
Humic Substances Isolated from Recycled Biomass Trigger Jasmonic Acid Biosynthesis and Signalling
 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Humic Acids Characteristics
2.2. Plant Phenotypes and Growth
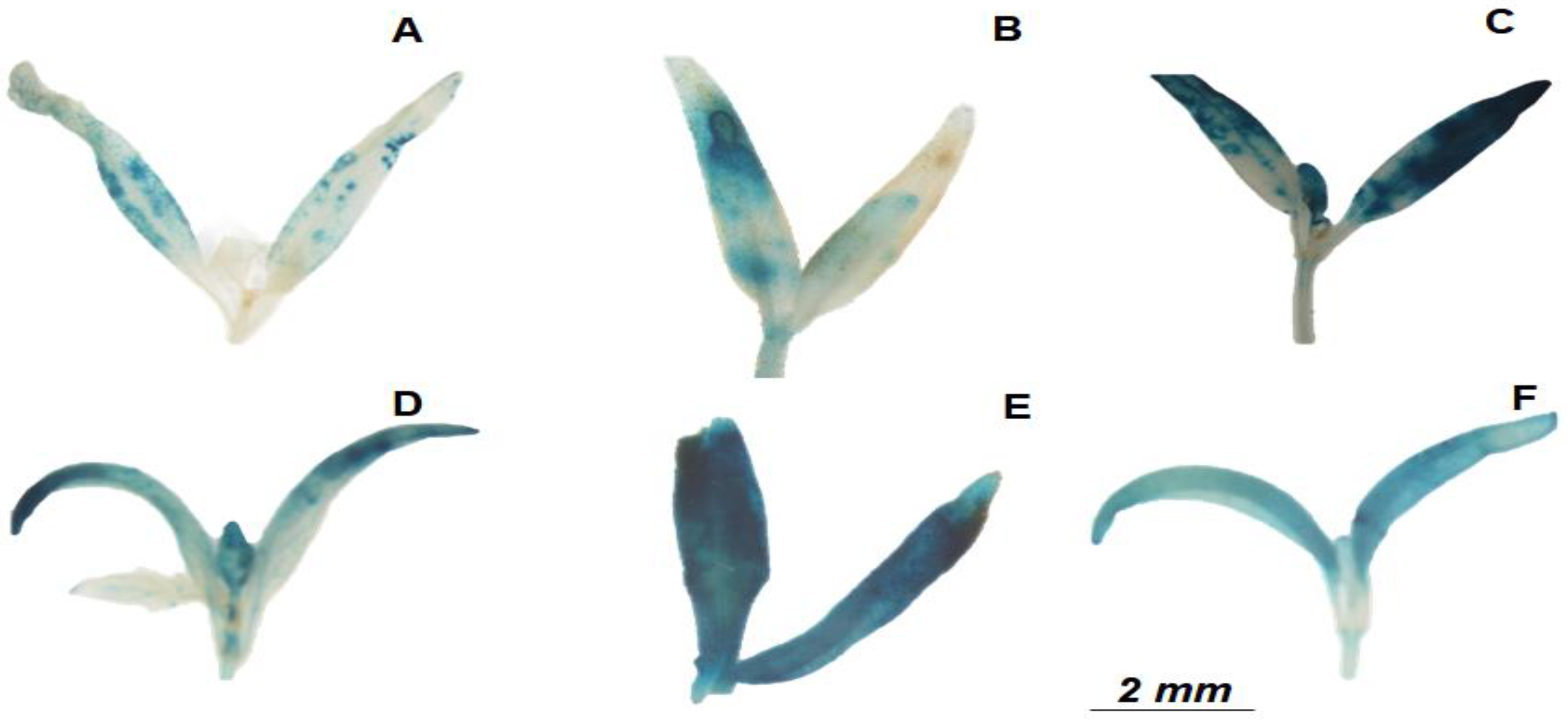
2.3. JERE::GUS Gene Reporter
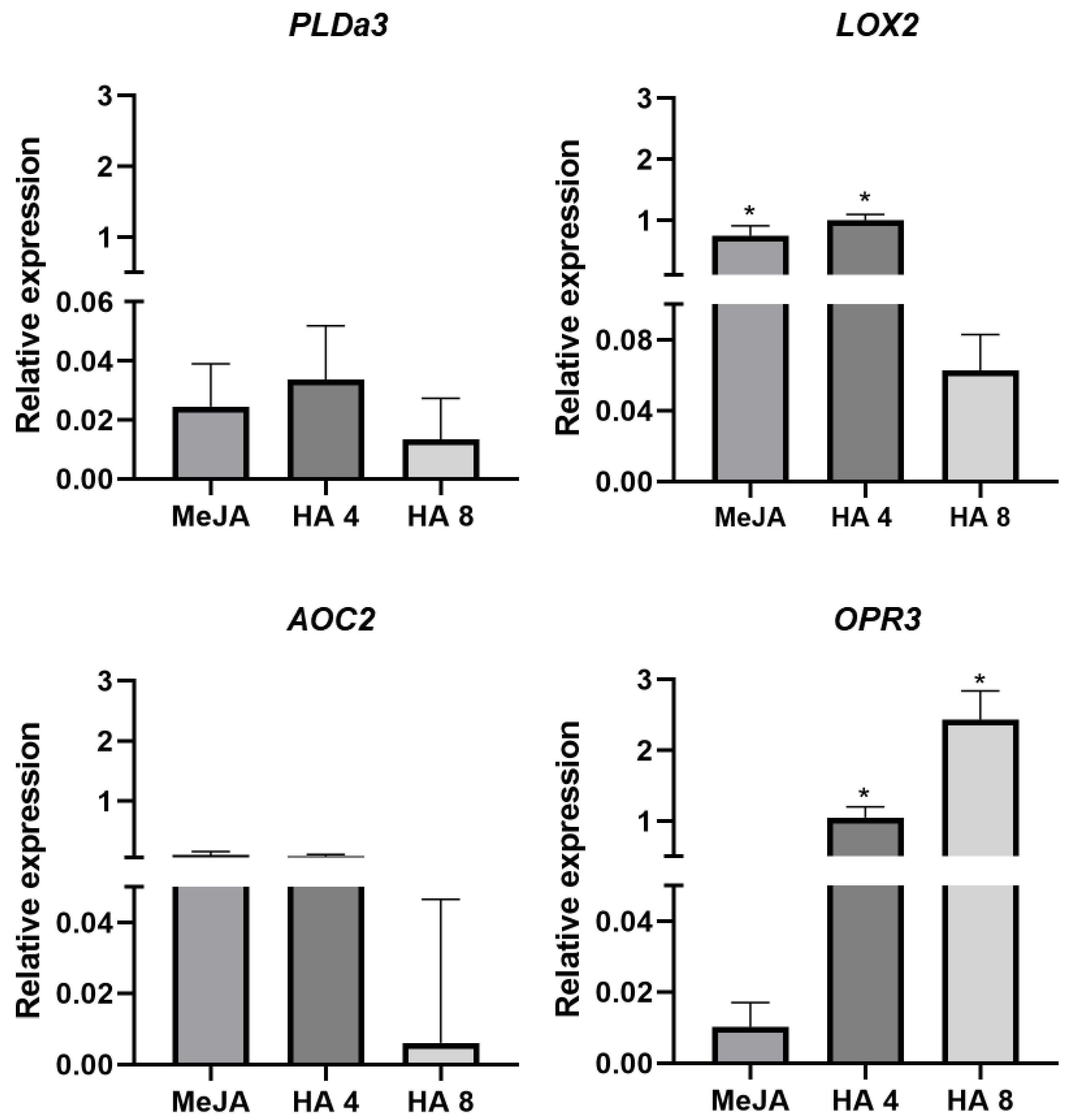
2.4. JA Synthesis and Signalling
3. Discussion
4. Materials and Methods
4.1. Vermicompost Production
4.2. Extraction and Purification of Humic Matter from Vermicompost
4.3. Humic Acids Characteristics
4.4. Plant Assay
4.5. Differential Transcription Level of Genes with RT-qPCR
4.6. Measurement of Trichome Density and Number
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devi, P.I.; Manjula, M.; Bhavani, R.V. Agrochemicals, environment, and human health. Ann. Rev. Environ. Res. 2022, 47, 399–421. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Canellas, N.A.; Jindo, K.; Rosa, R.C.C.; Piccolo, A. Challenge of transition: The history of a case study involving tropical fruits polyculture stimulated by humic acids and plant-growth promoting bacteria. Chem. Biol. Technol. Agric. 2022, 9, 76. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hort. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Li, J.; Van Gerrewey, T.; Geelen, D. A Meta-analysis of biostimulant yield effectiveness in field trials. Front. Plant Sci. 2022, 13, 836702. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef]
- Zaller, J.G. Vermicompost as a substitute for peat in potting media: Effects on germination, biomass allocation, yields and fruit quality of three tomato varieties. Sci. Hort. 2007, 112, 191–199. [Google Scholar] [CrossRef]
- Bamdad, H.; Papari, S.; Lazarovits, G.; Berruti, F. Soil amendments for sustainable agriculture: Microbial organic fertilizers. Soil Use Manag. 2022, 38, 94–120. [Google Scholar] [CrossRef]
- Padmavathiamma, P.K.; Li, L.Y.; Kumari, U.R. An experimental study of vermi-biowaste composting for agricultural soil improvement. Bioresour. Technol. 2008, 99, 1672–1681. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Edwards, C.A.; Lee, S.; Byrne, R. Effects of humic acids from vermicomposts on plant growth. Eur. J. Soil Biol. 2006, 42, S65–S69. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Olivares, F.L.; Novotny, E.H.; Dobbss, L.B.; Martinez-Balmori, D.; Santos-Júnior, L.G.; Chagas, J.G.; Façanha, A.R.; Canellas, L.P. Bioactivity of humic acids isolated from vermicomposts at different maturation stages. Plant Soil 2013, 362, 161–174. [Google Scholar] [CrossRef]
