
Growth, Physiology and Nutritional Quality of C4 Halophyte Portulaca oleracea L. Grown Aeroponically in Different Percentages of Artificial Seawater under Different Light-Emitting Diode Spectral Qualities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
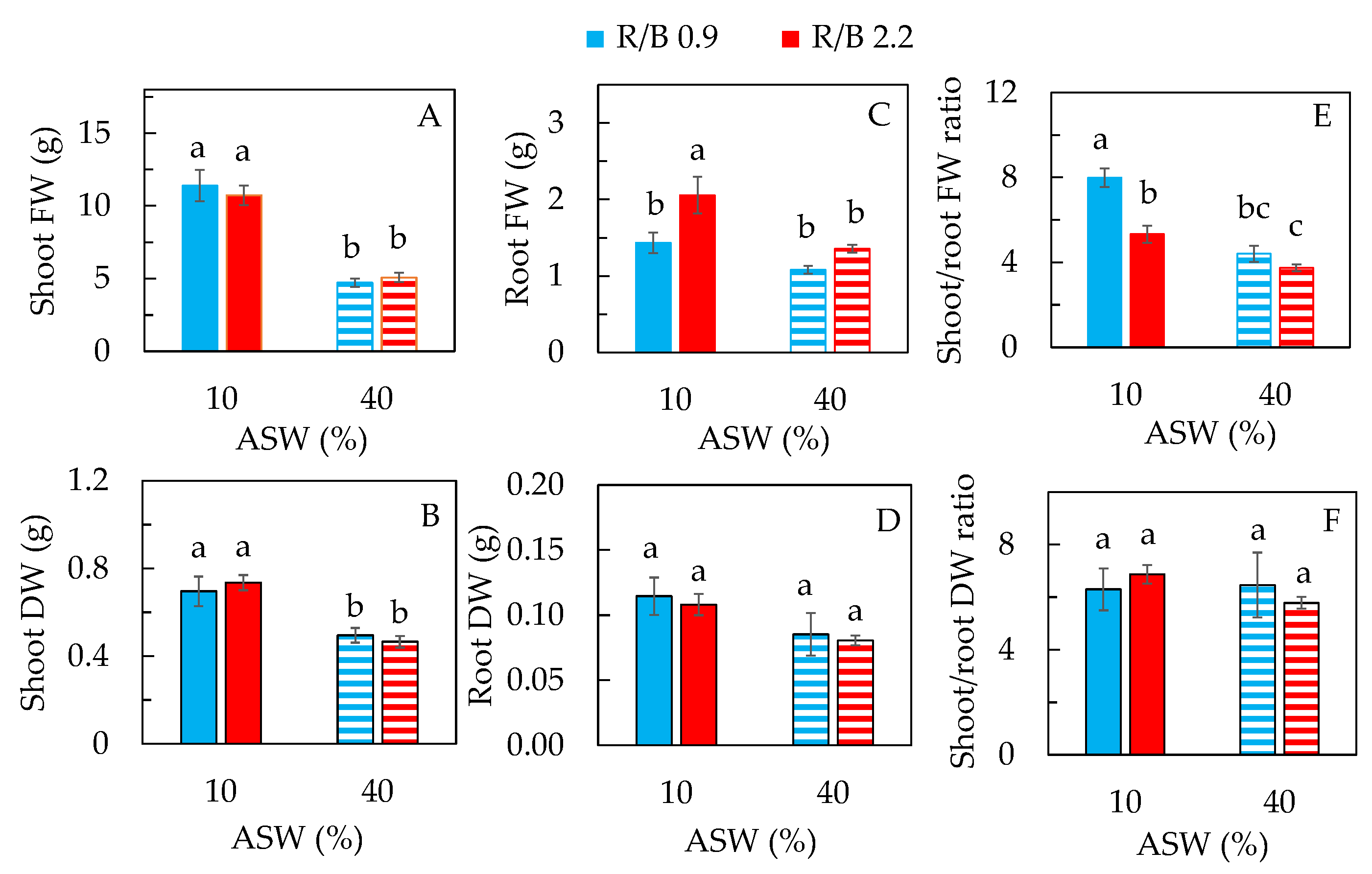
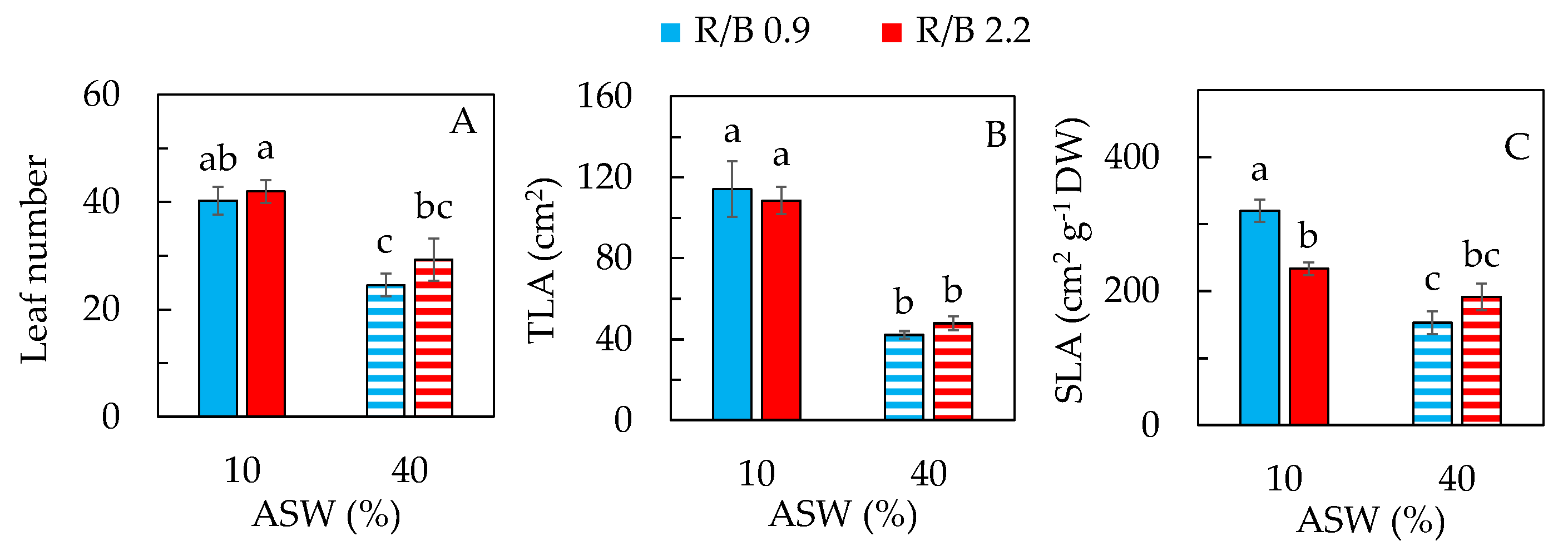
2.1. Productivity and Leaf Traits
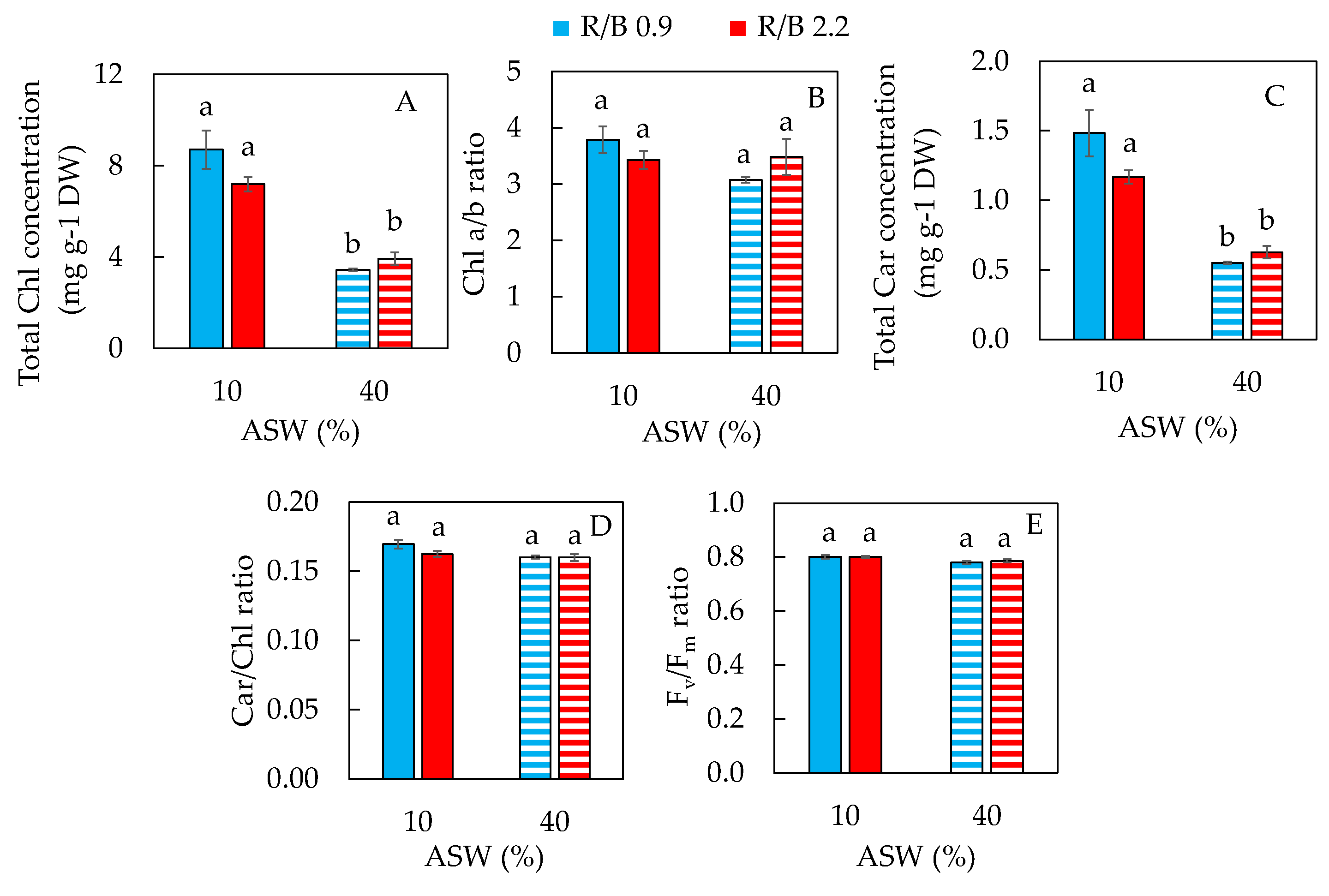
2.2. Photosynthetic Pigments and Maximum Quantum Efficiency of PSII
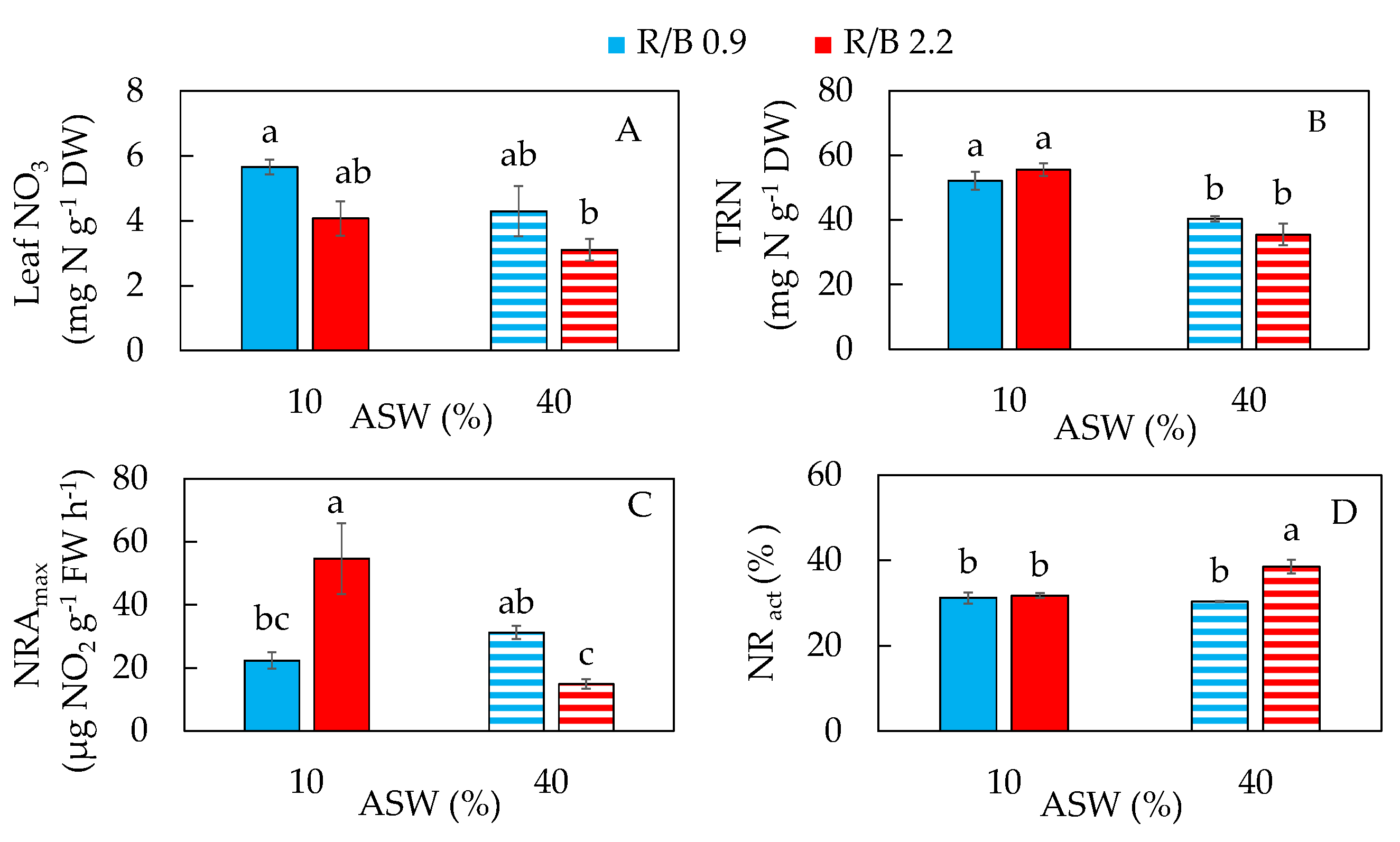
2.3. Nitrogen Metabolism
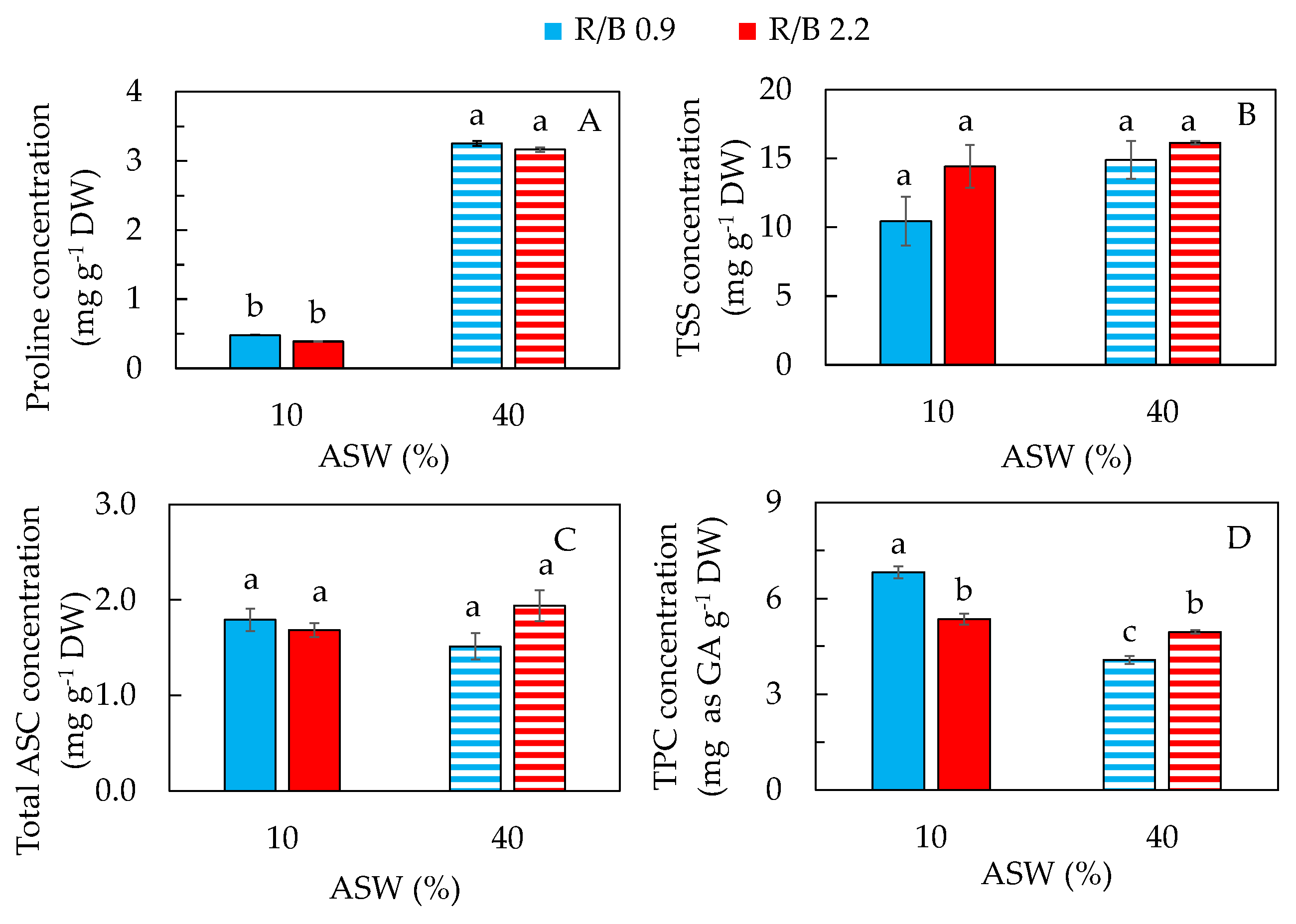
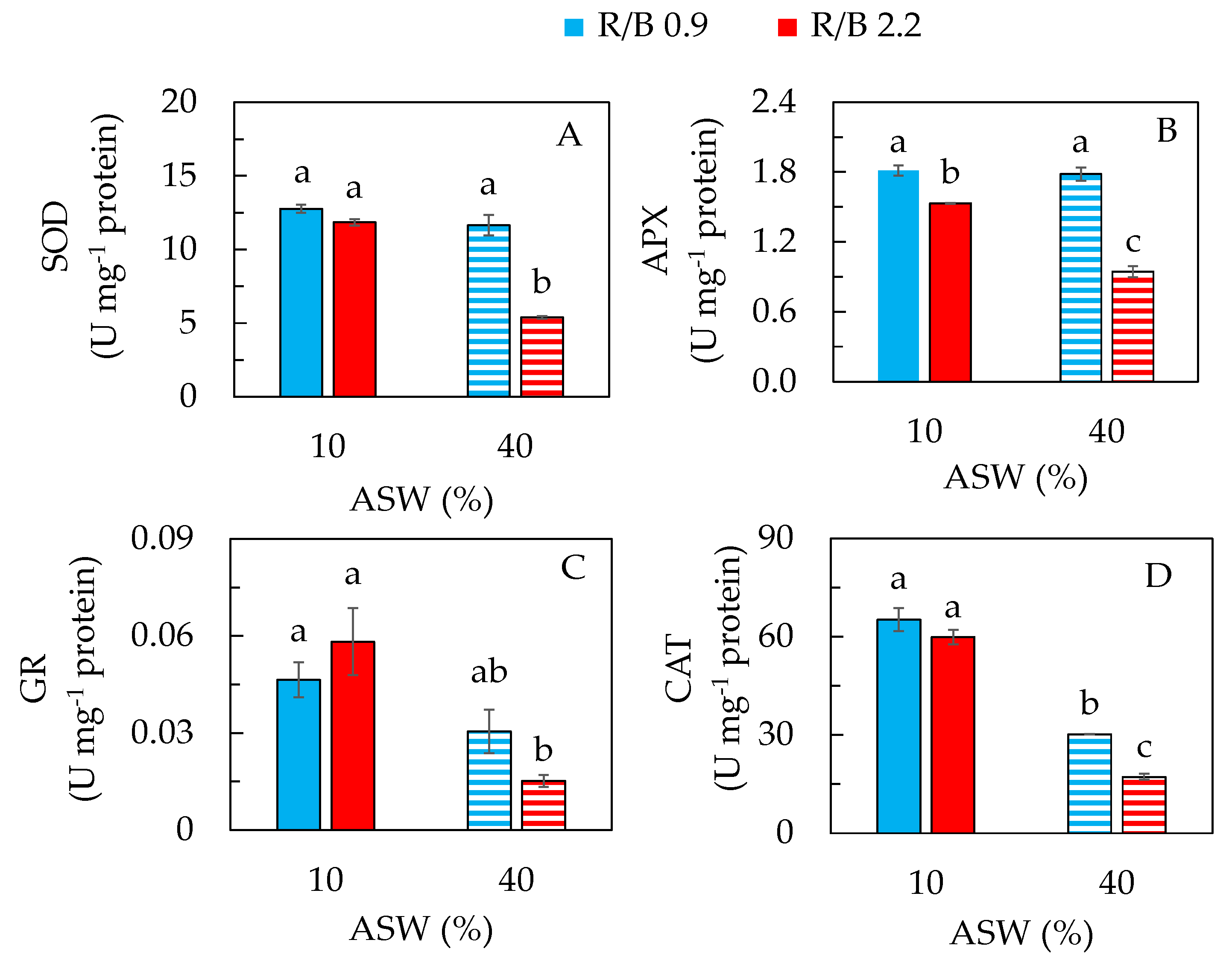
2.4. Phytochemical, Antioxidant and Antioxidant Enzyme Activities
3. Materials and Methods
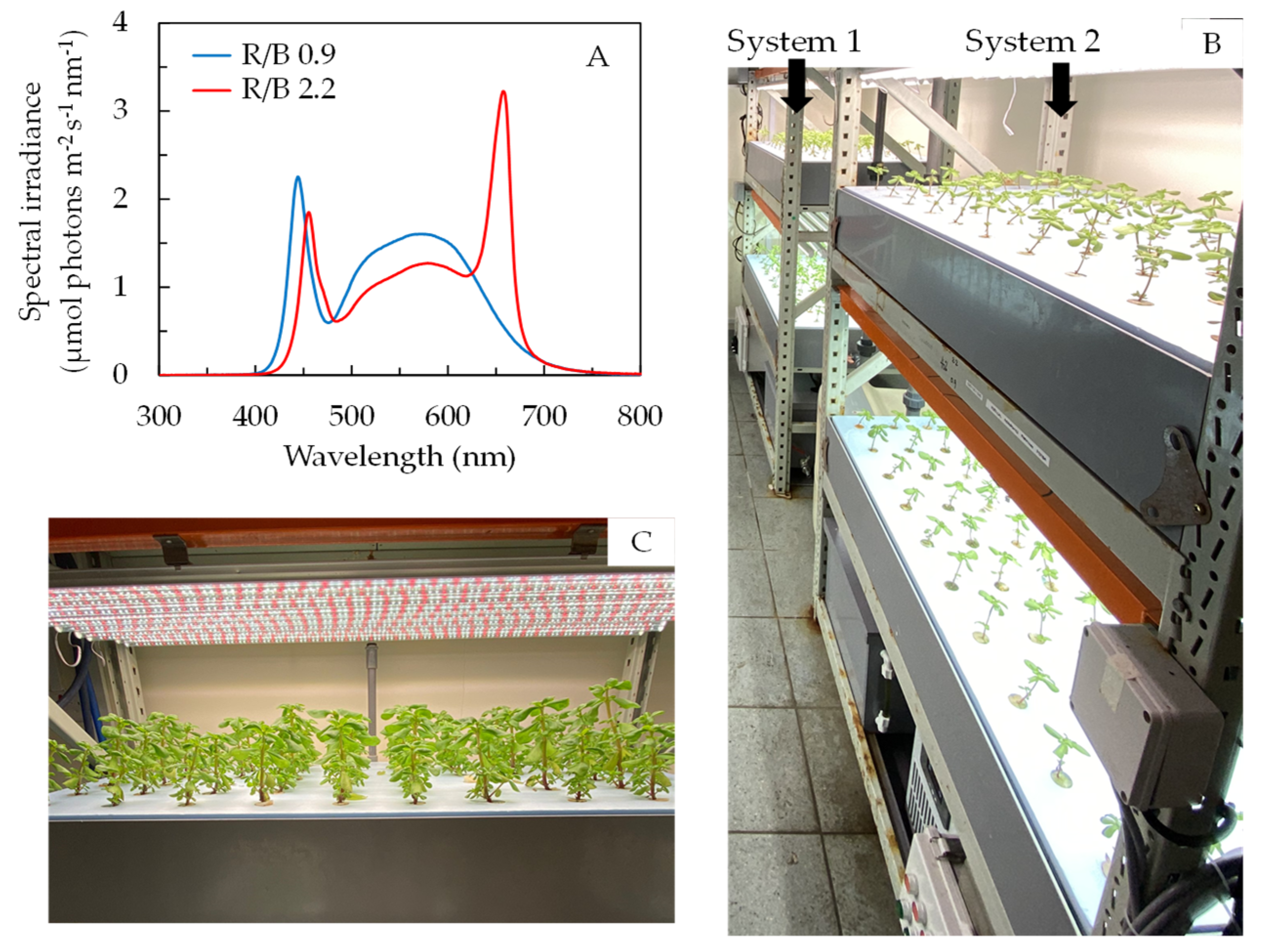
3.1. Plant Materials
3.2. Measurement of Shoot and Root Productivity and Leaf Traits
3.3. Measurement of Photosynthetic Pigments
3.4. Measurement of Chl Fluorescence Fv/Fm Ratio
3.5. Measurements of Leaf NO3− and TRN
3.6. Measurements of NRAmax and NRact
3.7. Determination of TSS Concentration
3.8. Determinations of Proline, Total ASC and TPC Concentrations
3.9. Determination of Activities of Antioxidant Enzymes
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ocampo, G.; Columbus, J.T. Molecular phylogenetics, historical biogeography, and chromosome number evolution of Portulaca (Portulacaceae). Mol. Phylogenet. Evol. 2012, 63, 97–112. [Google Scholar] [CrossRef]
- Carrascosa, A.; Pascual, J.A.; Ros, M.; Petropoulos, S.A.; Alguacil, M.d.M. Agronomical practices and management for commercial cultivation of Portulaca oleracea as a Crop: A Review. Plants 2023, 12, 1246. [Google Scholar] [CrossRef]
- Nyffeler, R.; Eggli, U. Disintegrating portulacaceae: A new familial classification of the suborder portulacineae (Caryophyllales) based on molecular and morphological data. Taxon 2010, 59, 227–240. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Dias, M.I.; Vasilakoglou, I.B.; Petrotos, K.; Barros, L.; Ferreira, I.C.F.R. Nutritional value, chemical composition and cytotoxic properties of common purslane (Portulaca oleracea L.) in relation to harvesting stage and plant part. Antioxidants 2019, 8, 293. [Google Scholar] [CrossRef]
