
Differential Morpho-Physiological and Biochemical Responses of Duckweed Clones from Saudi Arabia to Salinity
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Salinity Treatments
2.3. Measurements
2.3.1. Vegetative Growth Parameters
2.3.2. Proline Content
2.3.3. Ion Content Analysis
2.3.4. Chlorophyll Pigment Contents
2.4. RNA Isolation and cDNA Synthesis
2.5. Primer’s Design
2.6. Quantitative Real-Time PCR (qPCR)
2.7. Statistical Analysis
3. Results
3.1. Effects of Salinity on the Growth and Morphological Features of Duckweeds
3.2. Effects of Salinity on Duckweed Yield Traits
3.3. Effect of Salinity on Duckweed Chlorophyll Content
3.4. Influence of Salinity on Duckweed Na+ and K+ Contents
3.5. Effect of Salinity on the Proline Content of Duckweed
3.6. Principal Component Analysis for Growth Parameters
3.7. Agglomerative Hierarchical Clustering (AHC) Analysis for Salt Tolerance Indices
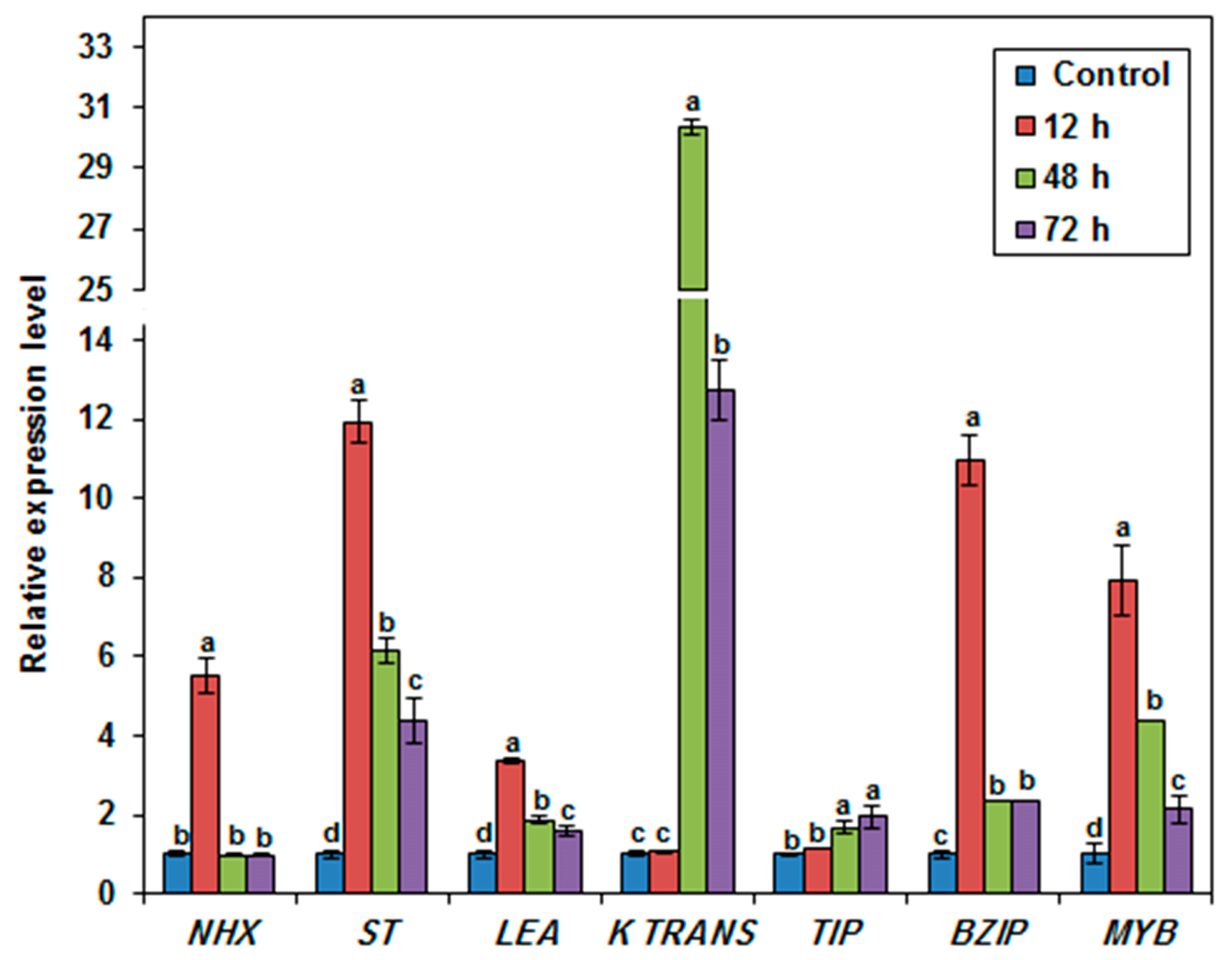
3.8. Expression Profile of Selected Stress-Related Genes in the Salt-Tolerant Duckweed Genotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.J. Relative in vitro growth rates of duckweeds (L emnaceae)–the most rapidly growing higher plants. Plant Biol. 2015, 17, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hu, S.; Li, G.; Khan, S.; Kumar, S.; Yao, L.; Duan, P.; Hou, H. Transformation Development in Duckweeds. In The Duckweed Genomes; Cao, X.H., Fourounjian, P., Wang, W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 143–155. [Google Scholar]
- Sree, K.S.; Bog, M.; Appenroth, K.J. Taxonomy of duckweeds (Lemnaceae), potential new crop plants. Emir. J. Food Agric. 2016, 28, 291–302. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, H.; Stomp, A.-M.; Cheng, J.J. The production of duckweed as a source of biofuels. Biofuels 2012, 3, 589–601. [Google Scholar]
- Cui, W.; Cheng, J.J. Growing duckweed for biofuel production: A review. Plant Biol. 2015, 17 (Suppl. S1), 16–23. [Google Scholar] [PubMed]
- An, D.; Li, C.; Zhou, Y.; Wu, Y.; Wang, W. Genomes and Transcriptomes of Duckweeds. Front. Chem. 2018, 6, 230. [Google Scholar] [CrossRef]
- Kumar, P.; Eid, E.M.; Taher, M.A.; El-Morsy, M.H.E.; Osman, H.E.M.; Al-Bakre, D.A.; Adelodun, B.; Abou Fayssal, S.; Andabaka, Z.; Goala, M.; et al. Sustainable Upcycling of Mushroom Farm Wastewater through Cultivation of Two Water Ferns (Azolla spp.) in Stagnant and Flowing Tank Reactors. Horticulturae 2022, 8, 506. [Google Scholar] [CrossRef]
- Landolt, E. The Family of Lemnaceae-a Monographic Study; Veroffentlichungen des Geobotanischen Institutes der ETH: Zurich, Switzerland, 1987; Volume 2. [Google Scholar]
- Zhang, L.M.; Jin, Y.; Yao, S.M.; Lei, N.F.; Chen, J.S.; Zhang, Q.; Yu, F.H. Growth and morphological responses of duckweed to clonal fragmentation, nutrient availability, and population density. Front. Plant Sci. 2020, 11, 618. [Google Scholar] [CrossRef]
- Dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological responses to drought, salinity, and heat stress in plants: A review. Stresses 2022, 2, 113–135. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Hillman, W.S.; Culley, D.D. The uses of duckweed: The rapid growth, nutritional value, and high biomass productivity of these floating plants suggest their use in water treatment, as feed crops, and in energy-efficient farming. Am. Sci. 1978, 66, 442–451. [Google Scholar]