- Aquino, A.M.; Canellas, L.P.; da Silva, A.P.S.; Canellas, N.O.A.; Lima, L.S.; Olivares, F.L.; Piccolo, A.; Spaccini, R. Evaluation of molecular properties of humic acids from vermicompost by 13C-CPMAS-NMR spectroscopy and thermochemolysis–GC–MS. J. Ann. App. Pyrol. 2019, 141, 104634. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Ertani, A. Hormone-like activity of the soil organic matter. Appl. Soil Ecol. 2018, 123, 517–520. [Google Scholar] [CrossRef]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. A meta-analysis and review of plant-growth response to humic substances: Practical implications for agriculture. Adv. Agronom. 2014, 124, 37–89. [Google Scholar]
- Souza, A.C.; Olivares, F.L.; Peres, L.E.P.; Piccolo, A.; Canellas, L.P. Plant hormone cross-talk mediated by humic acids. Chem. Biol. Technol. Agric. 2022, 9, 29. [Google Scholar] [CrossRef]
- Canellas, L.P.; Canellas, N.O.A.; Irineu, L.E.S.D.S.; Olivares, F.L.; Piccolo, A. Plant chemical priming by humic acids. Chem. Biol. Technol. Agric. 2020, 7, 1–17. [Google Scholar] [CrossRef]
- Silva, R.M.; Canellas, L.P. Organic matter in the pest and plant disease control: A meta-analysis. Chem. Biol. Technol. Agric. 2022, 9, 70. [Google Scholar]
- Tiwari, S.; Lata, C.; Singh Chauhan, P.; Prasad, V.; Prasad, M. A Functional Genomic Perspective on Drought Signalling and its Cross-talk with Phytohormone-mediated Signalling Pathways in Plants. Curr. Genom. 2017, 18, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross talk between gibberellin and cytokinin: The Arabidopsis GA response inhibitor SPINDLY plays a positive role in cytokinin signaling. Plant Cell 2005, 171, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Bravo, R.; Klinkhamer, P.G.L.; Leiss, K.A. Induction of defenses associated with jasmonic acid by Thrips alters the suitability of the host for cospecifics and correlates with the increase in the density of trichomes in tomato. Plants Cell Physiol. 2017, 58, 622–634. [Google Scholar] [CrossRef]
- Azad, H.; Fazeli-Nasab, B.; Sobhanizade, A. A study into the effect of jasmonic and humic acids on some germination characteristics of rosselle (Hibiscus sabdariffa) Seed under Salinity Stress. Iran. J. Seed Res. 2017, 4, 1–18. [Google Scholar] [CrossRef]