- Kumar, A.; Sreedharan, S.; Kashyap, A.K.; Singh, P.; Ramchiary, N.A. A review on bioactive phytochemicals and ethnopharmacological potential of purslane (Portulaca oeracea L.). Heliyon 2021, 8, e08669. [Google Scholar] [CrossRef]
- Chugh, V.; Mishra, V.; Sharma, K. Purslane (Portulaca oleracea L.): An underutilized wonder plant with potential pharmacological value. Pharma Innov. 2019, 8, 236–246. [Google Scholar]
- Petropoulos, S.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Phytochemical composition and bioactive compounds of common purslane (Portulaca oleracea L.) as affected by crop management practices. Trends Food Sci. Technol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- He, J.; You, X.; Qin, L. High salinity reduces plant growth and photosynthetic performance but enhances certain nutritional quality of C4 halophyte Portulaca oleracea L. grown hydroponically under LED lighting. Front. Plant Sci. 2021, 12, 651341. [Google Scholar] [CrossRef]
- Gonnella, M.; Charfeddine, M.; Conversa, G.; Santamaria, P. Purslane: Review of its potential for health and agricultural aspect. Eur. J. Sci. Biotechol. 2010, 4, 131–136. [Google Scholar]
- D’Andrea, R.M.; Andreo, C.S.; Lara, M.V. Deciphering the mechanisms involved in Portulaca oleracea (C4) response to drought: Metabolic changes including crassulacean acid-like metabolism induction and reversal upon re-watering. Physiol. Plant. 2014, 152, 414–430. [Google Scholar] [CrossRef]
- Mulry, K.R.; Hanson, B.A.; Dudle, D.A. Alternative strategies in response to saline stress in two varieties of Portulaca oleracea (purslane). PLoS ONE 2015, 10, e0138723. [Google Scholar] [CrossRef]
- He, J.; Qin, L. Impacts of reduced nitrate supply on nitrogen metabolism, photosynthetic light-use efficiency, and nutritional values of edible Mesembryanthemum crystallinum. Front. Plant Sci. 2021, 12, 686910. [Google Scholar] [CrossRef]
