
Light-Emitting Diodes Modify Medicinal Quality of Mown Rabdosia rubescens, with Changes in Growth, Physiology, and Antioxidant Activity, under Drought Stress
 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Weight of Potted Seedlings Exposed to Drought
2.2. Growth, Oridonin Content, and Dry Mass
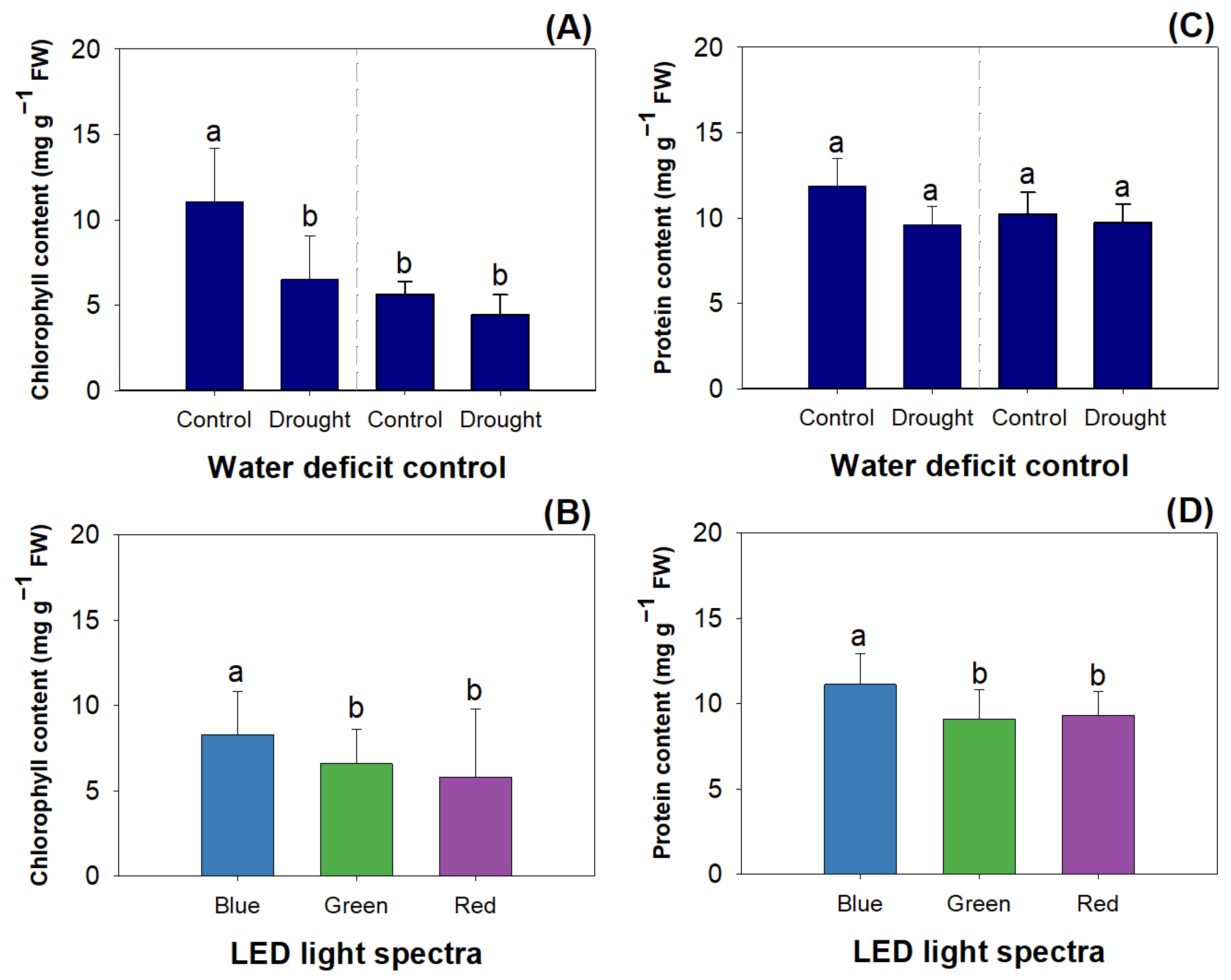
2.3. Physiology, Carbohydrate, and Nitrogen Responses
2.4. Antioxidant Activity
2.5. Regression of Medicinal Quality against Plant Parameters
3. Materials and Methods
3.1. Plant Materials
3.2. LED Light Illumination
3.3. Drought Treatment Implementation
3.4. Mowing Implementation and Sampling
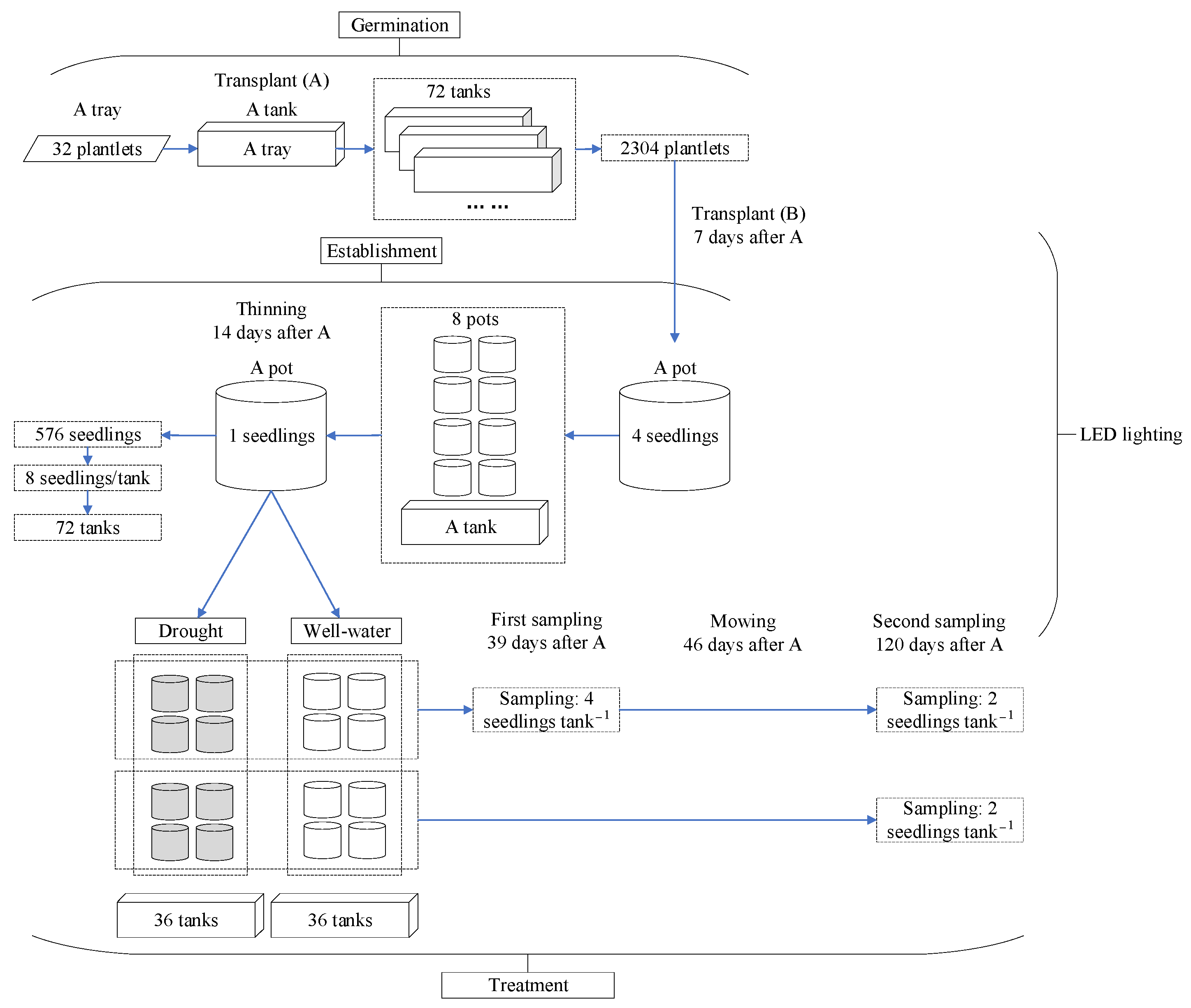
3.5. Experimental Design and Arrangement
3.6. Parameter Determination
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bisbis, M.B.; Gruda, N.; Blanke, M. Potential impacts of climate change on vegetable production and product quality—A review. J. Clean. Prod. 2018, 170, 1602–1620. [Google Scholar] [CrossRef]
- Mphande, W.; Farrell, A.D.; Kettlewell, P.S. Commercial uses of antitranspirants in crop production: A review. Outlook Agric. 2023, 52, 3–10. [Google Scholar] [CrossRef]
- Gunasekara, C.J.; Ranwala, S.M.W. Growth responses of Lantana (Lantana camara L.) varieties to varying water availability and light conditions. J. Natl. Sci. Found. Sri Lanka 2018, 46, 69–79. [Google Scholar] [CrossRef]
- He, C.X.; Zhao, Y.; Zhang, J.S.; Gao, J. Chitosan Oligosaccharide Addition to Buddhist Pine (Podocarpus macrophyllus (Thunb) Sweet) under Drought: Reponses in Ecophysiology and delta C-13 Abundance. Forests 2020, 11, 526. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Duan, Y.D.; Guo, B.T.; Zhang, L.; Li, J.X.; Li, S.; Zhao, W.B.; Yang, G.; Zhou, S.; Zhou, C.W.; Song, P.H.; et al. Interactive climate-soil forces shape the spatial distribution of foliar N:P stoichiometry in Vaccinium uliginosum planted in agroforests of Northeast China. Front. Ecol. Evol. 2022, 10, 1065680. [Google Scholar] [CrossRef]
- Byeon, S.; Kim, S.; Hong, J.; Kim, T.K.; Huh, W.; Kim, K.; Lee, M.S.; Lee, H.; Kim, S.; Park, C.; et al. Drought hardening effect on improving transplant stress tolerance in Pinus densiflora. Environ. Exp. Bot. 2023, 207, 105222. [Google Scholar] [CrossRef]
- He, J.; Chua, E.L.; Qin, L. Drought does not induce crassulacean acid metabolism (CAM) but regulates photosynthesis and enhances nutritional quality of Mesembryanthemum crystallinum. PLoS ONE 2020, 15, e0229897. [Google Scholar] [CrossRef]
