
Seasonal Variation in Cell Wall Composition and Carbohydrate Metabolism in the Seagrass Posidonia oceanica Growing at Different Depths

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Non-Structural Carbohydrates Content
2.1.1. Starch Content and Amylase Activity
2.1.2. Soluble Carbohydrates
2.2. Composition of Cell-Wall Polysaccharides
2.2.1. Non-Cellulosic Sugar Content
2.2.2. Cellulose Content
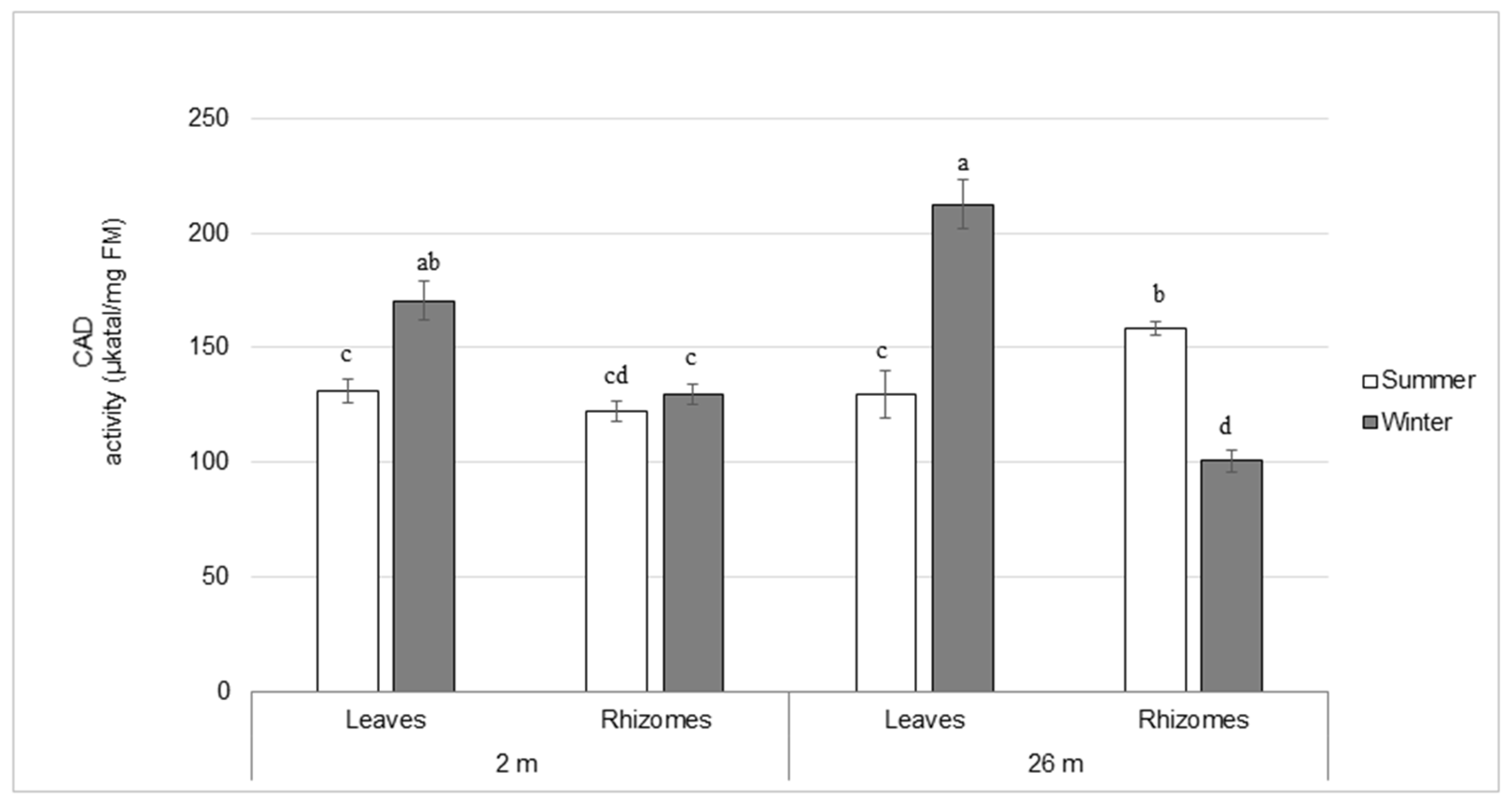
2.3. Lignin Composition and Enzyme Activities Involved in Lignin Biosynthesis
2.4. Boron Content
2.5. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sampling Conditions
4.2. Soluble Sugar Analyses
4.3. Starch Extraction and Solubilization and Amylase Activity
4.4. Extraction and Derivatization of Phenylalanine for GC–MS
4.5. Cell Wall Carbohydrate Composition
4.5.1. The Non-Cellulosic Monosaccharide Composition
4.5.2. Crystalline Cellulose Content
4.6. Lignin Content and Subunit Composition
4.7. PAL and CAD Enzyme Assays
4.7.1. Extraction and Assay of PAL
4.7.2. Oxidation of Cinnamyl Alcohols
4.8. Sample Preparation and Boron Analyses by ICP-OES
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pasqualini, V.; Pergent-Martini, C.; Clabaut, P.; Pergent, G. Mapping of Posidonia oceanica Using Aerial Photographs and Side Scan Sonar: Application of the Island of Corsica (France). Estuar. Coast. Shelf Sci. 1998, 47, 359–367. [Google Scholar] [CrossRef]
- Procaccini, G.; Buia, M.C.; Gambi, M.C.; Perez, M.; Pergent, G.; Pergent-Martini, C.; Romero, J. Seagrass Status and Extent Along the Mediterranean Coasts of Italy, France and Spain. In World Atlas of Seagrass: Present Status and Future Conservation; Green, E.P., Short, F.T., Spalding, M.D., Eds.; University of California Press: Berkeley, CA, USA, 2003; pp. 56–66. [Google Scholar]
- Borum, J.; Duarte, C.M.; Krause-Jensen, D.; Greve, T.M. European Seagrasses: An Introduction to Monitoring and Management. The EU project Monitoring & Management of European Seagrasses (M&MS). 2004, p. 61. Available online: http://www.seagrasses.org (accessed on 25 August 2023).
- Díaz-Almela, E.; Marbà, N.; Álvarez, E.; Santiago, R.; Holmer, M.; Grau, A.; Mirto, S.; Danovaro, R.; Petrou, A.; Argyrou, M.; et al. Benthic Input Rates Predict Seagrass (Posidonia oceanica) Fish Farm-Induced Decline. Mar. Pollut. Bull. 2008, 56, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Pergent, G.; Bazairi, H.; Bianchi, C.N.; Boudouresque, C.F.; Buia, M.C.; Calvo, S.; Clabaut, P.; Harmelin-Vivien, M.; Mateo, M.A.; Montefalcone, M.; et al. Climate Change and Mediterranean Seagrass Meadows: A Synopsis for Environmental Managers. Medit. Mar. Sci. 2014, 15, 462–473. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Ceccherelli, G. Epiphyte Assemblages of the Mediterranean Seagrass Posidonia oceanica: An Overview. Mar. Ecol. 2016, 37, 3–41. [Google Scholar] [CrossRef]
