

Quantifying Chilling Injury on the Photosynthesis System of Strawberries: Insights from Photosynthetic Fluorescence Characteristics and Hyperspectral Inversion


Abstract
:1. Introduction
2. Results
2.1. Impact of Continuous Dynamic Chilling Stress on Photosynthetic Parameters in Short-Day and Long-Day Strawberry Varieties
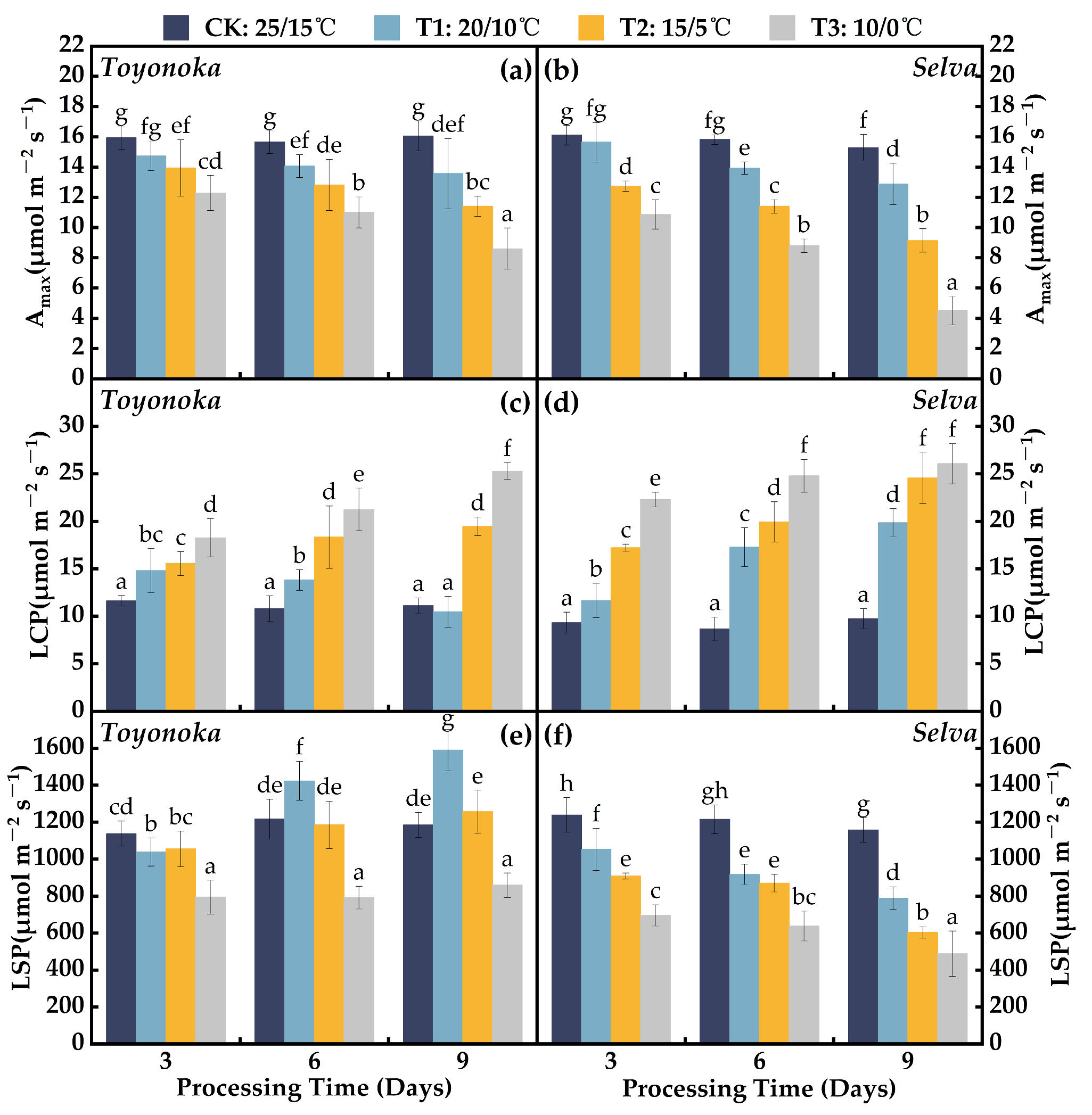
2.1.1. Light Response Parameters
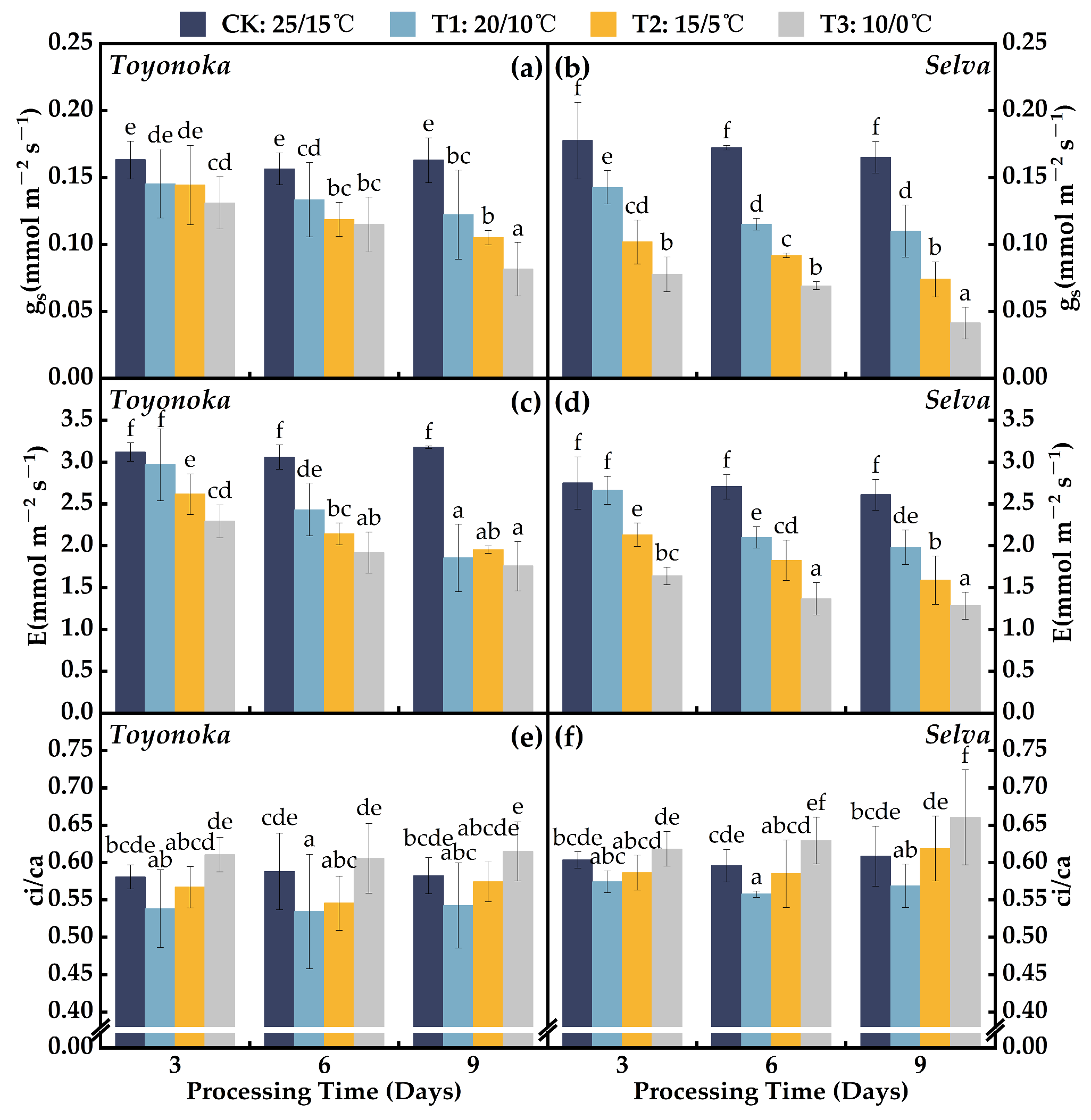
2.1.2. Gas Exchange Parameters
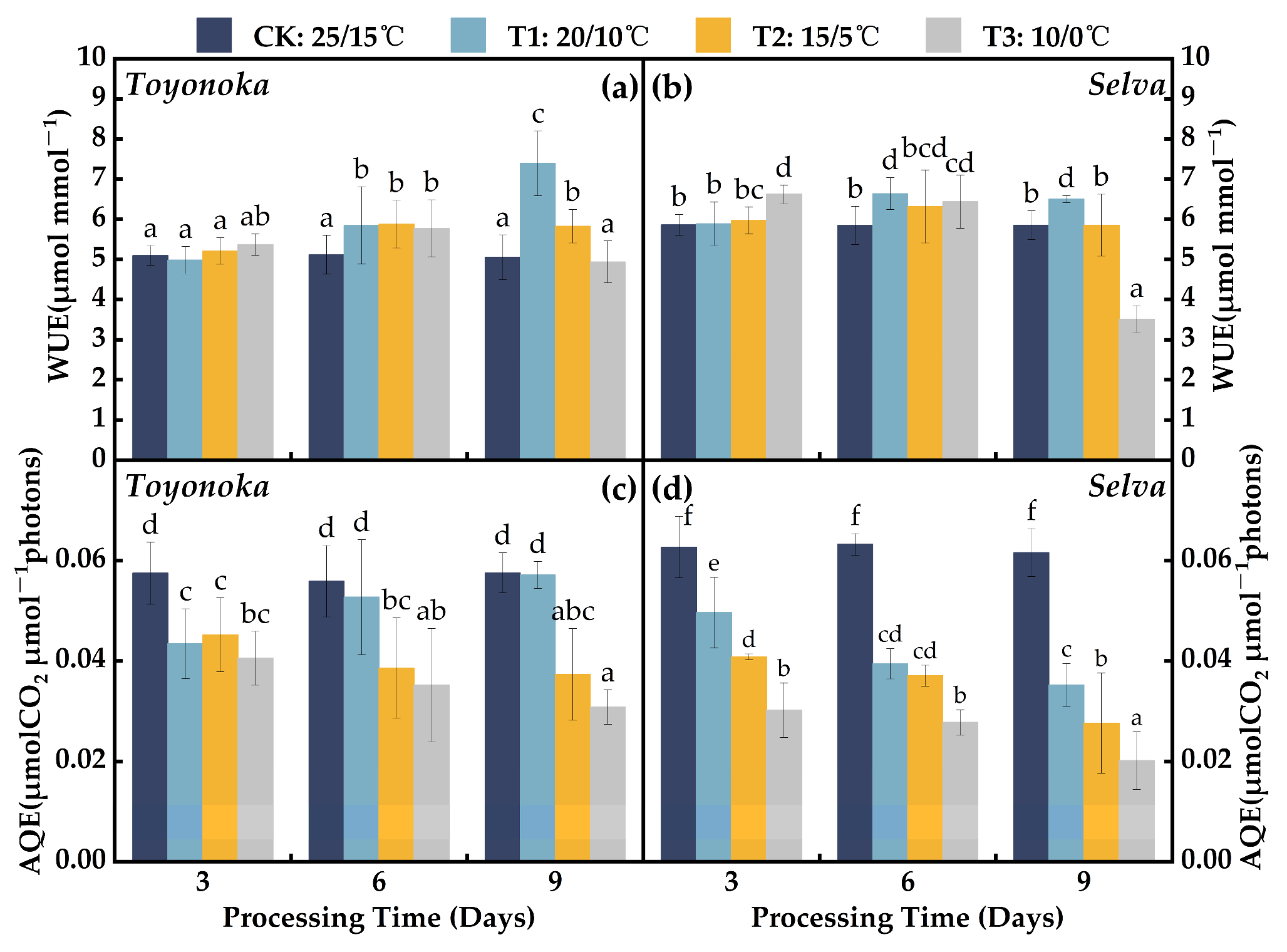