- Morsi, N.A.A.; Hashem, O.S.M.; Abd El-Hady, M.A.; Abd-Elkrem, Y.M.; El-temsah, M.E.; Galal, E.G.; Gad, K.I.; Boudiar, R.; Silvar, C.; El-Hendawy, S.; et al. Assessing Drought Tolerance of Newly Developed Tissue-Cultured Canola Genotypes under Varying Irrigation Regimes. Agronomy 2023, 13, 836. [Google Scholar] [CrossRef]
- Zhang, S.H.; Xu, X.F.; Sun, Y.M.; Zhang, J.L.; Li, C.Z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef]
- Sconiers, W.B.; Rowland, D.L.; Eubanks, M.D. Pulsed drought: The effects of varying water stress on plant physiology and predicting herbivore response. Crop. Sci. 2020, 60, 2543–2561. [Google Scholar] [CrossRef]
- Abeli, T.; Ghitti, M.; Sacchi, R. Does ecological marginality reflect physiological marginality in plants? Plant Biosyst. 2020, 154, 149–157. [Google Scholar] [CrossRef]
- Cameron, R.; Harrison-Murray, R.; Fordham, M.; Wilkinson, S.; Davies, W.; Atkinson, C.; Else, M. Regulated irrigation of woody ornamentals to improve plant quality and precondition against drought stress. Ann. Appl. Biol. 2008, 153, 49–61. [Google Scholar] [CrossRef]
- Liu, M.; Pietzarka, U.; Meyer, M.; Kniesel, B.; Roloff, A.; Fini, A. Annual shoot length of temperate broadleaf species responses to drought. Urban For. Urban Green. 2022, 73, 127592. [Google Scholar] [CrossRef]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Sarrou, E.; Adamakis, I.D.; Karamanoli, K.; Martens, S.; Molassiotis, A. Metabolic mechanisms underpinning vegetative bud dormancy release and shoot development in sweet cherry. Environ. Exp. Bot. 2018, 155, 1–11. [Google Scholar] [CrossRef]
- Wei, H.X.; Zhao, H.T.; Chen, X.; He, X.Y. Secondary metabolites, carbohydrate accumulation, and nutrient uptake in Aralia elata (Miq.) Seem seedlings exposed to shoot cutting and different LED spectra. Acta Physiol. Plant 2020, 42, 162. [Google Scholar] [CrossRef]
- Zhou, C.W.; Cui, W.J.; Yuan, T.; Cheng, H.Y.; Su, Q.; Guo, P. Water content, carbohydrate accumulation, and secondary metabolites in Allium victorialis sprouts exposed to shoot cutting in varied irradiations. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12524. [Google Scholar] [CrossRef]
- Zhou, C.W.; Shang, C.F.; Chen, F.Y.; Bao, J.Z.; Yu, L.F.; Guo, P. Light-emitting diode spectra modify nutritional status, physiological response, and secondary metabolites in Ficus hirta and Alpinia oxyphylla. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12314. [Google Scholar] [CrossRef]
- He, C.X.; Zhao, Y.; Wang, Y.; Cai, J.F.; Gao, J.; Zhang, J.S. Forage quality and physiological performance of mowed alfalfa (Medicago sativa L.) subjected to combined light quality and drought. Front. Plant Sci. 2022, 13, 1047294. [Google Scholar] [CrossRef]