- Sogin, E.M.; Michellod, D.; Gruber-Vodicka, H.; Bourceau, P.; Geier, B.; Meier, D.V.; Seidel, M.; Ahmerkamp, S.; Schorn, S.; D’Angelo, G.; et al. Sugars Dominate the Seagrass Rhizosphere. Nat. Ecol. Evol. 2022, 6, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Serrano, O.; Lavery, P.S.; López-Merino, L.; Ballesteros, E.; Mateo, M.A. Location and Associated Carbon Storage of Erosional Escarpments of Seagrass Posidonia Mats. Front. Mar. Sci. 2016, 3, 42. [Google Scholar] [CrossRef]
- Apostolaki, E.T.; Caviglia, L.; Santinelli, V.; Cundy, A.B.; Tramati, C.D.; Mazzola, A.; Vizzini, S. The Importance of Dead Seagrass (Posidonia oceanica) Matte as a Biogeochemical Sink. Front. Mar. Sci. 2022, 9, 861998. [Google Scholar] [CrossRef]
- Touchette, B.W.; Burkholder, J.M. Overview of the Physiological Ecology of Carbon Metabolism in Seagrasses. J. Exp. Mar. Biol. Ecol. 2000, 250, 169–205. [Google Scholar] [CrossRef]
- Pirc, H. Seasonal Changes in Soluble Carbohydrates, Starch, and Energy Content in Mediterranean Seagrasses. Mar. Ecol. 1989, 10, 97–105. [Google Scholar] [CrossRef]
- Larkum, A.W.D.; Drew, E.A.; Ralph, P.J. Seagrasses: Biology, Ecology and Conservation; Springer: Berlin, Germany, 2006; p. 691. [Google Scholar]
- Olsen, J.L.; Rouzé, P.; Verhelst, B.; Lin, Y.-C.; Bayer, T.; Collen, J.; Dattolo, E.; De Paoli, E.; Dittami, S.; Maumus, F.; et al. The Genome of the Seagrass Zostera marina Reveals Angiosperm Adaptation to the Sea. Nature 2016, 530, 331–335. [Google Scholar] [CrossRef]
- Seifert, G.J. Nucleotide Sugar Interconversions and Cell Wall Biosynthesis: How to Bring the Inside to the Outside. Curr. Opin. Plant Biol. 2004, 7, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, L.; Classen, B. The Cell Wall of Seagrasses: Fascinating, Peculiar and a Blank Canvas for Future Research. Front. Plant Sci. 2020, 23, 588754. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, L.; van Erven, G.; Sinclair, E.A.; Duarte, C.M.; Kabel, M.A.; Classen, B. Profiling the Cell Walls of Seagrasses from A (Amphibolis) to Z (Zostera). BMC Plant Biol. 2022, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, L.; Shafee, T.; Johnson, K.L.; Bacic, A.; Classen, B. Arabinogalactan-Proteins of Zostera marina L. Contain Unique Glycan Structures and Provide Insight into Adaption Processes to Saline Environments. Sci. Rep. 2020, 10, 8232. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.S.; Landeira-Fernandez, A.M.; Valente, A.P.; Andrade, L.R.; Mourão, P.A. Occurrence of Sulfated Galactans in Marine Angiosperms: Evolutionary Implications. Glycobiology 2005, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gloaguen, V.; Brudieux, V.; Closs, B.; Barbat, A.; Krausz, P.; Sainte-Catherine, O.; Kraemer, M.; Maes, E.; Guérardel, Y. Structural Characterization and Cytotoxic Properties of an Apiose-Rich Pectic Polysaccharide Obtained from the Cell Wall of the Marine Phanerogam Zostera marina. J. Nat. Prod. 2010, 73, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, Y.; Khozhaenko, E.; Kovalev, V.; Khotimchenko, M. Cerium Binding Activity of Pectins Isolated from the Seagrasses Zostera marina and Phyllospadix iwatensis. Mar. Drugs 2012, 10, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.; Key, R.M.; Heuven, S.; van Lauvset, S.K.; Velo, A.; Lin, X.; Schirnick, C.; Kozyr, A.; Tanhua, T.; Hoppema, M.; et al. The Global Ocean Data Analysis Project Version 2 (Glodapv2)—An Internally Consistent Data Product for the World Ocean . Earth Syst. Sci. Data 2016, 8, 297–323. [Google Scholar] [CrossRef]
- Kaal, J.; Serrano, O.; José, C.; Rencoret, J. Radically Different Lignin Composition in Posidonia Species May Link to Differences in Organic Carbon Sequestration Capacity. Org. Geochem. 2018, 124, 247–256. [Google Scholar] [CrossRef]
- Rencoret, J.; Marques, G.; Serrano, O.; Kaal, J.; Martínez, A.T.; del Río, J.C.; Gutiérrez, A. Deciphering the Unique Structure and Acylation Pattern of Posidonia oceanica Lignin. ACS Sustain. Chem. Eng. 2020, 8, 12521–12533. [Google Scholar] [CrossRef]
- Kaal, J.; Serrano, O.; Nierop, K.G.J.; Schellekens, J.; Martínez Cortizas, A.; Mateo, M.A. Molcular Composition of Plant Parts and Sediment Organic Matter in a Mediterranean Seagrass (Posidonia oceanica) Mat. Aquat. Bot. 2016, 133, 50–61. [Google Scholar] [CrossRef]
- Touchette, B.W.; Burkholder, J.M. Effects of Temperature and Nitrate on Phosphomonoesterase Activities Between Carbon Source and Sink Tissues in Zostera marina L. J. Exp. Mar. Biol. Ecol. 2007, 342, 313–324. [Google Scholar] [CrossRef]
- Ruocco, M.; Entrambasaguas, L.; Dattolo, E.; Milito, A.; Marín-Guirao, L.; Procaccini, G. A King and Vassals’ Tale: Molcular Signatures of Clonal Integration in Posidonia oceanica Under Chronic Light Shortage. J. Ecol. 2021, 109, 294–312. [Google Scholar] [CrossRef]