2.1.3. Parameters of Water and Light Use Efficiency
2.2. Impact of Continuous Dynamic Chilling Stress on Chlorophyll Fluorescence Induction Kinetic Parameters in Short-Day and Long-Day Strawberry Varieties
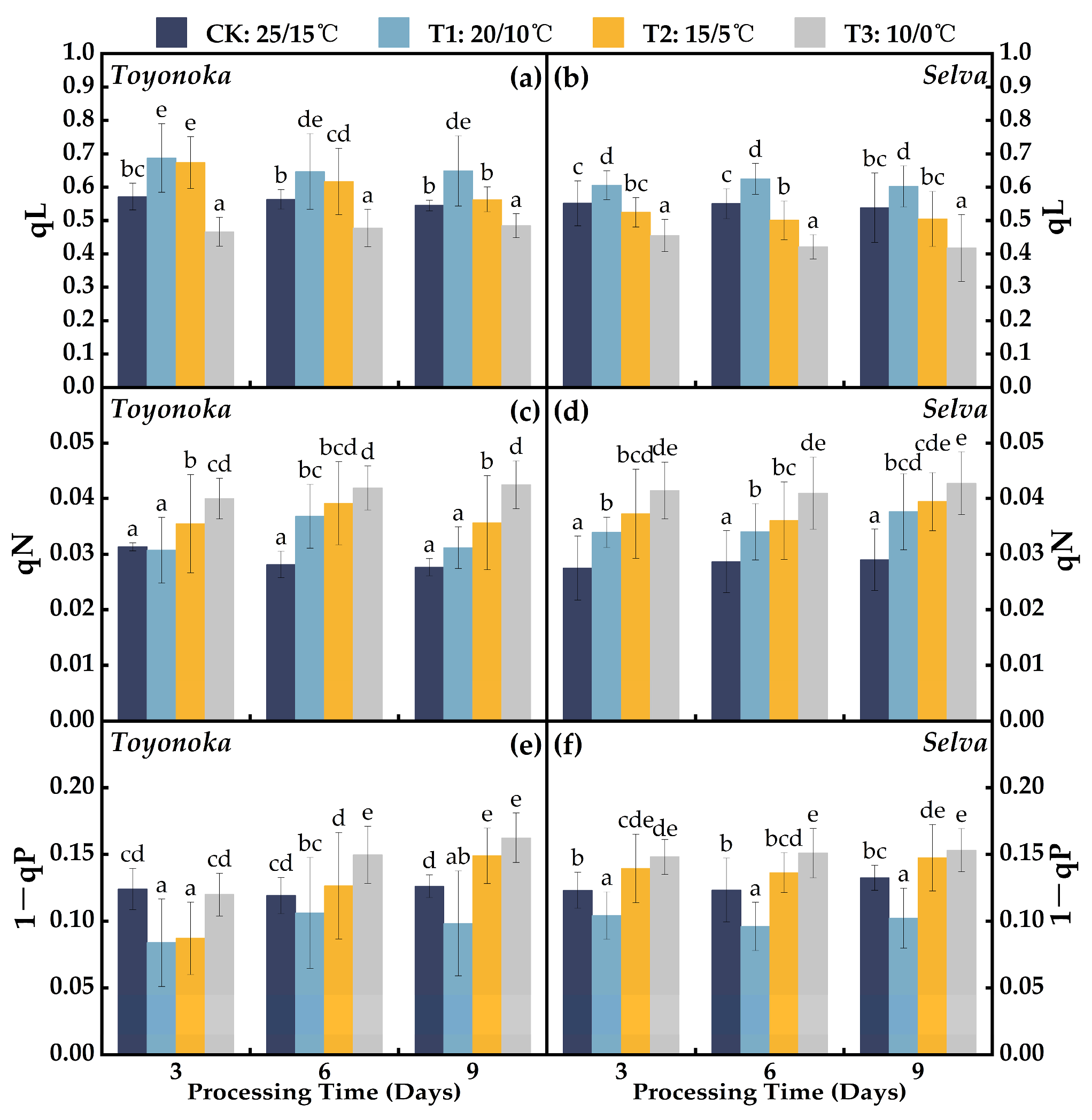
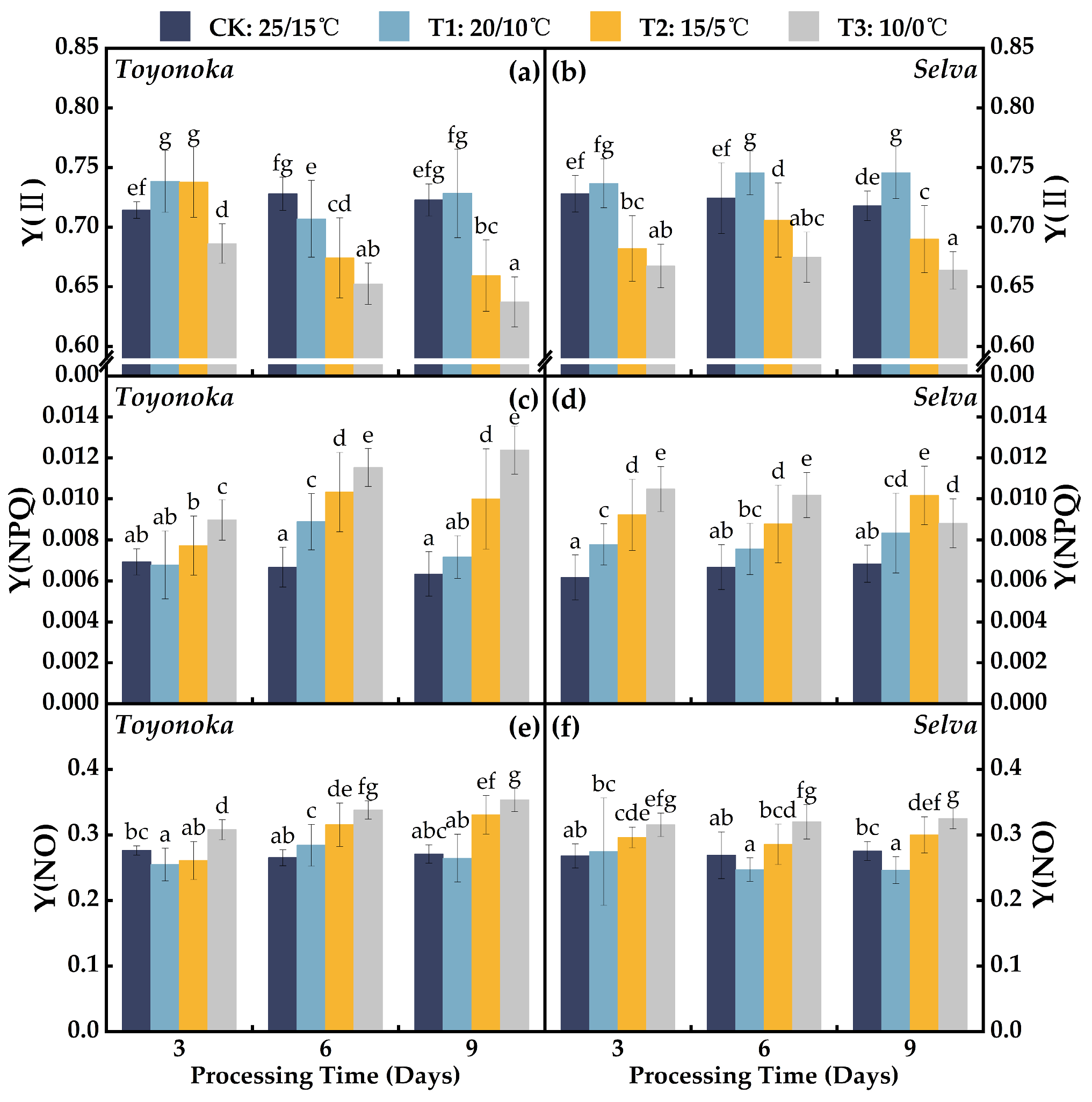
2.2.1. Photosystem II Photochemical Efficiency Parameters
2.2.2. Fluorescence Quenching Coefficients
2.2.3. Photosystem II Quantum Yield Parameters
2.3. Impact of Continuous Dynamic Chilling Stress on Solar-Induced Chlorophyll Fluorescence in Short-Day and Long-Day Strawberry Varieties