- Ullah, H.; Gul, B.; Khan, H.; Zeb, U. Effect of salt stress on proximate composition of duckweed (Lemna minor L.). Heliyon 2021, 7, e07399. [Google Scholar] [PubMed]
- Kuehdorf, K.; Appenroth, K.-J. Influence of salinity and high temperature on turion formation in the duckweed Spirodela polyrhiza. Aquat. Bot. 2012, 97, 69–72. [Google Scholar] [CrossRef]
- De Morais, M.B.; Barbosa-Neto, A.G.; Willadino, L.; Ulisses, C.; Calsa Junior, T. Salt stress induces increase in starch accumulation in duckweed (Lemna aequinoctialis, Lemnaceae): Biochemical and physiological aspects. J. Plant Growth Regul. 2019, 38, 683–700. [Google Scholar]
- Fourounjian, P.; Fakhoorian, T.; Cao, X.H. Importance of duckweeds in basic research and their industrial applications. In The Duckweed Genomes; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–17. [Google Scholar]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant 2015, 37, 204. [Google Scholar]
- Chang, I.-H.; Cheng, K.-T.; Huang, P.-C.; Lin, Y.-Y.; Cheng, L.-J.; Cheng, T.-S. Oxidative stress in greater duckweed (Spirodela polyrhiza) caused by long-term NaCl exposure. Acta Physiol. Plant. 2011, 34, 1165–1176. [Google Scholar]
- Sikorski, L.; Piotrowicz-Cieslak, A.I.; Adomas, B. Phytotoxicity of sodium chloride towards common duckweed (Lemna Minor L.) and Yellow Lupin (Lupinus Luteus L.). Arch. Environ. Prot. 2013, 39, 117–128. [Google Scholar]
- Cheng, T.S.; Hung, M.J.; Cheng, Y.I.; Cheng, L.J. Calcium-induced proline accumulation contributes to amelioration of NaCl injury and expression of glutamine synthetase in greater duckweed (Spirodela polyrhiza L.). Aquat Toxicol 2013, 144–145, 265–274. [Google Scholar]
- Yang, L.; Han, H.J.; Liu, M.M.; Zuo, Z.J.; Zhou, K.Q.; Lu, J.C.; Zhu, Y.R.; Bai, Y.L.; Wang, Y. Overexpression of the Arabidopsis photorespiratory pathway gene, serine: Glyoxylate aminotransferase (AtAGT1), leads to salt stress tolerance in transgenic duckweed (Lemna minor). Plant Cell Tiss. Org. 2013, 113, 407–416. [Google Scholar]
- Hounkpe, S.; Aina, M.; Crapper, M.; Adjovi, E.; Mama, D. Influence of salinity on duckweed growth and duckweed based wastewater treatment system. J. Water Resour. Prot. 2013, 5, 993–999. [Google Scholar]
- Wang, G.H.; Chen, L.Z.; Hao, Z.J.; Li, X.Y.; Liu, Y.D. Effects of salinity stress on the photosynthesis of Wolffia arrhiza as probed by the ojip test. Fresen Environ. Bull. 2011, 20, 432–438. [Google Scholar]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.; Cho, Y.G. Plant hormones in salt stress tolerance. J. Plant Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Ding, Z.; Sun, X.; Zhang, J. Physiological and transcriptomic analysis reveals distorted ion homeostasis and responses in the freshwater plant Spirodela polyrhiza L. under Salt Stress. Genes 2019, 10, 743. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 2. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar]
