populus glandulosa. Plants 2023, 12, 3238. https://doi.org/10.3390/plants12183238
Kim T-L, Lim H, Denison MIJ, Oh C. Transcriptomic and Physiological Analysis Reveals Genes Associated with Drought Stress Responses in Populus alba × Populus glandulosa. Plants. 2023; 12(18):3238. https://doi.org/10.3390/plants12183238
Chicago/Turabian StyleKim, Tae-Lim, Hyemin Lim, Michael Immanuel Jesse Denison, and Changyoung Oh. 2023. "Transcriptomic and Physiological Analysis Reveals Genes Associated with Drought Stress Responses in Populus alba × Populus glandulosa" Plants 12, no. 18: 3238. https://doi.org/10.3390/plants12183238