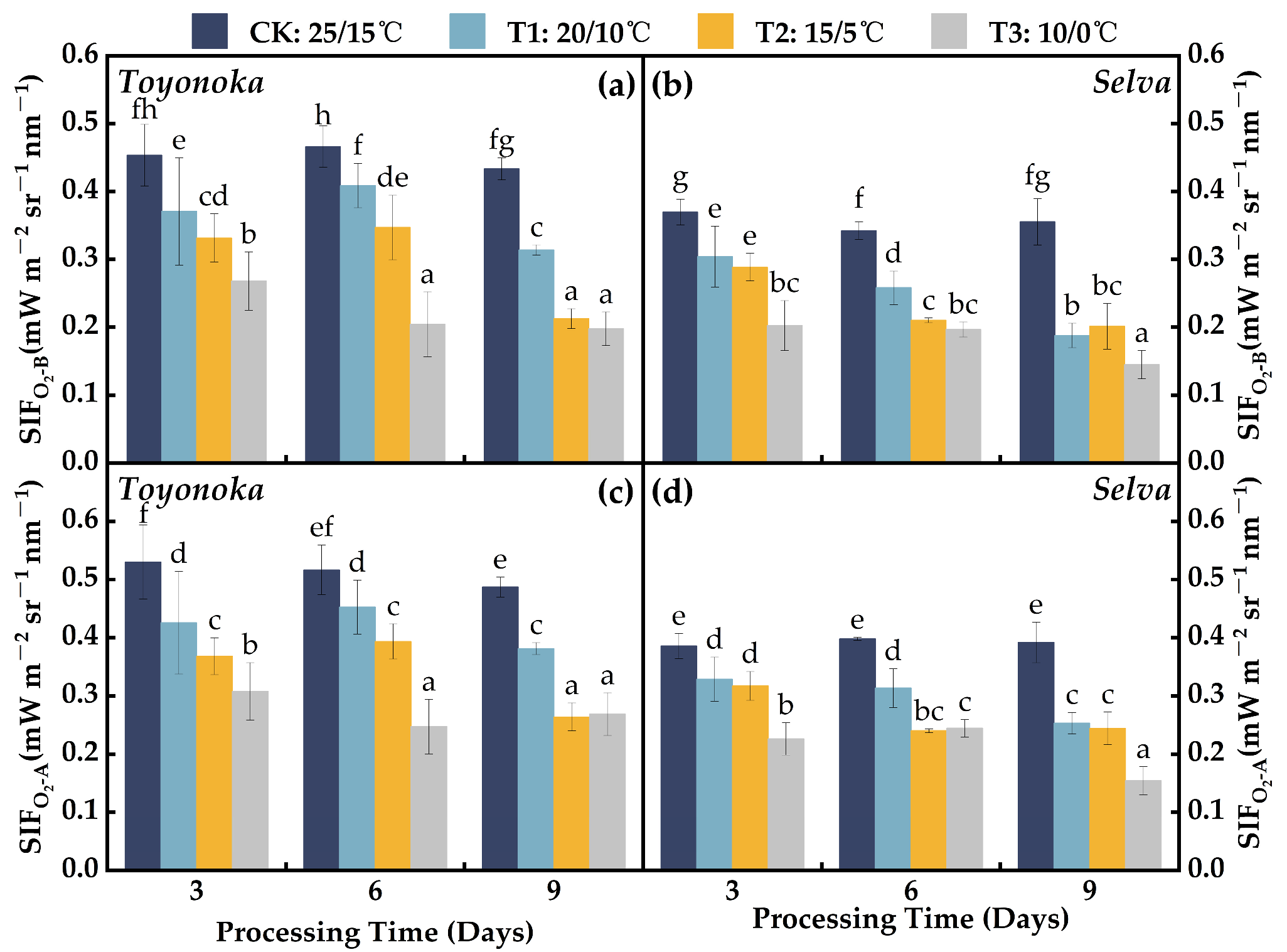
2.3.1. Solar-Induced Chlorophyll Fluorescence
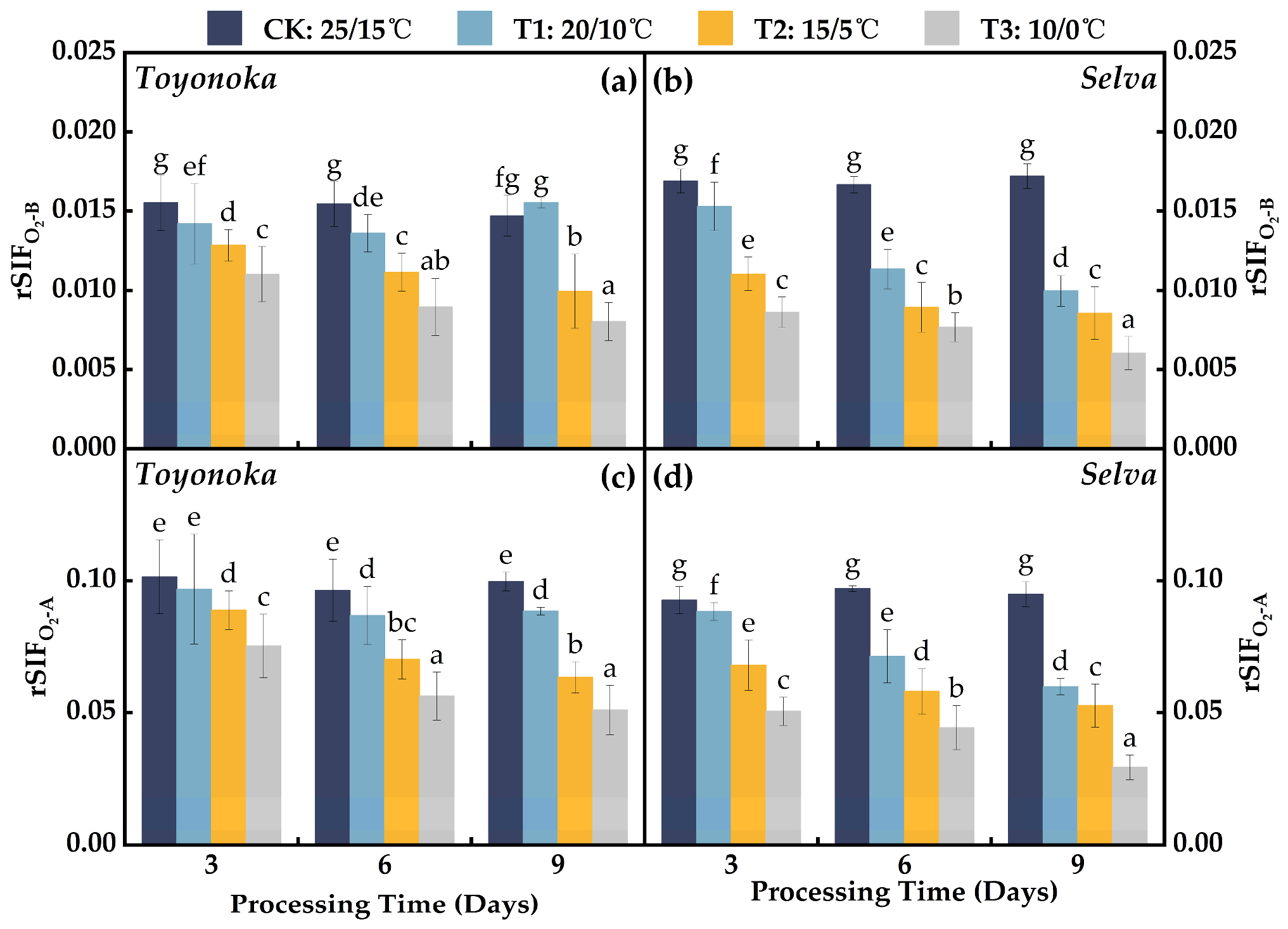
2.3.2. Relative Solar-Induced Chlorophyll Fluorescence
2.4. Construction of a Comprehensive Evaluation System for Chilling Stress on the Photosynthetic System of Strawberry with Different Photoperiod Types
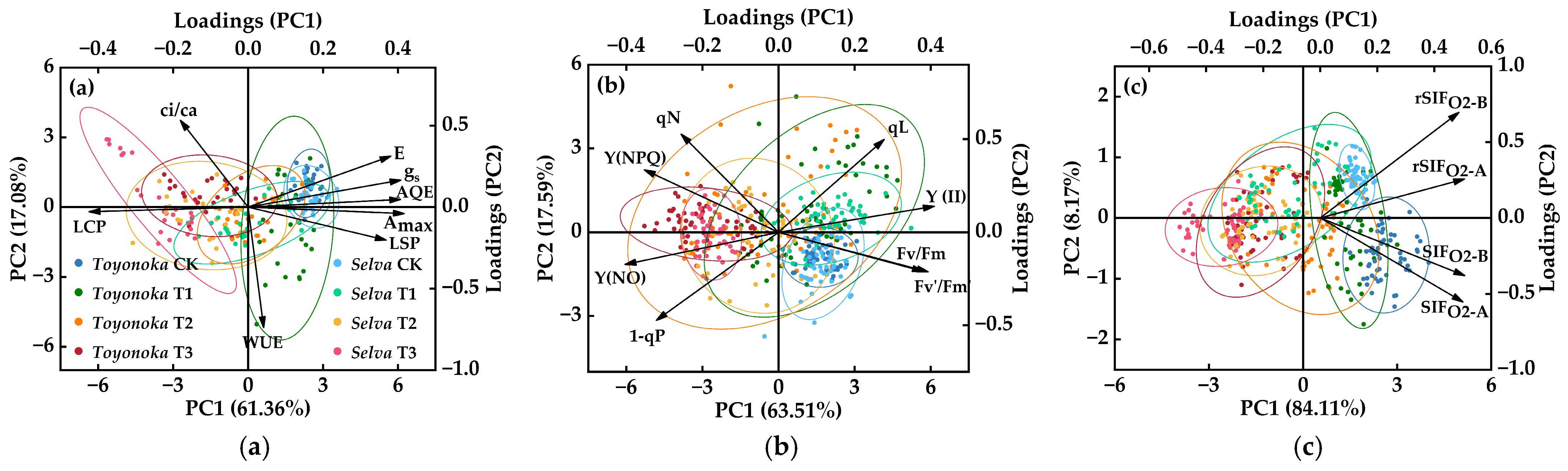
2.4.1. Extraction of Photosynthetic Physiological Characteristic Indices