- Ramula, S. Annual mowing has the potential to reduce the invasion of herbaceous Lupinus polyphyllus. Biol. Invasions 2020, 22, 3163–3173. [Google Scholar] [CrossRef]
- Wei, H.X.; Chen, X.; Chen, G.S.; Zhao, H.T. Foliar nutrient and carbohydrate in Aralia elata can be modified by understory light quality in forests with different structures at Northeast China. Ann. For. Res. 2019, 62, 125–137. [Google Scholar] [CrossRef]
- Sathasivam, R.; Park, S.U.; Kim, J.K.; Park, Y.J.; Kim, M.C.; Nguyen, B.V.; Lee, S.Y. Metabolic Profiling of Primary and Secondary Metabolites in Kohlrabi (Brassica oleracea var. gongylodes) Sprouts Exposed to Different Light-Emitting Diodes. Plants 2023, 12, 1296. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.X.; Hauer, R.J.; Chen, G.S.; Chen, X.; He, X.Y. Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra. Forests 2020, 11, 44. [Google Scholar] [CrossRef]
- Ahmadi, T.; Shabani, L.; Sabzalian, M.R. LED light mediates phenolic accumulation and enhances antioxidant activity in Melissa officinalis L. under drought stress condition. Protoplasma 2020, 257, 1231–1242. [Google Scholar] [CrossRef]
- Ginzburg, D.N.; Klein, J.D. LED pre-exposure shines a new light on drought tolerance complexity in lettuce (Lactuca sativa) and rocket (Eruca sativa). Environ. Exp. Bot. 2020, 180, 104240. [Google Scholar] [CrossRef]
- Erice, G.; Louahlia, S.; Irigoyen, J.J.; Sánchez-Díaz, M.; Alami, I.T.; Avice, J.-C. Water use efficiency, transpiration and net CO2 exchange of four alfalfa genotypes submitted to progressive drought and subsequent recovery. Environ. Exp. Bot. 2011, 72, 123–130. [Google Scholar] [CrossRef]
- Cao, X.; Shen, Q.; Liu, L.; Cheng, J. Relationships of growth, stable carbon isotope composition and anatomical properties of leaf and xylem in seven mulberry cultivars: A hint towards drought tolerance. Plant Biol. 2020, 22, 287–297. [Google Scholar] [CrossRef]
- Churakova, O.V.; Lehmann, M.M.; Siegvvolf, R.T.W.; Saurer, M.; Fonti, M.V.; Schmid, L.; Timofeeva, G.; Rinne-Garmston, K.T.; Bigler, C. Compound-specific carbon isotope patterns in needles of conifer tree species from the Swiss National Park under recent climate change. Plant Physiol. Biochem. 2019, 139, 264–272. [Google Scholar] [CrossRef]
- Peng, M.F.; Liu, B.S.; Mao, M.S. Study on the Application of Chinese Patent Drug and Chinese Formula of Rabdosia Rubescens. In Proceedings of the 5th Annual International Conference on Material Science and Environmental Engineering (MSEE2017), Xiamen, China, 15–17 December 2017. [Google Scholar] [CrossRef]