- Waisel, Y. Biology of Halophytes; Academic: New York, NY, USA, 1972. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Kafi, M.; Rahimi, Z. Effect of salinity and silicon on root characteristics, growth, water status, proline content and ion accumulation of purslane (Portulaca oleracea L.). J. Soil Sci. 2011, 57, 341–347. [Google Scholar] [CrossRef]
- Anastaćio, A.; Carvalho, I.S. Accumulation of fatty acids in purslane grown in hydroponic salt stress conditions. Int. J. Food Sci. Nutr. 2013, 64, 235–242. [Google Scholar] [CrossRef]
- Yuan, F.; Guo, J.; Shabala, S.; Wang, B. Reproductive physiology of halophytes: Current standing. Front. Plant Sci. 2019, 9, 1954. [Google Scholar] [CrossRef]
- Reyna-Llorens, I.; Aubry, S. As right as rain: Deciphering drought-related metabolic flexibility in the C4–CAM Portulaca. J. Exp. Bot. 2022, 73, 4615–4619. [Google Scholar] [CrossRef]
- Yazici, I.; Türkan, I.; Sekmen, A.H.; Demiral, T. Salinity tolerance of purslane (Portulaca oleracea L.) is achieved by enhanced antioxidative system, lower level of lipid peroxidation and proline accumulation. Environ. Exp. Bot. 2007, 61, 49–57. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- He, J.; Koh, D.J.Q.; Qin, L. LED spectral quality and NaCl salinity interact to affect growth, photosynthesis and phytochemical production of Mesembryanthemum crystallinum. Funct. Plant Biol. 2022, 49, 483–495. [Google Scholar] [CrossRef]
- Sdouga, D.; Amor, F.B.; Ghribi, S.; Kabtni, S.; Tebini, M.; Branca, F.; Trifi-Farah, N.; Marghali, S. An insight from tolerance to salinity stress in halophyte Portulaca oleracea L.: Physio-morphological, biochemical and molecular responses. Ecotoxicol. Environ. Saf. 2019, 172, 45–52. [Google Scholar] [CrossRef]
- Alam, M.A.; Juraimi, A.S.; Yusop, M.R.; Hamid, A.A.; Hakim, A. Morphophysiological and mineral nutrient characterization of 45 collected purslane (Portulaca oleracea L.) accessions. Bragantia 2014, 73, 426–437. [Google Scholar] [CrossRef]