2.4.2. Construction and Grading of a Comprehensive Score for Photosynthetic System Chilling Injury in Strawberries
2.5. Hyperspectral Inversion of Comprehensive Score for Photosynthetic System Chilling Injury in Strawberries with Different Photoperiod Types
2.5.1. Hyperspectral Transformation Results
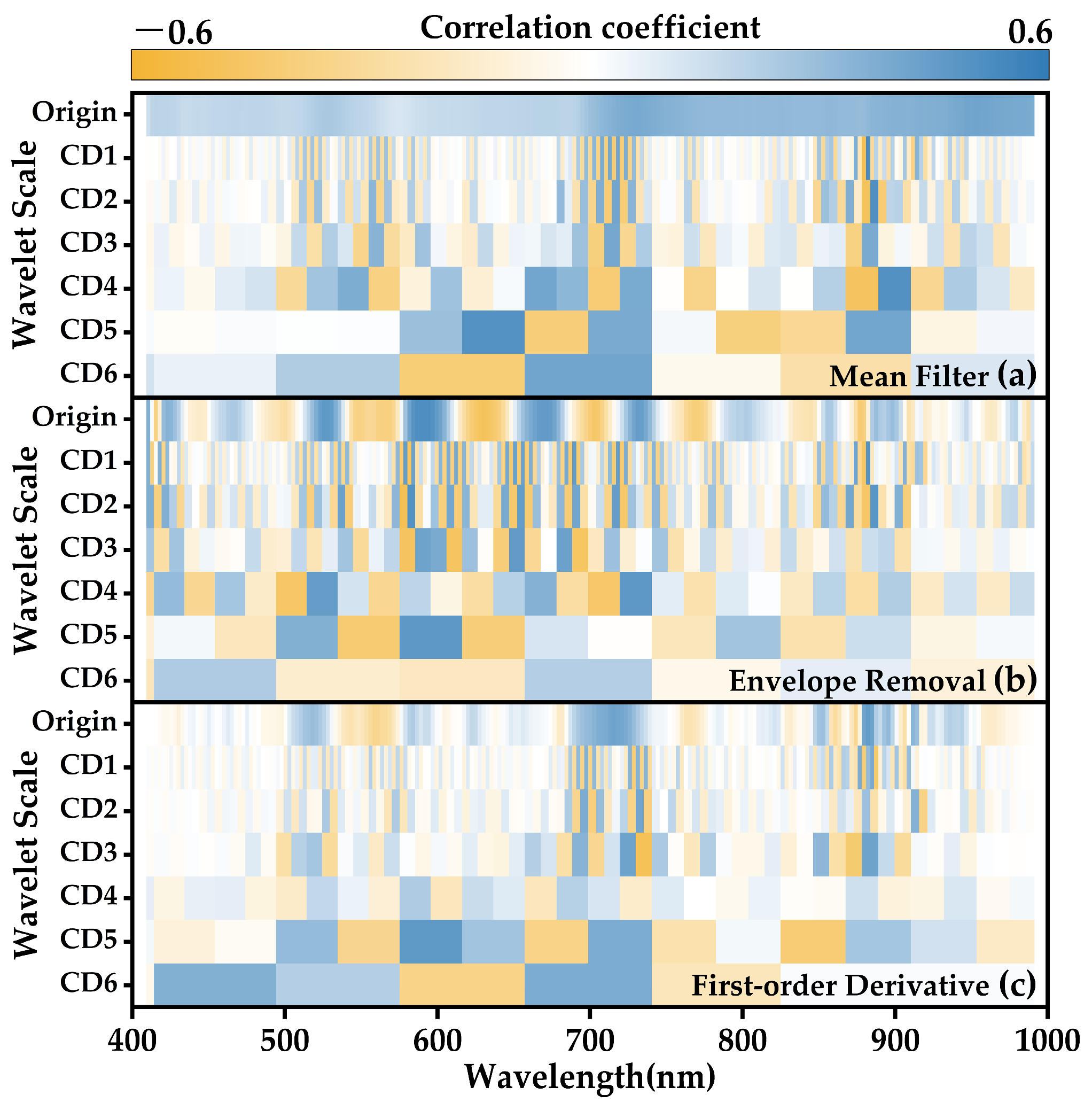
2.5.2. Extraction of Spectral Features for Photosynthetic System Chilling Injury
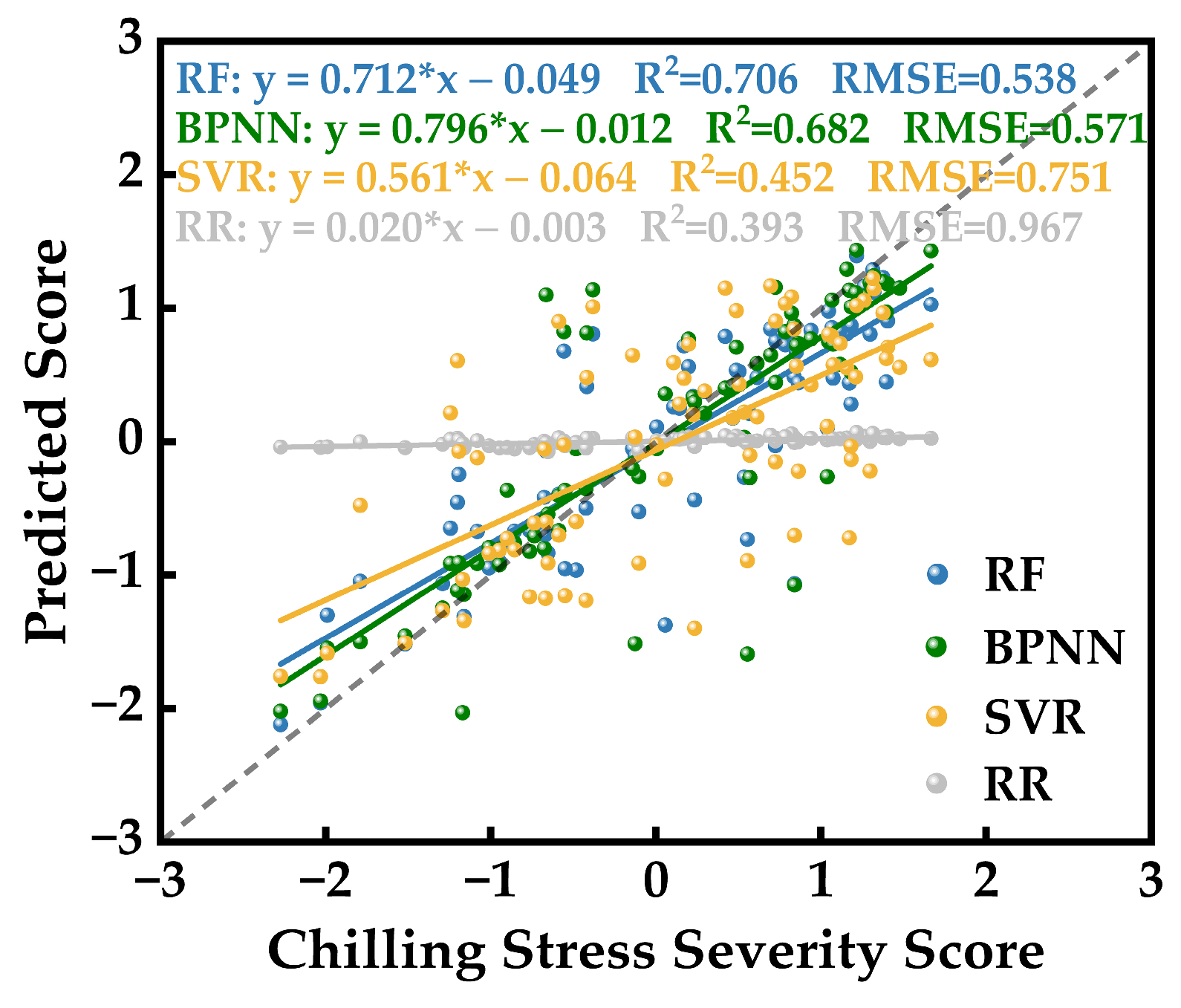
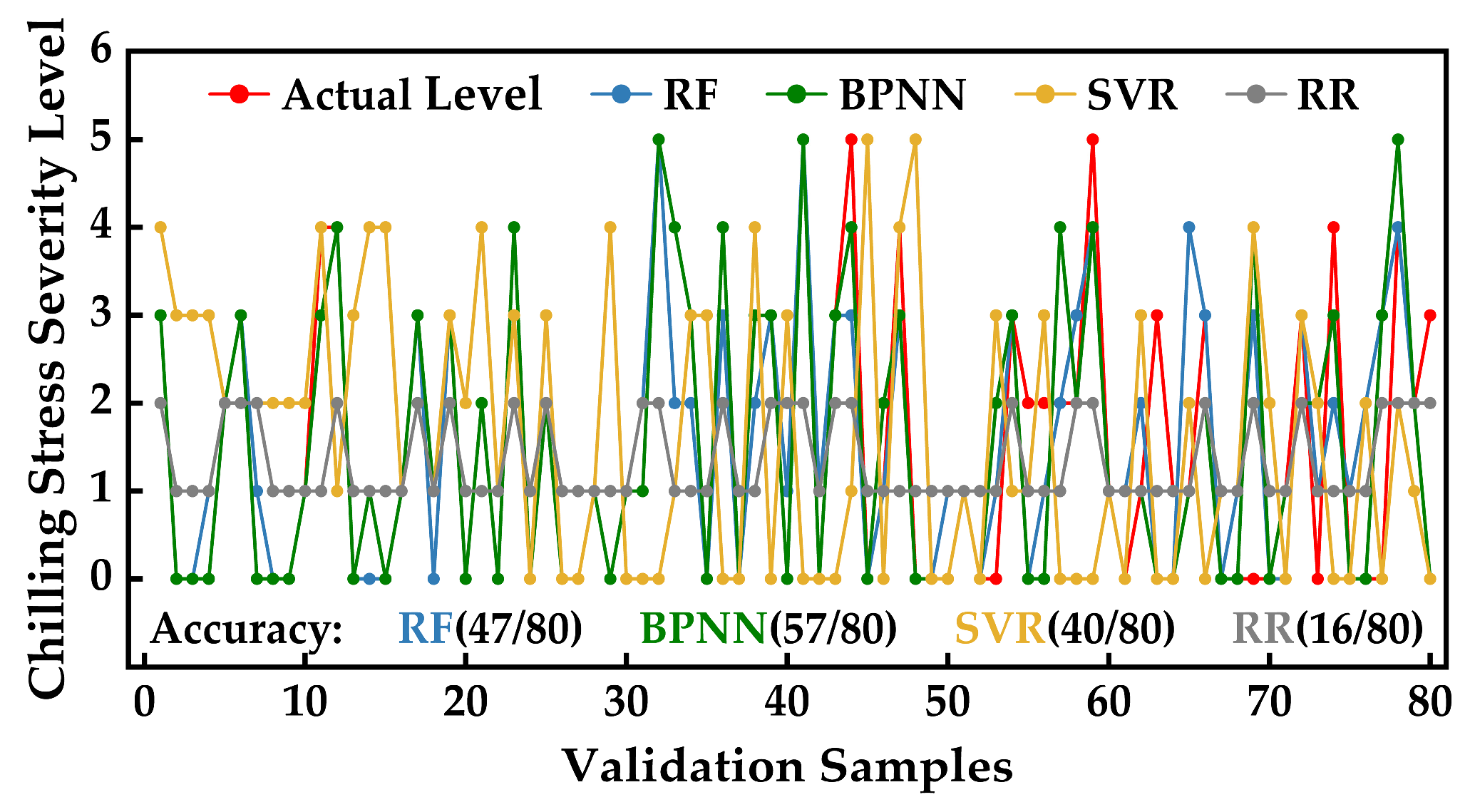