- Ai, P.F.; Lu, L.P.; Song, J.J. Cryopreservation of in vitro-grown shoot-tips of Rabdosia rubescens by encapsulation-dehydration and evaluation of their genetic stability. Plant Cell Tissue Organ Cult. 2012, 108, 381–387. [Google Scholar] [CrossRef]
- Guo, J.F.; Chen, T.; Ma, Z.Y.; Qiao, C.X.; Yuan, F.L.; Guo, X.T.; Liu, J.; Shen, Y.Y.; Yu, L.; Xiang, A.Z. Oridonin inhibits 4T1 tumor growth by suppressing Treg differentiation via TGF-beta receptor. Int. Immunopharmacol. 2020, 88, 106831. [Google Scholar] [CrossRef]
- An, Y.P.; Zhu, J.; Wang, X.; Sun, X.P.; Luo, C.X.; Zhang, Y.K.; Ye, Y.W.; Li, X.W.; Abulizi, A.; Huang, Z.Z.; et al. Oridonin Delays Aging Through the AKT Signaling Pathway. Front. Pharmacol. 2022, 13, 888247. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Yuan, S.J.; Wu, H.; Chang, G.S. The anticancer effects of oridonin in vitro. Oncol. Res. 2006, 15, 501. [Google Scholar]
- Harris, E.S.J.; Cao, S.G.; Schoville, S.D.; Dong, C.M.; Wang, W.Q.; Jian, Z.Y.; Zhao, Z.Z.; Eisenberg, D.M.; Clardy, J. Selection for High Oridonin Yield in the Chinese Medicinal Plant Isodon (Lamiaceae) Using a Combined Phylogenetics and Population Genetics Approach. PLoS ONE 2012, 7, e50753. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.C.; Wu, Q.J.; Song, Z.H.; Zhang, H.; Zhang, J.F.; Zhang, L.L.; Zhang, T.Y.; Wang, C.; Wang, T. Effects of Oridonin on growth performance and oxidative stress in broilers challenged with lipopolysaccharide. Poult. Sci. 2016, 95, 2281–2289. [Google Scholar] [CrossRef]
- Turcsan, A.; Steppe, K.; Sarkozi, E.; Erdelyi, E.; Missoorten, M.; Mees, G.; Mijnsbrugge, K.V. Early Summer Drought Stress during the First Growing Year Stimulates Extra Shoot Growth in Oak Seedlings (Quercus petraea). Front. Plant Sci. 2016, 7, 193. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, R.; Liu, X.Z.; Zhou, L.J.; Dong, M.H.; Rehman, M.; Fahad, S.; Liu, L.J.; Deng, G. Effects of Light Spectra on Morphology, Gaseous Exchange, and Antioxidant Capacity of Industrial Hemp. Front. Plant Sci. 2022, 13, 937436. [Google Scholar] [CrossRef]
- Ajdanian, L.; Babaei, M.; Aroiee, H. Investigation of photosynthetic effects, carbohydrate and starch content in cress (Lepidium sativum) under the influence of blue and red spectrum. Heliyon 2020, 6, e05628. [Google Scholar] [CrossRef]