- Dattolo, E.; Ruocco, M.; Brunet, C.; Lorenti, M.; Lauritano, C.; D’Esposito, D.; De Luca, P.; Sanges, R.; Mazzuca, S.; Procaccini, G. Response of the Seagrass Posidonia oceanica to Different Light Environments: Insights from a Combined Molcular and Photo-Physiological Study. Mar. Environ. Res. 2014, 101, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Barrote, I.; Costa, M.M.; Albano, S.; Santos, R. Physiological Responses of Zostera marina and Cymodocea nodosa to Light-Limitation Stress. PLoS ONE 2013, 8, e81058. [Google Scholar] [CrossRef] [PubMed]
- Enríquez, S.; Olivé, I.; Cayabyab, N.; Hedley, J.D. Structural Complexity Governs Seagrass Acclimatization to Depth with Relevant Consequences for Meadow Production, Macrophyte Diversity and Habitat Carbon Storage Capacity. Sci. Rep. 2019, 9, 14657. [Google Scholar] [CrossRef] [PubMed]
- Monnier, B.; Pergent, G.; Mateo, M.Á.; Clabaut, P.; Pergent-Martini, C. Quantification of Blue Carbon Stocks Associated with Posidonia oceanica Seagrass Meadows in Corsica (NW Mediterranean). Sci. Total Environ. 2022, 10, 155864. [Google Scholar] [CrossRef] [PubMed]
- Mulder, M.M.; Van der Hage, E.R.E.; Boon, J.J. Analytical in Source Pyrolytic Methylation Electron Impact Mass Spectrometry of Phenolic Acids in Biological Matrices. Phytochem. Anal. 1992, 3, 165–172. [Google Scholar] [CrossRef]
- Jun, S.Y.; Sattler, S.A.; Cortez, G.S.; Vermerris, W.; Sattler, S.E.; Kang, C. Biochemical and Structural Analysis of Substrate Specificity of a Phenylalanine Ammonia-lyase. Plant Physiol. 2018, 176, 1452–1468. [Google Scholar] [CrossRef]
- Collier, C.J.; Langlois, L.M.; McMahon, K.M.; Udy, J.; Rasheed, M.; Lawrence, E.; Carter, A.B.; Fraser, M.W.; McKenzie, L.J. What Lies Beneath: Predicting Seagrass Below-Ground Biomass from Above-Ground Biomass, Environmental Conditions and Seagrass Community Composition. Ecol. Indic. 2021, 121, 107156. [Google Scholar] [CrossRef]
- Charras, Q.; Charlie, M.; Semchonok, D.; Guillemain, G.; Rey, P.; Lebrun, R.; Laganier, H.; Siponen, M.; Farhan, A.; Hienerwadel, R.; et al. The Remarkable Photosynthesis of the Seagrass Posidonia oceanica: Analysis of the Photosynthetic Complexes and Isolation of Abundant Blue Shifted Large-PSI-LHCII Supercomplexes. In Proceedings of the Poster and Oral Presentation, Nordic Congress of Photosynthesis (NPC15), Gothenburg, Sweden, 5–8 October 2021. [Google Scholar]
- Procaccini, G.; Ruocco, M.; Marín-Guirao, L.; Dattolo, E.; Brunet, C.; D’Esposito, D.; Lauritano, C.; Mazzuca, S.; Anna Serra, I.; Bernardo, L.; et al. Depth-Specific Fluctuations of Gene Expression and Protein Abundance Modulate the Photophysiology in the Seagrass Posidonia oceanica. Sci. Rep. 2017, 7, 42890. [Google Scholar] [CrossRef] [PubMed]