- Dolferus, R. To grow or not to grow: A stressful decision for plants. Plant Sci. 2014, 229, 247–261. [Google Scholar]
- Al-Dakhil, M.; Alghamdi, S.; Migdadi, H.; Afzal, M.; Ali, A.A. Morphological Characterization and DNA Barcoding of Duckweed Species in Saudi Arabia. Plants 2021, 10, 2438. [Google Scholar] [CrossRef]
- Tarroum, M.; Ben Romdhane, W.; Ali, A.A.M.; Al-Qurainy, F.; Al-Doss, A.; Fki, L.; Hassairi, A. Harnessing the rhizosphere of the halophyte drass Aeluropus littoralis for halophilic plant-growth-promoting fungi and evaluation of their biostimulant activities. Plants 2021, 10, 784. [Google Scholar] [CrossRef]
- Ceschin, S.; Leacche, I.; Pascucci, S.; Abati, S. Morphological study of Lemna minuta Kunth, an alien species often mistaken for the native L. minor L. (Araceae). Aquat. Bot. 2016, 131, 51–56. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hoenig, M.; Baeten, H.; Vanhentenrijk, S.; Vassileva, E.; Quevauviller, P. Critical discussion on the need for an efficient mineralization procedure for the analysis of plant material by atomic spectrometric methods. Anal. Chim. Acta 1998, 358, 85–94. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Shi, H.J.; Wang, M.L.; Cui, L.; Zhao, H.; Zhao, Y. Effect of nitrogen and phosphorus deficiency on transcriptional regulation of genes encoding key enzymes of starch metabolism in duckweed (Landoltia punctata). Plant Physiol Biochem 2015, 86, 72–81. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mustafa, H.M.; Hayder, G. Recent studies on applications of aquatic weed plants in phytoremediation of wastewater: A review article. Ain Shams Eng. J. 2021, 12, 355–365. [Google Scholar] [CrossRef]
- Forni, C.; Tommasi, F. Duckweed: A tool for ecotoxicology and a candidate for phytoremediation. Curr. Biotechnol. 2016, 5, 2–10. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Okon, O.G. Effect of salinity on physiological processes in plants. In Microorganisms in Saline Environments: Strategies and Functions; Springer: Berlin/Heidelberg, Germany, 2019; pp. 237–262. [Google Scholar]
- Pazzaglia, J.; Reusch, T.B.H.; Terlizzi, A.; Marin-Guirao, L.; Procaccini, G. Phenotypic plasticity under rapid global changes: The intrinsic force for future seagrasses survival. Evol. Appl. 2021, 14, 1181–1201. [Google Scholar] [CrossRef] [PubMed]
- Oostra, V.; Saastamoinen, M.; Zwaan, B.J.; Wheat, C.W. Strong phenotypic plasticity limits potential for evolutionary responses to climate change. Nat. Commun. 2018, 9, 1005. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, D.D. Effects of salinity on growth and nickel accumulation capacity of Lemna gibba (Lemnaceae). J. Hazard. Mater. 2007, 147, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.S. NaCl-induced responses in giant duckweed (Spirodela polyrhiza). J. Aquat. Plant Manag. 2011, 59, 104–105. [Google Scholar]
- Leblebici, Z.; Aksoy, A.; Duman, F. Influence of salinity on the growth and heavy metal accumulation capacity of Spirodela polyrrhiza (Lemnaceae). Turk. J. Biol. 2011, 35, 215–220. [Google Scholar] [CrossRef]
- Sree, K.S.; Adelmann, K.; Garcia, C.; Lam, E.; Appenroth, K.J. Natural variance in salt tolerance and induction of starch accumulation in duckweeds. Planta 2015, 241, 1395–1404. [Google Scholar] [CrossRef]