- Liu, J.Y.; Li, L.; Ji, L.; Li, Y.L.; Liu, J.J.; Li, F.Y. Divergent effects of grazing versus mowing on plant nutrients in typical steppe grasslands of Inner Mongolia. J. Plant Ecol. 2023, 16, rtac032. [Google Scholar] [CrossRef]
- Pandey, A.; Jaiswal, D.; Agrawal, S.B. Ultraviolet-B mediated biochemical and metabolic responses of a medicinal plant Adhatoda vasica Nees. at different growth stages. J. Photochem. Photobiol. B Biol. 2021, 216, 112142. [Google Scholar] [CrossRef]
- Guo, S.; Cui, X.Q.; Jiang, M.; Bai, L.; Tian, X.; Guo, T.T.; Liu, Q.C.; Zhang, L.; Ho, C.T.; Bai, N.S. Simultaneous characterization and quantification of 17 main compounds in Rabdosia rubescens by high performance liquid chromatography. J. Food Drug Anal. 2017, 25, 417–424. [Google Scholar] [CrossRef]
- Cui, H.X.; Cheng, F.R.; Yuan, K. HPLC Determination of Anti-cancer Components Isolated from Rabdosia rubescens. Pharmacogn. Mag. 2022, 18, 1196–1201. [Google Scholar]
- Yang, Y.C.; Wei, M.C.; Huang, T.C. Optimisation of an Ultrasound-assisted Extraction Followed by RP-HPLC Separation for the Simultaneous Determination of Oleanolic Acid, Ursolic Acid and Oridonin Content in Rabdosia rubescens. Phytochem. Anal. 2012, 23, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.Y.; Xu, G.F.; Dai, L. Analysis on the accumulation of oridonin in different portions of Isodon rubescens (Hemsley) H. Hara. Bangladesh J. Bot. 2019, 48, 1193–1197. [Google Scholar]
- Heckathorn, S.A.; DeLucia, E.H.; Zielinski, R.E. The contribution of drought-related decreases in foliar nitrogen concentration to decreases in photosynthetic capacity during and after drought in prairie grasses. Physiol. Plant. 1997, 101, 173–182. [Google Scholar] [CrossRef]
- Wang, M.M.; Xu, Z.G.; Song, J.H.; Liu, X.Y.; Jiao, X.L. Effects of different mowing treatments and stubble heights on the compensatory growth and quality of lettuce (Lactuca sativa L.). J. Hortic. Sci. Biotechnol. 2018, 93, 537–544. [Google Scholar] [CrossRef]
- Meng, X.Y.; Wang, Z.; He, S.L.; Shi, L.Y.; Song, Y.L.; Lou, X.Y.; He, D. LED-Supplied Red and Blue Light Alters the Growth, Antioxidant Status, and Photochemical Potential of in Vitro-Grown Gerbera jamesonii Plantlets. Hortic. Sci. Technol. 2019, 37, 473–489. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, J.S.; He, C.X.; Wang, Q.R. Effects of light spectra and N-15 pulses on growth, leaf morphology, physiology, and internal nitrogen cycling in Quercus variabilis Blume seedlings. PLoS ONE 2021, 16, e0243954. [Google Scholar] [CrossRef]