- Ali, A.Y.A.; Ibrahim, M.E.H.; Zhou, G.; Nimir, N.E.A.; Jiao, X.; Zhu, G.; Lu, H. Ameliorative effects of jasmonic acid and humic acid on antioxidant enzymes and salt tolerance of forage sorghum under salinity conditions. Agron. J. 2019, 111, 3099–3108. [Google Scholar] [CrossRef]
- Ali, A.Y.; Ibrahim, M.E.; Zhou, G.; Nimir, N.E.; Jiao, X.; Zhu, G.; Elsiddig, A.M.; Suliman, M.S.; Elradi, S.B.; Yue, W. Exogenous jasmonic acid and humic acid increased salinity tolerance of sorghum. Agron. J. 2020, 112, 871–884. [Google Scholar] [CrossRef]
- Bijanzadeh, E.; Emam, Y.; Pessarakli, M. Biochemical responses of triticale under water stress (X Triticosecalewittmack) to humic acid and jasmonic acid. J. Plant. Nutr. 2020, 44, 252–269. [Google Scholar] [CrossRef]
- De Hita, D.; Fuentes, M.; Fernández, V.; Zamarreño, A.M.; Olaetxea, M.; García-Mina, J.M. Discriminating the short-term action of root and foliar application of humic acids on plant growth: Emerging role of jasmonic acid. Front. Plant Sci. 2020, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Funct. Plant Biol. Rev. 2019, 46, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Strnad, M. Jasmonates: News on Occurrence, Biosynthesis, Metabolism and Action of an Ancient Group of Signaling Compounds. Int. J. Mol. Sci. 2018, 19, 2539. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, X.; Gao, J.; Wang, P.; Xu, X.; Liu, Z. A buckwheat (Fagopyrum esculentum) DRE-binding transcription factor gene, FeDREB1, enhances freezing and drought tolerance of transgenic Arabidopsis. Plant Mol. Biol. Rep. 2015, 33, 1510–1525. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Olivares, F.L.; Novotny, E.H.; Canellas, L.P. Changes in metabolic profiling of sugarcane leaves induced by endophytic diazotrophic bacteria and humic acids. PeerJ 2018, 6, e5445. [Google Scholar] [CrossRef] [PubMed]
- Canellas, N.A.; Olivares, F.L.; Canellas, L.P. Metabolite fingerprints of maize and sugarcane seedlings: Searching for markers after inoculation with plant growth-promoting bacteria in humic acids. Chem. Biol. Technol. Agric. 2019, 6, 14. [Google Scholar] [CrossRef]
- Santos-Jiménez, J.L.; Montebianco, C.B.; Olivares, F.L.; Canellas, L.P.; Barreto-Bergter, E.; Rosa, R.C.C.; Vaslin, M.F.S. Passion fruit plants treated with biostimulants induce defense-related and phytohormone-associated genes. Plant Gene 2022, 30, 100357. [Google Scholar] [CrossRef]