- Giménez, A.; Martínez-Ballesta, M.d.C.; Egea-Gilabert, C.; Gómez, P.A.; Artés-Hernández, F.; Pennisi, G.; Orsini, F.; Crepaldi, A.; Fernández, J.A. Combined effect of salinity and LED lights on the yield and quality of purslane (Portulaca oleracea L.) microgreens. Horticulturae 2021, 7, 180. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Ageyeva, M.; Veselov, A.; Vodeneev, V.; Brilkina, A. cell-type-specific length and cytosolic pH response of superficial cells of Arabidopsis root to chronic salinity. Plants 2022, 11, 3532. [Google Scholar] [CrossRef]
- Pungin, A.; Lartseva, L.; Loskutnikova, V.; Shakhov, V.; Popova, E.; Skrypnik, L.; Krol, O. Effect of salinity stress on phenolic compounds and antioxidant activity in halophytes Spergularia marina (L.) Griseb and Glaux maritima L. Cultured In Vitro. Plants 2023, 12, 1905. [Google Scholar] [CrossRef]
- Liu, Y.; Su, M.; Han, Z. Effects of NaCl stress on the growth, physiological characteristics and anatomical structures of Populus talassica X Populus euphratica seedlings. Plants 2022, 11, 3025. [Google Scholar] [CrossRef]
- Xie, D.; Tarin, M.W.K.; Chen, L.; Ren, K.; Yang, D.; Zhou, C.; Wan, J.; He, T.; Rong, J.; Zheng, Y. Consequences of LED lights on root morphological traits and compounds accumulation in Sarcandra glabra seedlings. Int. J. Mol. Sci. 2021, 22, 7179. [Google Scholar] [CrossRef]
- Lim, Y.J.; Eom, S.H. Effects of different light types on root formation of Ocimum basilicum L. cuttings. Sci. Hortic. 2013, 164, 552–555. [Google Scholar] [CrossRef]
- Izzo, L.G.; Hay Mele, B.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Taliesin, E.; Rodríguez, A.A.; Bustos, D.; Erdei, L.; Ortega, L.; Senn, M.E. Leaf expansion in grasses under salt stress. J. Plant Physiol. 2009, 166, 1123–1140. [Google Scholar] [CrossRef]
- Ghanem, A.E.M.F.M.; Mohamed, E.; Kasem, A.M.M.A.; El-Ghamery, A.A. Differential salt tolerance strategies in three halophytes from the same ecological habitat: Augmentation of antioxidant enzymes and compounds. Plants 2021, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Zribi, L.; Fatma, G.; Fatma, R.; Salwa, R.; Hassan, N.; Néjib, R.M. Application of chlorophyll fluorescence for the diagnosis of salt stress in tomato “Solanum lycopersicum (Variety Rio Grande)”. Sci. Hortic. 2008, 120, 367–372. [Google Scholar] [CrossRef]