- Kibria, M.G.; Hoque, M.A. A review on plant responses to soil salinity and amelioration strategies. Open J. Soil Sci. 2019, 9, 219. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Ben Romdhane, W.; El-Said, R.A.; Ghazy, A.; Attia, K.; Al-Doss, A. Agro-physiologic responses and stress-related gene expression of four doubled haploid wheat lines under salinity stress conditions. Biology 2021, 10, 56. [Google Scholar] [CrossRef]
- Petrusa, L.M.; Winicov, I. Proline status in salt-tolerant and salt-sensitive alfalfa cell lines and plants in response to NaCl. Plant Physiol. Biochem. 1997, 35, 303–310. [Google Scholar]
- Arteaga, S.; Yabor, L.; Diez, M.J.; Prohens, J.; Boscaiu, M.; Vicente, O. The Use of proline in screening for tolerance to drought and salinity in Common Bean (Phaseolus vulgaris L.) Genotypes. Agronomy 2020, 10, 817. [Google Scholar] [CrossRef]
- Gerona, M.E.B.; Deocampo, M.P.; Egdane, J.A.; Ismail, A.M.; Dionisio-Sese, M.L. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage. Rice Sci. 2019, 26, 207–219. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Proline alleviates abiotic stress induced oxidative stress in plants. J. Plant Growth Regul. 2023, 42, 4629–4651. [Google Scholar] [CrossRef]
- Bouteraa, M.T.; Mishra, A.; Romdhane, W.B.; Hsouna, A.B.; Siddique, K.H.M.; Saad, R.B. Bio-Stimulating effect of natural polysaccharides from Lobularia maritima on durum wheat seedlings: Improved plant growth, salt stress tolerance by modulating biochemical responses and ion homeostasis. Plants 2022, 11, 1991. [Google Scholar] [CrossRef] [PubMed]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, L.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Gupta, A.K.; Kaur, K.; Kaur, N. Stem reserve mobilization and sink activity in wheat under drought conditions. Am. J. Plant Sci. 2011, 02, 70–77. [Google Scholar] [CrossRef]
- Ali, A.A.M.; Ben Romdhane, W.; Tarroum, M.; Al-Dakhil, M.; Al-Doss, A.; Alsadon, A.A.; Hassairi, A. Analysis of salinity tolerance in tomato introgression lines based on Morpho-physiological and molecular Traits. Plants 2021, 10, 2594. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- Jiang, X.; Leidi, E.O.; Pardo, J.M. How do vacuolar NHX exchangers function in plant salt tolerance? Plant Signal Behav 2010, 5, 792–795. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A. Expression profiling of abiotic stress-inducible genes in response to multiple stresses in rice (Oryza sativa L.) varieties with contrasting level of stress tolerance. Biomed Res. Int. 2014, 2014, 706890. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kalita, A.; Srivastava, R.; Sahoo, L. Co-expression of Arabidopsis NHX1 and bar Improves the tolerance to salinity, oxidative stress, and herbicide in transgenic Mungbean. Front. Plant Sci. 2017, 8, 1896. [Google Scholar] [CrossRef]
- Yarra, R. The wheat NHX gene family: Potential role in improving salinity stress tolerance of plants. Plant Gene 2019, 18, 100178. [Google Scholar] [CrossRef]