- Dattolo, E.; Marín-Guirao, L.; Ruiz, J.M.; Procaccini, G. Long-Term Acclimation to Reciprocal Light Conditions Suggests Depth-Related Selection in the Marine Foundation Species Posidonia oceanica. Ecol. Evol. 2017, 7, 1148–1164. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Box, A.; Terrados, J.; Deudero, S.; Pons, A. Antioxidant Response of the Seagrass Posidonia Oceanica When Epiphytized by the Invasive Macroalgae Lophocladia Lallemandii. Mar. Environ. Res. 2008, 66, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.M.; Barrote, I.; Silva, J.; Olivé, I.; Alexandre, A.; Albano, S.; Santos, R. Epiphytes Modulate Posidonia Oceanica Photosynthetic Production, Energetic Balance, Antioxidant Mechanisms, and Oxidative Damage. Front. Mar. Sci. 2015, 2, 111. [Google Scholar] [CrossRef]
- Griffiths, L.L.; Melvin, S.D.; Connolly, R.M.; Pearson, R.M.; Brown, C.J. Metabolomic Indicators for Low-Light Stress in Seagrass. Ecol. Indic. 2020, 114, 106316. [Google Scholar] [CrossRef]
- Govers, L.L.; Suykerbuyk, W.; Hoppenreijs, J.H.T.; Giesen, K.; Bouma, T.J.; van Katwijk, M.M. Rhizome Starch as Indicator for Temperate Seagrass Winter Survival. Ecol. Indic. 2015, 49, 53–60. [Google Scholar] [CrossRef]
- Premarathne, C.; Jiang, Z.; He, J.; Fang, Y.; Chen, Q.; Cui, L.; Wu, Y.; Liu, S.; Chunyu, Z.; Vijerathna, P.; et al. Low Light Availability Reduces the Subsurface Sediment Carbon Content in Halophila beccarii from the South China Sea. Front. Plant Sci. 2021, 12, 664060. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Kulkarni, A.R.; Backe, J.; Boyd, M.; O’Neill, M.A.; York, W.S. Structural Diversity of Xylans in the Cell Walls of Monocots. Planta 2016, 244, 589–606. [Google Scholar] [CrossRef]
- Zhao, X.; Moates, G.K.; Wellner, N.; Collins, S.R.; Coleman, M.J.; Waldron, K.W. Chemical Characterisation and Analysis of the Cell Wall Polysaccharides of Duckweed (Lemna minor). Carbohydr. Polym. 2014, 111, 410–418. [Google Scholar] [CrossRef]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving Views of Pectin Biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef]
- Sowinski, E.E.; Gilbert, S.; Lam, E.; Carpita, N.C. Linkage Structure of Cell-Wall Polysaccharides from Three Duckweed Species. Carbohydr. Polym. 2019, 223, 115119. [Google Scholar] [CrossRef]
- Pfeifer, L. Enigmatic Apiogalacturonans—What Do We Know about This Group of Pectic Polysaccharides? Annu. Plant Rev. Online. 2023, 6, 1–30. [Google Scholar] [CrossRef]