- Giuffrida, F.; Scuderi, D.; Giurato, R.; Leonardi, C. Physiological response of broccoli and cauliflower as affected by NaCl salinity. Acta Hortic. 2013, 1005, 435–442. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Rabhi, M.; Castagna, A.; Remorini, D.; Scattino, C.; Smaoui, A.; Ranieri, A.; Abdelly, C. Photosynthetic responses to salinity in two obligate halophytes: Sesuvium portulacastrum and Tecticornia indica. S. Afr. J. Bot. 2012, 79, 39–47. [Google Scholar] [CrossRef]
- He, J.; Qin, L.; Chong, E.L.C.; Choong, T.-W.; Lee, S.K. Plant growth and photosynthetic characteristics of Mesembryanthemum crystallinum grown aeroponically under different blue- and red-LEDs. Front. Plant Sci. 2017, 8, 361. [Google Scholar] [CrossRef]
- Broettoa, F.; Duarteb, H.M.; Lüttge, U. Responses of chlorophyll fluorescence parameters of the facultative halophyte and C3–CAM intermediate species Mesembryanthemum crystallinum to salinity and high irradiance stress. J. Plant Physiol. 2007, 164, 904–912. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants-focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Lara, M.V.; Disante, K.; Podestá, F.E.; Andreo, C.S.; Drincovich, M.F. Induction of a crassulacean acid like metabolism in the C4 succulent plant, Portulaca oleracea L.: Physiological and morphological changes are accompanied by specific modifications in phosphoenolpyruvate carboxylase. Photosynth. Res. 2003, 77, 241–254. [Google Scholar] [CrossRef]
- Corrè, W.J.; Breimer, T. Nitrate and Nitrite in Vegetables; Pudoc: Wageningen, The Netherlands, 1979; p. 85. [Google Scholar]
- Abd-El Baki, G.K.; Siefritz, F.; Man, H.-M.; Weiner, H.; Kaldenhoff, R.; Kaiser, W.M. Nitrate reductase in Zea mays L. under salinity. Plant Cell Environ. 2000, 23, 515–521. [Google Scholar] [CrossRef]