- Wei, H.X.; Guo, P. Carbohydrate metabolism during new root growth in transplanted Larix olgensis seedlings: Post-transplant response to nursery-applied inorganic fertilizer and organic amendment. Iforest-Biogeosci. For. 2017, 10, 15–22. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Lasceve, G.; Leymarie, J.; Vavasseur, A. Alterations in light-induced stomatal opening in a starch-deficient mutant of Arabidopsis thaliana L deficient in chloroplast phosphoglucomutase activity. Plant Cell Environ. 1997, 20, 350–358. [Google Scholar] [CrossRef]
- Janecek, S.; Lanta, V.; Klimesova, J.; Dolezal, J. Effect of abandonment and plant classification on carbohydrate reserves of meadow plants. Plant Biol. 2011, 13, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.D.; Wei, X.; Zhao, W.B.; Li, J.X.; Yang, G.; Zhou, S.; Zhou, C.W.; Zhang, L.; Li, P.J.; Hou, S.; et al. Natural Bioactive Substances in Fruits of Aronia melanocarpa (Michx.) Elliott Exposed to Combined Light-Type, Chitosan Oligosaccharide, and Spent Mushroom Residue Treatments. Plants 2023, 12, 604. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cao, B.; Wang, Y.T.; Wei, Z.P.; Ye, J.F.; Wei, H.X. Spectral effect of streetlamps on urban trees: A simulated study on tissue water, nitrogen, and carbohydrate contents in maple and oak. PLoS ONE 2021, 16, e0248463. [Google Scholar] [CrossRef] [PubMed]
- Avendano-Abarca, V.H.; Alvarado-Camarillo, D.; Valdez-Aguilar, L.A.; Sanchez-Ortfz, E.A.; Gonzalez-Fuentes, J.A.; Cartmill, A.D. Response of Strawberry to the Substitution of Blue Light by Green Light in an Indoor Vertical Farming System. Agronomy 2023, 13, 99. [Google Scholar] [CrossRef]
- Wei, H.; Guo, P.; Zheng, H.; He, X.; Wang, P.; Ren, Z.; Zhai, C. Micro-scale heterogeneity in urban forest soils affects fine root foraging by ornamental seedlings of Buddhist pine and Northeast yew. Urban For. Urban Green. 2017, 28, 63–72. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, C.W.; Wu, Y.J. Dry mass input into fruits can be predicted by fine root morphology of pepper cultivars exposed to varied lighting spectra. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12930. [Google Scholar] [CrossRef]
- Yuan, W.; Cai, W.; Chen, Y.; Liu, S.; Dong, W.; Zhang, H.; Yu, G.; Chen, Z.; He, H.; Guo, W.; et al. Severe summer heatwave and drought strongly reduced carbon uptake in Southern China. Sci. Rep. 2016, 6, 18813. [Google Scholar] [CrossRef]
- Chen, H.; Quan, W.; Liu, H.; Ding, G. Effects of Suillus luteus and S. bovinus on the physiological response and nutrient absorption of Pinus massoniana seedlings under phosphorus deficiency. Plant Soil 2022, 471, 577–590. [Google Scholar] [CrossRef]