- Pagliuso, D.; Grandis, A.; Igarashi, E.S.; Lam, E.; Buckeridge, M.S. Correlation of Apiose Levels and Growth Rates in Duckweeds. Front. Chem. 2018, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, C.; Pan, Z.; Liu, Y.; Peng, S. Boron Deficiency in Woody Plants: Various Responses and Tolerance Mechanisms. Front. Plant Sci. 2015, 6, 916. [Google Scholar] [CrossRef]
- Dell, B.; Huang, L. Physiological Response of Plants to Low Boron. Plant Soil 1997, 193, 103–120. [Google Scholar] [CrossRef]
- O’Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and Function of a Borate Cross-Linked Cell Wall Pectic Polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Masaoka, N.; Ishii, T.; Satoh, S. A Pectin Glucuronyltransferase Gene is Essential for Intercellular Attachment in the Plant Meristem. Proc. Natl. Acad. Sci. USA 2002, 99, 16319–16324. [Google Scholar] [CrossRef]
- Karlen, S.D.; Free, H.C.A.; Padmakshan, D.; Smith, B.G.; Ralph, J.; Harris, P.J. Commelinid Monocotyledon Lignins are Acylated By p-Coumarate. Plant Physiol. 2018, 177, 513–521. [Google Scholar] [CrossRef]
- Vermerris, W.; Boon, J.J. Tissue-Specific Patterns of Lignification are Disturbed in the brown midrib2 Mutant of Maize (Zea mays L.). J. Agric. Food Chem. 2001, 49, 721–728. [Google Scholar] [CrossRef]
- Duran Garzon, C.; Lequart, M.; Charras, Q.; Fournet, F.; Bellenger, L.; Sellier-Richard, H.; Giauffret, C.; Vermerris, W.; Domon, J.M.; Rayon, C. The Maize Low-Lignin brown midrib3 Mutant Shows Pleiotropic Effects on Photosynthetic and Cell Wall Metabolisms in Response to Chilling. Plant Physiol. Biochem. 2022, 184, 75–86. [Google Scholar] [CrossRef]
- De Souza, A.P.; Leite, D.C.C.; Pattathil, S.; Hanh, M.G.; Buckeridge, M.S. Composition and structure of sugarcane cell wall polysaccharides: Implications for second-generation bioethanol production. Bioenergy Res. 2013, 6, 564–579. [Google Scholar] [CrossRef]
- Cui, K.H.; Peng, S.B.; Xing, Y.Z.; Xu, C.G.; Yu, S.B.; Zhang, Q. Molcular Dissection of Seedling-Vigor and Associated Physiological Traits in Rice. Theor. Appl. Genet. 2002, 105, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Quéro, A.; Jousse, C.; Lequart-Pillon, M.; Gontier, E.; Guillot, X.; Courtois, B.; Courtois, J.; Pau-Roblot, C. Improved Stability of TMS Derivatives for the Robust Quantification of Plant Polar Metabolites by Gas Chromatography-Mass Spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2014, 970, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Foster, C.E.; Martin, T.M.; Pauly, M. Comprehensive Compositional Analysis of Plant Cell Walls (Lignocellulosic biomass) Part II: Carbohydrates. J. Vis. Exp. 2010, 37, e1837. [Google Scholar] [CrossRef]