- Liu, A.R.; Zhao, K.F. Effects of salt stress on the growth and the nitrate reductase activity in Thellungiella halophila. J. Plant Physiol. Mol. Biol. 2005, 31, 469–476. (In Chinese) [Google Scholar]
- Slama, S.; Bouchereau, A.; Flowers, T.; Abdelly, C.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Shi, Y.; Mu, C.; Wang, J. Differences in organic solute and metabolites of Leymus chinensis in response to different intensities of salt and alkali stress. Plants 2023, 12, 1916. [Google Scholar] [CrossRef]
- Guo, X.; Ahmad, N.; Zhao, S.; Zhao, C.; Zhong, W.; Wang, X.; Li, G. Effect of salt stress on growth and physiological properties of asparagus seedlings. Plants 2022, 11, 2836. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wang, B.; Wang, D.; Li, P.; Wang, L.; Yi, X.; Huang, Q.; Peng, M.; Guo, A. Comparative proteomics of Thellungiella halophila leaves from plants subjected to salinity reveals the importance of chloroplastic starch and soluble sugars in halophyte salt tolerance. Mol. Cell. Proteom. 2013, 12, 2174–2195. [Google Scholar] [CrossRef]
- Balibrea, M.E.; Rus-Alvarez, A.M.; Bolarín, M.C.; Pérez-Alfocea, F. Fast changes in soluble carbohydrates and proline contents in tomato seedlings in response to ionic and non-ionic iso-osmotic stresses. J. Plant Physiol. 1997, 151, 221–226. [Google Scholar] [CrossRef]
- Akhani, P.; Ghorbanli, M. A contribution to the halophytic vegetation and flora of Iran. In Towards the Rational Use of High Salinity Tolerant Plants; Lieth, H., Al-Masoom, A., Eds.; Kluwer Academic Publishers: Dodrecht, The Netherlands, 1993; Volume 1, pp. 35–44. [Google Scholar]
- Öztürk, M.; Altay, V.; Altundağ, E.; Gücel, S. 18-Halophytic plant diversity of unique habitats in Turkey: Salt Mine Caves of Çankırı and Iğdır. In Halophytes for Food Security in Dry Lands; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M.Z., Eds.; Elsevier Academic Publisher: Oxford, MS, USA, 2016; pp. 291–315. [Google Scholar] [CrossRef]