- da Silva, S.F.; Spaccini, R.; Mazzei, P.; de Rezende, C.E.; Canellas, L.P. Changes in water-extractable organic matter in tropical forest and agricultural soils as detected by the DRIFT spectroscopy technique. Land Degrad. Dev. 2021, 32, 4755–4767. [Google Scholar] [CrossRef]
- Canellas, L.P.; Canellas, N.O.A.; da Silva, R.M.; Spaccini, R.; Mota, G.P.; Olivares, F.L. Biostimulants Using Humic Substances and Plant-Growth-Promoting Bacteria: Effects on Cassava (Manihot esculentus) and Okra (Abelmoschus esculentus) Yield. Agronomy 2022, 13, 80. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 3. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Ali, A.Y.A.; Zhou, G.; Elsiddig, A.M.; Zhu, G.; Meng, T.; Jiao, X.; Ahmed, I.; Ibrahim Salih, E.G.; Ibrahim, M.E.H. Effects of jasmonic acid in foliar spray and an humic acid amendment to saline soils on forage sorghum plants’ growth and antioxidant defense system. PeerJ 2022, 10, e13793. [Google Scholar] [CrossRef]
- Harwood, J.L. Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta 1996, 1301, 7–56. [Google Scholar] [CrossRef]
- Bonaventure, G.; Gfeller, A.; Proebsting, W.M.; Hörtensteiner, S.; Chételat, A.; Martinoia, E.; Farmer, E.E. A gain-of-function allele of TPC1 activates oxylipin biogenesis after leaf wounding in Arabidopsis. Plant 2007, 49, 889–898. [Google Scholar] [CrossRef]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef]
- Ramos, A.C.; Olivares, F.L.; Silva, L.S.; Aguiar, N.O.; Canellas, L.P. Humic matter elicits proton and calcium fluxes and signalling dependent on Ca2+-dependent protein kinase (CDPK) at early stages of lateral plant root development. Chem. Biol. Technol. Agric. 2015, 1, 1–12. [Google Scholar]
- Pauwels, L.; Inzé, D.; Goossens, A. Jasmonate-inducible gene: What does it mean? Trends Plant Sci. 2009, 14, 87–91. [Google Scholar] [CrossRef]
- Xie, D.-X.; Feys, B.; James, S.; Nieto-Rostro, M.; Turner, J. COI1: Na Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 1998, 280, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Yan, J.; Xie, D. Comparison of phytohormone signaling mechanisms. Curr. Opin. Plant Biol. 2012, 15, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. JAZ repressors and the orchestration of phytohormone cross-talk. Trends Plant Sci. 2012, 17, 22–31. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, J.; Zhai, Q.; Zhou, W.; Qi, L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H.; Qi, J.; et al. The basic helix–loop–helix transcription factor MYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef]