- Sima, Y.-H.; Yao, J.-M.; Hou, Y.-S.; Wang, L.; Zhao, L.-C. Variations of hydrogen peroxide and catalase expression in Bombyx eggs during diapause initiation and termination. Arch. Insect Biochem. Physiol. 2011, 77, 72–80. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Wang, X.; Li, Y.; Peng, F.; Wang, L. Regulation of POD activity by pelargonidin during vegetative growth in radish (Raphanus sativus L.). Sci. Hortic 2014, 174, 105–111. [Google Scholar] [CrossRef]
- Ding, X.G.; Li, X.C.; Qi, Y.; Zhao, Z.Y.; Sun, D.X.; Wei, H.X. Depth-Dependent C-N-P Stocks and Stoichiometry in Ultisols Resulting from Conversion of Secondary Forests to Plantations and Driving Forces. Forests 2021, 12, 1300. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Seedling Growth, Dry Mass Production (DM), and Quality | |||
|---|---|---|---|---|
| Height | RCD 1 | Oridonin | DM | |
| Mowing (M) | <0.0001 2 | 0.1489 | 0.0036 | 0.2246 |
| LED (L) | 0.0428 | 0.0625 | 0.0409 | 0.0052 |
| Drought (D) | 0.0007 | 0.0081 | 0.0013 | 0.0018 |
| M × L | 0.5187 | 0.3628 | 0.9790 | 0.0494 |
| M × D | 0.1142 | 0.1223 | 0.7481 | 0.3913 |
| L × D | 0.4407 | 0.3431 | 0.1193 | 0.2193 |
| M × L × D | 0.6022 | 0.3964 | 0.9688 | 0.7512 |
| Source | Species | Provincial Habitat | Sampling Time | Organ | Oridonin Content (% DW) |
|---|---|---|---|---|---|
| Guo et al. [41] | Rabdosia rubescens | Henan | Unknown | Whole plant | 0.550 |
| Guangxi | 0.540 | ||||
| Jiangxi | 0.590 | ||||
| Sichuan | 0.580 | ||||
| Cui et al. [42] | R. rubescens | Henan | July | Whole plant | 0.470 |
| August | 0.620 | ||||
| September | 0.630 | ||||
| October | 0.450 | ||||
| Yang et al. [43] | R. rubescens | Taiwan | Unknown | Whole plant | 0.468 |
| Harris et al. [34] | Isodon enaderianus QD | Guizhou | October and November | Leaves | 0.000 |
| I. enanderianus TR | Guizhou | 0.000 | |||
| I. henryi XN | Henan | 0.000 | |||
| I. japonicus XX | Henan | 0.752 | |||
| I. lophanthoides var. micranthus TR | Guizhou | 0.000 | |||
| I. rubescens HB | Henan | 0.394 | |||
| I. rubescens JY | Henan | 0.317 | |||
| I. rubescens QY | Henan | 0.256 | |||
| I. rubescens XX | Henan | 0.272 | |||
| I. rubescens YC | Hubei | 0.150 | |||
| I. enaderianus QD | Guizhou | August | 0.000 | ||
| I. enanderianus TR | Guizhou | 0.000 | |||
| I. henryi XN | Henan | 0.000 | |||
| I. japonicus XX | Henan | 0.211 | |||
| I. rubescens HB | Henan | 0.200 | |||
| I. rubescens JY | Henan | 0.289 | |||
| I. rubescens QY | Henan | 0.263 | |||
| I. rubescens XX | Henan | 0.174 | |||