- Uddin, K.; Juraimi, A.S.; Ali, E.; Ismail, M.R. Evaluation of antioxidant properties and mineral composition of purslane (Portulaca oleracea L.) at different growth stages. Int. J. Mol. Sci. 2012, 13, 10257–10267. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Quah, E.P.L. Antioxidant properties of different cultivars of Portulaca oleracea. Food Chem. 2007, 103, 734–740. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of salt stress on growth, physiological parameters, and ionic concentration of water dropwort (Oenanthe javanica) cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Paccio, M.R. Tolerance of Kikuyu grass to long term salt stress in associated with induction of antioxidant defenses. Plant Growth Reg. 2003, 41, 57–62. [Google Scholar] [CrossRef]
- He, J.; Chua, E.L.; Qin, L. Drought does not induce crassulacean acid metabolism (CAM) but regulates photosynthesis and enhances nutritional quality of Mesembryanthemum crystallinum. PLoS ONE 2020, 15, e0229897. [Google Scholar] [CrossRef] [PubMed]
- He, J. Mineral nutrition of aeroponically grown subtropical and temperate crops in the tropics with manipulation of root-zone temperature at different growth irradiances. Plant Stress 2010, 4, 14–30. [Google Scholar]
- Welburn, A.R. The spectral determination of chloro11phylls a and b, as well as carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Kuo, T.-M.; Warner, R.L.; Kleinhofs, A. In vitro stability of nitrate reductase from barley leaves. Phytochemistry 1982, 21, 531–533. [Google Scholar] [CrossRef]
- Kaiser, W.M.; Huber, S.C. Correlation between apparent activation state of nitrate reductase (NR), NR hysteresis and degradation of NR protein. J. Exp. Bot. 1997, 48, 1367–1374. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water- stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Leipner, J.; Fracheboud, Y.; Stamp, P. Acclimation by suboptimal temperature diminishes photooxidative damage in maize leaves. Plant Cell Environ. 1997, 20, 366–372. [Google Scholar] [CrossRef]