- Robaglia, C.; Thomas, M.; Meyer, C. Sensing nutrient and energy status by SnRK1 and TOR kinases. Curr. Opin. Plant. Biol. 2012, 15, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Lee, G.; Howe, G. Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato. Proc. Natl. Acad. Sci. USA 2002, 99, 6416–6421. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Botton, A.; Vaccaro, S.; Vezzaro, A.; Quaggiotti, S.; Nardi, S. Humic substances affect Arabidopsis physiology by altering the expression of genes involved in primary metabolism, growth and development. Environ. Exp. Bot. 2011, 74, 45–55. [Google Scholar] [CrossRef]
- Baía, D.C.; Olivares, F.L.; Zandonadi, D.B.; Soares, C.P.; Spaccini, R.; Canellas, L.P. Humic acids trigger the weak acids stress response in maize seedlings. Chem. Biol. Technol. Agric. 2020, 7, 31. [Google Scholar] [CrossRef]
- Wang, F.; Wu, N.; Zhang, L.; Ahammed, G.J.; Chen, X.; Xiang, X.; Zhou, J.; Xia, X.; Shi, K.; Yu, J.; et al. Light Signaling light-dependent regulation of photoinhibition e850 photoprotection in tomato. Plant Physiol. 2018, 176, 1311–1326. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Goossens, A.; Lacchini, E. Jasmonate: A hormone of primary importance for plant metabolism. Curr. Opin Plant Biol. 2022, 67, 102197. [Google Scholar] [CrossRef]
- Grunewald, W.; Vanholme, B.; Pauwels, L.; Plovie, E.; Inze, D.; Gheysen, G.; Goossens, A. Expression of the Arabidopsis jasmonate signalling repressor JAZ1/TIFY10A is stimulated by auxin. EMBO Rep. 2009, 10, 923–928. [Google Scholar] [CrossRef]
- Reeves, P.H.; Ellis, C.M.; Ploense, S.E.; Wu, M.-F.; Yadav, V.; Tholl, D.; Chételat, A.; Haupt, I.; Kennerley, B.J.; Hodgens, C.; et al. A regulatory network for coordinated flower maturation. PLoS Genet. 2012, 8, e1002506. [Google Scholar] [CrossRef] [PubMed]
- Tabata, R.; Ikezaki, M.; Fujibe, T.; Aida, M.; Tian, C.-E.; Ueno, Y.; Yamamoto, K.T.; Machida, Y.; Nakamura, K.; Ishiguro, S. Arabidopsis AUXIN RESPONSE FACTOR6 and 8 regulate jasmonic acid biosynthesis and floral organ development via repression of class 1 KNOX genes. Plant Cell Physiol. 2010, 51, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Tooker, J.; Peiffer, M.; Chung, S.; Felton, G. Role of trichomes in defense against herbivores: Comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 2012, 236, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Tissier, A. Glandular trichomes: What comes after expressed sequence tags? Plant J. 2012, 70, 51–68. [Google Scholar] [CrossRef]