- Foster, C.E.; Martin, T.M.; Pauly, M. Comprehensive Compositional Analysis of Plant Cell Walls (Lignocellulosic biomass) Part I: Lignin. J. Vis. Exp. 2010, 37, 1745. [Google Scholar] [CrossRef]
- Domon, J.M.; Baldwin, L.; Acket, S.; Caudeville, E.; Arnoult, S.; Zub, H.; Gillet, F.; Lejeune-Hénaut, I.; Brancourt-Hulmel, M.; Pelloux, J.; et al. Cell Wall Compositional Modifications of Miscanthus Ecotypes in Response to Cold Acclimation. Phytochemistry 2013, 85, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, C.; Monties, B.; Rolando, C. Thioacidolysis of Lignin: Comparison with Acidolysis. J. Wood Chem. Technol. 1985, 2, 277–292. [Google Scholar] [CrossRef]
- Grossman, A.B.; Rice, K.C.; Vermerris, W. Lignin Solvated in Zwitterionic Good’s Buffers Displays Antibacterial Synergy Against Staphylococcus aureus. J. Appl. Polym. Sci. 2020, 137, e49107. [Google Scholar] [CrossRef]
- Ralph, J.; Hatfield, R.D. Pyrolysis-GC-MS Characterization of Forage Materials. J. Agric. Food Chem. 1991, 39, 1426–1437. [Google Scholar] [CrossRef]
- Sattler, S.E.; Saballos, A.; Xin, Z.; Funnell-Harris, D.L.; Vermerris, W.; Pedersen, J.F. Characterization of Novel Sorghum brown midrib Mutants From an EMS-Mutagenized Population. G3 2014, 4, 2115–2124. [Google Scholar] [CrossRef]
- Wyrambick, D.; Grisebach, H. Purification and Properties of Isoenzymes of Cinnamyl Alcohol Dehydrogenase from Soybean. Eur. J. Biochem. 1975, 59, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25, 1–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2 m | ||||
|---|---|---|---|---|
| Summer | Winter | |||
| Leaves | Rhizomes | Leaves | Rhizomes | |
| Trait | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Cellulose (% of DCW) | 24.2 ± 3.4 bc | 26.2 ± 3.9 b | 17.7 ± 2.8 de | 19.1 ± 3.2 d |
| Non-cellulosic monosaccharide (mol%) | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Ara | 3.6 ± 0.7 cd | 16.2 ± 1.5 a | 2.8 ± 0.3 d | 13.8 ± 0.8 b |
| Gal | 2.6 ± 0.4 c | 8.2 ± 1 b | 2.7 ± 0.3 c | 8.1 ± 0.7 b |
| Xyl | 43 ± 1 ab | 32 ± 1.6 c | 39.1 ± 1.3 b | 32.9 ± 1 c |
| Api | 25 ± 1.6 b | 12.4 ± 0.7 e | 30.7 ± 1.6 a | 14.9 ± 1 d |
| GalUA | 7.9 ± 1 cd | 9.5 ± 2.4 abc | 9.2 ± 1.1 bc | 11.3 ± 1.3 ab |
| GlcUA | 0.9 ± 0.2 bc | 1.3 ± 0.2 a | 0.8 ± 0.1 c | 0.8 ± 0.2 c |
| 26 m | ||||
| Summer | Winter | |||
| Leaves | Rhizomes | Leaves | Rhizomes | |
| Trait | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Cellulose (% of DCW) | 32.4 ± 3.9 a | 35 ± 4.2 a | 15.7 ± 2.9 e | 22.8 ± 2 c |
| Non-cellulosic monosaccharide (mol%) | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Ara | 3.1 ± 0.3 d | 14.1 ± 0.8 ab | 4.3 ± 0.2 c | 14.3 ± 1.5 b |