- Ragee, S.; Abdel-Aal, E.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Droillard, M.J.; Paulin, A.; Massot, J.C. Free radical production, catalase and superoxide dismutase activities and membrane integrity during senescence of petals of cut carnations (Dianthus caryophyllus). Physiol. Plant. 1987, 71, 197–202. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by acorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Rangani, J.; Parida, A.K.; Panda, A.; Kumari, A. Coordinated changes in antioxidative enzymes protect the photosynthetic machinery from salinity induced oxidative damage and confer salt tolerance in an extreme halophyte Salvadora persica L. Front. Plant Sci. 2016, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Saikachout, S.; Jaffel Hamza, K.; Karray Bouraoui, N.; Leclerc, J.C.; Ouerghi, Z. Salt-inducted changes in antioxidative enzyme activities in shoot tissues of two atriplex varieties. Not. Bot. Horti Agrobot. 2013, 41, 115–121. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Gan, J.H.S.; Qin, L. Productivity, photosynthetic light-use efficiency, nitrogen metabolism and nutritional quality of C4 halophyte Portulaca oleracea L. grown indoors under different light intensities and durations. Front. Plant Sci. 2023, 14, 1106394. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Leng, S.Y.; Qin, L. Growth, Physiology and Nutritional Quality of C4 Halophyte Portulaca oleracea L. Grown Aeroponically in Different Percentages of Artificial Seawater under Different Light-Emitting Diode Spectral Qualities. Plants 2023, 12, 3214. https://doi.org/10.3390/plants12183214
He J, Leng SY, Qin L. Growth, Physiology and Nutritional Quality of C4 Halophyte Portulaca oleracea L. Grown Aeroponically in Different Percentages of Artificial Seawater under Different Light-Emitting Diode Spectral Qualities. Plants. 2023; 12(18):3214. https://doi.org/10.3390/plants12183214
Chicago/Turabian StyleHe, Jie, Su Yee Leng, and Lin Qin. 2023. "Growth, Physiology and Nutritional Quality of C4 Halophyte Portulaca oleracea L. Grown Aeroponically in Different Percentages of Artificial Seawater under Different Light-Emitting Diode Spectral Qualities" Plants 12, no. 18: 3214. https://doi.org/10.3390/plants12183214