| I. rubescens YC | Hubei | 0.160 | |||
| Jian et al. [44] | I. rubescens (HEMSLEY) H. Hara | Henan | Unknown | Root | 0.008 |
| Source of Variation | Physiological Parameters | ||||||
|---|---|---|---|---|---|---|---|
| Chlorophyll | Protein | Sugar | Starch | TC 1 | δ13C | TN 2 | |
| Mowing (M) | <0.0001 3 | <0.0001 | <0.0001 | <0.0001 | 0.0026 | 0.0005 | <0.0001 |
| LED (L) | 0.0262 | 0.0002 | 0.0019 | <0.0001 | 0.1865 | 0.0066 | 0.0050 |
| Drought (D) | 0.0005 | <0.0001 | 0.0009 | <0.0001 | 0.0110 | 0.1078 | 0.0031 |
| M × L | 0.6250 | 0.0784 | 0.5941 | 0.0292 | 0.7660 | 0.2914 | 0.8615 |
| M × D | 0.0260 | 0.7045 | 0.0886 | 0.0795 | 0.6220 | 0.1614 | 0.8613 |
| L × D | 0.1386 | 0.4164 | 0.1599 | 0.0329 | 0.5425 | 0.1264 | 0.5941 |
| M × L × D | 0.2129 | 0.6399 | 0.3737 | 0.5574 | 0.7867 | 0.7391 | 0.9806 |
| Source of Variation | Antioxidant Activity | ||
|---|---|---|---|
| CAT 1 | POD 2 | SOD 3 | |
| Mowing (M) | <0.0001 | 0.0047 | 0.0044 |
| LED (L) | 0.0180 | 0.0007 | 0.7400 |
| Drought (D) | 0.0126 | <0.0001 | <0.0001 |
| M × L | 0.4685 | 0.0852 | 0.8261 |
| M × D | 0.7877 | 0.6173 | 0.9715 |
| L × D | 0.8174 | 0.2702 | 0.7273 |
| M × L × D | 0.7202 | 0.6666 | 0.2622 |
| Variable | Model 1 1 | Model 2 2 | Model 3 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | SE 4 | p | Parameter | SE | p | Parameter | SE | p | |
| Intercept | −1.9291 | 1.2884 | 0.1436 | 0.2539 | 1.4838 | 0.8653 | −1.4182 | 1.2337 | 0.2589 |
| C13 5 | −0.1125 | 0.0388 | 0.0066 | −0.0757 | 0.0364 | 0.0464 | −0.0911 | 0.0368 | 0.0187 |
| CAT 6 | −0.0004 | 0.0002 | 0.0496 | −0.0004 | 0.0002 | 0.0176 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Meng, P.; Zhao, Y.; Zhang, J.; He, C.; Wang, Q.; Cai, J. Light-Emitting Diodes Modify Medicinal Quality of Mown Rabdosia rubescens, with Changes in Growth, Physiology, and Antioxidant Activity, under Drought Stress. Plants 2023, 12, 3189. https://doi.org/10.3390/plants12183189
Gao J, Meng P, Zhao Y, Zhang J, He C, Wang Q, Cai J. Light-Emitting Diodes Modify Medicinal Quality of Mown Rabdosia rubescens, with Changes in Growth, Physiology, and Antioxidant Activity, under Drought Stress. Plants. 2023; 12(18):3189. https://doi.org/10.3390/plants12183189
Chicago/Turabian StyleGao, Jun, Ping Meng, Yan Zhao, Jinsong Zhang, Chunxia He, Qirui Wang, and Jinfeng Cai. 2023. "Light-Emitting Diodes Modify Medicinal Quality of Mown Rabdosia rubescens, with Changes in Growth, Physiology, and Antioxidant Activity, under Drought Stress" Plants 12, no. 18: 3189. https://doi.org/10.3390/plants12183189