- Schiavon, M.; Pizzeghello, D.; Muscolo, A.; Vaccaro, S.; Francioso, O.; Nardi, S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J. Chem. Ecol. 2010, 36, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Fan, J.; Yang, P.; Wang, Z.; Opiyo, S.O.; Mackey, D.; Xia, Y. Involvement of Arabidopsis acyl carrier protein 1 in PAMP-triggered immunity. Mol. Plant-Microbe Int. 2022, 35, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Pino, L.E.; Lombardi-Crestana, S.; Azevedo, M.S.; Scotton, D.C.; Borgo, L.; Quecini, V.; Figueira, A.; Peres, L.E.P. The Rg1 allele as a valuable tool for genetic transformation of the tomato ‘Micro-Tom’ model system. Plant Methods 2010, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a DRIFT (cm−1) | |||||||||
| 2945 | 1660 | 1590 | 1460 | 1260 | 1100 | 1030 | |||
| CH2 alkyl chains | COOH fatty acids | C=C aromatic units | CH2 alkyl chains | C-O alkyl acids/aromatic ehter | C-O phenols/polysaccahrides | C-O carbohydrates | |||
| b 13C CPMAS NMR (ppm) | |||||||||
| C=O 190-160 | O-aryl-C 160-145 | Aromatic-C 145-110 | O-alkyl-C 110-60 | CH3O/CN 60-45 | Alkyl-C 45-0 | HB | A/OA | Ar | LR |
| 6.5 | 8.4 | 20.3 | 28.3 | 16.7 | 19.8 | 1.3 | 0.7 | 28.7 | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, R.M.; Peres, A.N.A.; Peres, L.E.P.; Olivares, F.L.; Sangi, S.; Canellas, N.A.; Spaccini, R.; Cangemi, S.; Canellas, L.P. Humic Substances Isolated from Recycled Biomass Trigger Jasmonic Acid Biosynthesis and Signalling. Plants 2023, 12, 3148. https://doi.org/10.3390/plants12173148
Silva RM, Peres ANA, Peres LEP, Olivares FL, Sangi S, Canellas NA, Spaccini R, Cangemi S, Canellas LP. Humic Substances Isolated from Recycled Biomass Trigger Jasmonic Acid Biosynthesis and Signalling. Plants. 2023; 12(17):3148. https://doi.org/10.3390/plants12173148
Chicago/Turabian StyleSilva, Rakiely M., Alice N. A. Peres, Lázaro E. P. Peres, Fábio L. Olivares, Sara Sangi, Natália A. Canellas, Riccardo Spaccini, Silvana Cangemi, and Luciano P. Canellas. 2023. "Humic Substances Isolated from Recycled Biomass Trigger Jasmonic Acid Biosynthesis and Signalling" Plants 12, no. 17: 3148. https://doi.org/10.3390/plants12173148