| Gal | 2.4 ± 0.5 cd | 8 ± 0.9 b | 1.8 ± 0.1 d | 10.7 ± 1 a |
| Xyl | 40.4 ± 2.1 b | 31.5 ± 0.7 c | 46.5 ± 1.4 a | 32.3 ± 1.1 c |
| Api | 28.2 ± 1.2 a | 14.4 ± 1.2 d | 24.5 ± 0.6 b | 18.5 ± 1.3 c |
| GalUA | 8.5 ± 1 cd | 12.3 ± 1.5 a | 6.7 ± 0.3 d | 8.5 ± 1.2 cd |
| GlcUA | 1.2 ± 0.1 a | 0.8 ± 0.2 c | 1.2 ± 0.2 ab | 0.9 ± 0.2 c |
| 2 m | ||||
|---|---|---|---|---|
| Summer | Winter | |||
| Leaves | Rhizomes | Leaves | Rhizomes | |
| Trait | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Lignin (% of DCW) | 8.8 ± 0.8 f | 13.8 ± 0.7 d | 10.3 ± 0.9 e | 16.4 ± 0.7 b |
| Phenylpropanoid units (%) | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| H | tr * | 10.4 ± 1.1 bc | tr * | 12.4 ± 0.5 b |
| G | 24.7 ± 4.4 d | 62.9 ± 2.4 ab | 21.9 ± 4.2 d | 65.6 ± 0.8 a |
| S | 74.7 ± 3.8 b | 26.7 ± 1.6 c | 78 ± 4.2 ab | 22 ± 1.3 d |
| S/G | 3.1 ± 0.7 b | 0.4 ± 0.04 cd | 3.7 ± 0.8 b | 0.3 ± 0.02 e |
| 26 m | ||||
| Summer | Winter | |||
| Leaves | Rhizomes | Leaves | Rhizomes | |
| Trait | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Lignin (% of DCW) | 7 ± 0.4 g | 20.9 ± 0.8 a | 15.4 ± 1.3 c | 12 ± 0.8 d |
| Phenylpropanoid units (%) | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| H | tr * | 15.5 ± 3 a | tr * | 15.4 ± 2.1 a |
| G | 22.8 ± 4.3 d | 51.3 ± 0.4 c | 13.5 ± 2.3 e | 61.3 ± 3.7 bc |
| S | 76.3 ± 2.7 b | 33.2 ± 0.6 c | 85.5 ± 1.2 a | 23.3 ± 2.3 d |
| S/G | 3.5 ± 0.9 b | 0.6 ± 0.02 c | 6.5 ± 1.2 a | 0.4 ± 0.1 de |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismael, M.; Charras, Q.; Leschevin, M.; Herfurth, D.; Roulard, R.; Quéro, A.; Rusterucci, C.; Domon, J.-M.; Jungas, C.; Vermerris, W.; et al. Seasonal Variation in Cell Wall Composition and Carbohydrate Metabolism in the Seagrass Posidonia oceanica Growing at Different Depths. Plants 2023, 12, 3155. https://doi.org/10.3390/plants12173155
Ismael M, Charras Q, Leschevin M, Herfurth D, Roulard R, Quéro A, Rusterucci C, Domon J-M, Jungas C, Vermerris W, et al. Seasonal Variation in Cell Wall Composition and Carbohydrate Metabolism in the Seagrass Posidonia oceanica Growing at Different Depths. Plants. 2023; 12(17):3155. https://doi.org/10.3390/plants12173155
Chicago/Turabian StyleIsmael, Marwa, Quentin Charras, Maïté Leschevin, Damien Herfurth, Romain Roulard, Anthony Quéro, Christine Rusterucci, Jean-Marc Domon, Colette Jungas, Wilfred Vermerris, and et al. 2023. "Seasonal Variation in Cell Wall Composition and Carbohydrate Metabolism in the Seagrass Posidonia oceanica Growing at Different Depths" Plants 12, no. 17: 3155. https://doi.org/10.3390/plants12173155