2.5.3. Hyperspectral Inversion Modeling and Accuracy Evaluation of Photosynthetic System Chilling Injury Severity in Strawberries
3. Discussion
4. Materials and Methods
4.1. Plant Materials
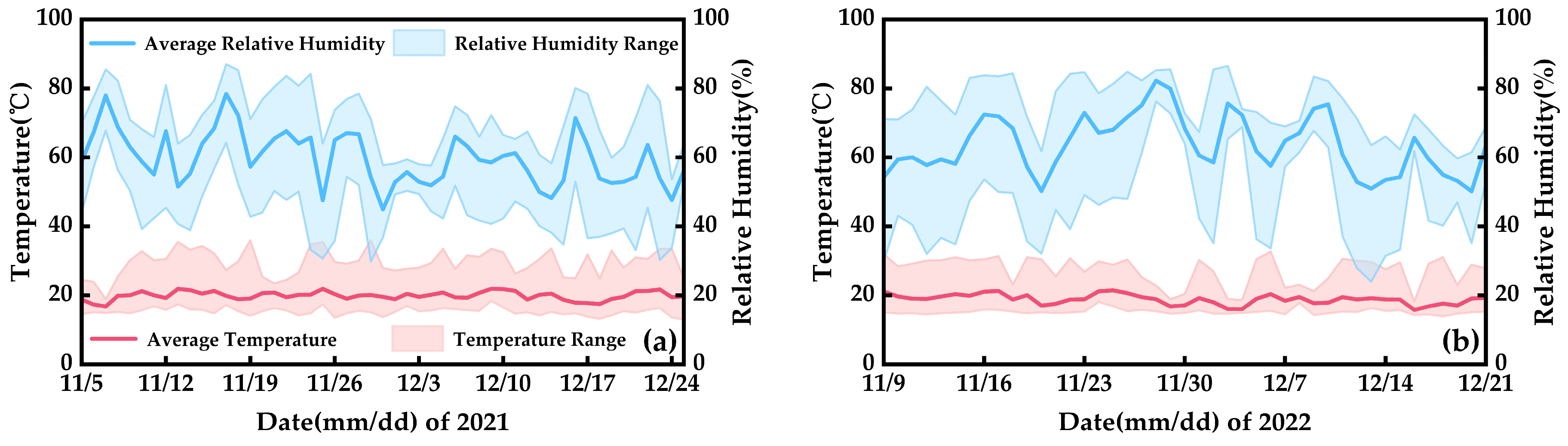
4.2. Experimental Management and Treatment
4.3. The Methods of Measurement
4.3.1. Photosynthetic Parameters
4.3.2. Chlorophyll Fluorescence Induction Kinetics Parameters
4.3.3. Solar-Induced Chlorophyll Fluorescence
4.3.4. Reflectance Spectra
4.4. Data Processing and Analysis
4.4.1. Hyperspectral Data Preprocessing
4.4.2. Statistical and Analytical Methods
4.4.3. Wavelet Transformation
4.4.4. Hyperspectral Inversion Modeling
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Basu, A.; Nguyen, A.; Betts, N.M.; Lyons, T.J. Strawberry As a Functional Food: An Evidence-Based Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 790–806. [Google Scholar] [CrossRef] [PubMed]
- Maksimović, J.D.; Poledica, M.; Mutavdžić, D.; Mojović, M.; Radivojević, D.; Milivojević, J. Variation in Nutritional Quality and Chemical Composition of Fresh Strawberry Fruit: Combined Effect of Cultivar and Storage. Plant Foods Hum. Nutr. 2015, 70, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and Human Health: Effects beyond Antioxidant Activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, D.S. Temperature and Growth Regulator Effects on Growth and Development of Strawberry (Fragaria × ananassa Duch.). 1998. Available online: https://hdl.handle.net/20.500.12123/6611 (accessed on 3 June 2021).