- Long, L.; Zhao, J.R.; Guo, D.D.; Ma, X.N.; Xu, F.C.; Yang, W.W.; Gao, W. Identification of NHXs in Gossypium species and the positive role of GhNHX1 in salt tolerance. BMC Plant Biol. 2020, 20, 147. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, F.J.; Baghour, M.; Hao, G.; Cagnac, O.; Rodríguez-Rosales, M.P.; Venema, K. Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol. Biochem. 2012, 51, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef]
- Sun, H.; Li, L.; Lou, Y.; Zhao, H.; Yang, Y.; Wang, S.; Gao, Z. The bamboo aquaporin gene PeTIP4;1-1 confers drought and salinity tolerance in transgenic Arabidopsis. Plant Cell Rep. 2017, 36, 597–609. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence 5′–3′ | Description | Reference |
|---|---|---|---|
| NHX2-F | ATGCAGGGTTCCAGGTCAAG | Na+/H+ antiporter | JZ905382 |
| NHX2-R | AGACCGAATCTGTAGCGGC | ||
| ST-F | AGAGAC TTCGCC TCG ATTGC | Salt-tolerance protein | JZ562398 |
| ST-R | GTTCGTTTGTGGCTCGTTGG | ||
| LEA-F | GAGACACCAGTTTGGGAA CC | LEA | JZ494646 |
| LEA-R | TCTCTGACCACCCCACCTC | ||
| K+ TRANSP-F | CAAGAAGGACACGAGAGGG | Potassium transporter | JZ981156 |
| K+ TRANSP-R | GACGTGCTTGACGTACATGG | ||
| TIP4-F | GGAACTGGACGGATCACTGG | Aquaporin TIP4-1 | JZ546376 |
| TIP4-R | GGCGAACGAAGACTTCAACG | ||
| BZIP-F | CTTTTGCGGATGCAGTGGTC | BZIP2-like protein | JZ983837 |
| BZIP-R | TCATCAGGCTTCACTGTGCC | ||
| MYB306-F | ACCGAGCAGGAGGAGAAGATC | Myb-related protein | JZ982222 |
| MYB306-R | TGCTTGGCCATCTCTATGTC | ||
| ACTIN-F | GGCTACTCCTTCACCACCAC | Beta-actin | [37] |
| ACTIN-R | GCTCGTAGGTCTTCTCGACG |
| PC1 | PC2 | PC3 | PC4 | PC5 | |
|---|---|---|---|---|---|
| Eigenvalue | 7.027 | 2.386 | 1.650 | 0.730 | 0.532 |
| Variability (%) | 54.055 | 18.353 | 12.691 | 5.614 | 4.092 |
| Cumulative % | 54.055 | 72.408 | 85.099 | 90.713 | 94.805 |
| FrL | 0.743 | 0.003 | 0.129 | 0.021 | 0.001 |
| FrN | 0.605 | 0.136 | 0.011 | 0.006 | 0.182 |
| RL | 0.558 | 0.073 | 0.291 | 0.007 | 0.060 |
| RN | 0.305 | 0.442 | 0.146 | 0.000 | 0.038 |
| Chla | 0.848 | 0.016 | 0.055 | 0.023 | 0.043 |
| Chlb | 0.843 | 0.066 | 0.029 | 0.002 | 0.019 |
| Chla+b | 0.828 | 0.069 | 0.043 | 0.003 | 0.030 |
| Proline | 0.039 | 0.492 | 0.289 | 0.001 | 0.080 |
| Na+ content | 0.459 | 0.268 | 0.019 | 0.147 | 0.007 |
| K+ content | 0.453 | 0.110 | 0.383 | 0.014 | 0.022 |
| Na+/K+ ratio | 0.659 | 0.138 | 0.052 | 0.143 | 0.001 |
| FW | 0.374 | 0.303 | 0.199 | 0.016 | 0.030 |
| DW | 0.312 | 0.271 | 0.003 | 0.349 | 0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Dakhil, M.; Ben Romdhane, W.; Alghamdi, S.; Ali, A.A.M. Differential Morpho-Physiological and Biochemical Responses of Duckweed Clones from Saudi Arabia to Salinity. Plants 2023, 12, 3206. https://doi.org/10.3390/plants12183206
Al-Dakhil M, Ben Romdhane W, Alghamdi S, Ali AAM. Differential Morpho-Physiological and Biochemical Responses of Duckweed Clones from Saudi Arabia to Salinity. Plants. 2023; 12(18):3206. https://doi.org/10.3390/plants12183206
Chicago/Turabian StyleAl-Dakhil, Mohammed, Walid Ben Romdhane, Salem Alghamdi, and Ahmed Abdelrahim Mohamed Ali. 2023. "Differential Morpho-Physiological and Biochemical Responses of Duckweed Clones from Saudi Arabia to Salinity" Plants 12, no. 18: 3206. https://doi.org/10.3390/plants12183206