- Atwell, B.J. Plants in Action: Adaptation in Nature, Performance in Cultivation; Macmillan Education AU: South Yarra, Australia, 1999. [Google Scholar]
- Simpson, D. The Economic Importance of Strawberry Crops. In The Genomes of Rosaceous Berries and Their Wild Relatives; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–7. [Google Scholar] [CrossRef]
- Hoffmann, R.; Muttarak, R.; Peisker, J.; Stanig, P. Climate change experiences raise environmental concerns and promote Green voting. Nat. Clim. Change 2022, 12, 148–155. [Google Scholar] [CrossRef]
- Ariza, M.T.; Soria, C.; Martínez-Ferri, E. Developmental stages of cultivated strawberry flowers in relation to chilling sensitivity. AoB Plants 2015, 7, plv012. [Google Scholar] [CrossRef]
- Khammayom, N.; Maruyama, N.; Chaichana, C. The Effect of Climatic Parameters on Strawberry Production in a Small Walk-In Greenhouse. Agriengineering 2022, 4, 104–121. [Google Scholar] [CrossRef]
- Stephenson, A.G. Flower and Fruit Abortion: Proximate Causes and Ultimate Functions. Annu. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Kadir, S.; Sidhu, G.; Al-Khatib, K. Strawberry (Fragaria × ananassa Duch.) Growth and Productivity as Affected by Temperature. HortScience 2006, 41, 1423–1430. [Google Scholar] [CrossRef]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Physiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- He, X.; Hao, J.; Fan, S.; Liu, C.; Han, Y. Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress. Plants 2022, 11, 1385. [Google Scholar] [CrossRef]
- Meng, A.; Wen, D.; Zhang, C. Maize Seed Germination under Low-Temperature Stress Impacts Seedling Growth Under Normal Temperature by Modulating Photosynthesis and Antioxidant Metabolism. Front. Plant Sci. 2022, 13, 843033. [Google Scholar] [CrossRef]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Linking leaf chlorophyll fluorescence properties to physiological responses for detection of salt and drought stress in coastal plant species. Physiol. Plant. 2007, 131, 422–433. [Google Scholar] [CrossRef]
- Massacci, A.; Nabiev, S.; Pietrosanti, L.; Nematov, S.; Chernikova, T.; Thor, K.; Leipner, J. Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2008, 46, 189–195. [Google Scholar] [CrossRef]
- Magney, T.S.; Bowling, D.R.; Logan, B.; Grossmann, K.; Stutz, J.; Blanken, P.D.; Burns, S.P.; Cheng, R.; Garcia, M.A.; Köhler, P.; et al. Mechanistic evidence for tracking the seasonality of photosynthesis with solar-induced fluorescence. Proc. Natl. Acad. Sci. USA 2019, 116, 11640–11645. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.; Celesti, M.; Acebron, K.; Alberti, G.; Cogliati, S.; Colombo, R.; Juszczak, R.; Matsubara, S.; Miglietta, F.; Palombo, A.; et al. Dynamics of sun-induced chlorophyll fluorescence and reflectance to detect stress-induced variations in canopy photosynthesis. Plant Cell Environ. 2020, 43, 1637–1654. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence and Photosynthesis: The Basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Mertens, S.; Verbraeken, L.; Sprenger, H.; Demuynck, K.; Maleux, K.; Cannoot, B.; De Block, J.; Maere, S.; Nelissen, H.; Bonaventure, G.; et al. Proximal Hyperspectral Imaging Detects Diurnal and Drought-Induced Changes in Maize Physiology. Front. Plant Sci. 2021, 12, 640914. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Alchanatis, V.; Yang, C.; Hirafuji, M.; Moshou, D.; Li, C. Sensing technologies for precision specialty crop production. Comput. Electron. Agric. 2010, 74, 2–33. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Masoni, A.; Ercoli, L.; Mariotti, M. Spectral Properties of Leaves Deficient in Iron, Sulfur, Magnesium, and Manganese. Agron. J. 1996, 88, 937–943. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Gu, Y.; Wylie, B.K.; Howard, D.M.; Phuyal, K.P.; Ji, L. NDVI saturation adjustment: A new approach for improving cropland performance estimates in the Greater Platte River Basin, USA. Ecol. Indic. 2013, 30, 1–6. [Google Scholar] [CrossRef]
- Endo, T.; Okuda, T.; Tamura, M.; Yasuoka, Y. Estimation of net photosynthetic rate based on in-situ hyperspectral data. In Hyperspectral Remote Sensing of the Land and Atmosphere; SPIE: Bellingham, WA, USA, 2001; Volume 4151, pp. 214–221. [Google Scholar] [CrossRef]
- Yang, Z.; Tian, J.; Wang, Z.; Feng, K. Monitoring the photosynthetic performance of grape leaves using a hyperspectral-based machine learning model. Eur. J. Agron. 2022, 140, 126589. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L.; Sampson, P.H. Estimation of chlorophyll fluorescence under natural illumination from hyperspectral data. Int. J. Appl. Earth Obs. Geoinf. 2001, 3, 321–327. [Google Scholar] [CrossRef]
- Jiang, N.; Yang, Z.; Zhang, H.; Xu, J.; Li, C. Effect of Low Temperature on Photosynthetic Physiological Activity of Different Photoperiod Types of Strawberry Seedlings and Stress Diagnosis. Agronomy 2023, 13, 1321. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, C.; Niu, Y.; Shen, L.; Wang, W. The severity of heat and cold waves amplified by high relative humidity in humid subtropical basins: A case study in the Gan River Basin, China. Nat. Hazards 2023, 115, 865–898. [Google Scholar] [CrossRef]
- Kurczyn, J.A.; Appendini, C.M.; Beier, E.; Sosa-López, A.; López-González, J.; Posada-Vanegas, G. Oceanic and atmospheric impact of central American cold surges (Nortes) in the Gulf of Mexico. Int. J. Clim. 2021, 41, E1450–E1468. [Google Scholar] [CrossRef]
- Bilger, W.; Schreiber, U. Energy-dependent quenching of dark-level chlorophyll fluorescence in intact leaves. Photosyn. Res. 1986, 10, 303–308. [Google Scholar] [CrossRef]
- Ryu, Y.; Jiang, C.; Kobayashi, H.; Detto, M. MODIS-derived global land products of shortwave radiation and diffuse and total photosynthetically active radiation at 5 km resolution from 2000. Remote Sens. Environ. 2018, 204, 812–825. [Google Scholar] [CrossRef]
- Bouwer, L.M. Observed and Projected Impacts from Extreme Weather Events: Implications for Loss and Damage. In Loss and Damage from Climate Change: Concepts, Methods and Policy Options; Springer Nature: Berlin/Heidelberg, Germany, 2019; pp. 63–82. [Google Scholar] [CrossRef]
- Li, H.; Li, T.; Gordon, R.J.; Asiedu, S.K.; Hu, K. Strawberry plant fruiting efficiency and its correlation with solar irradiance, temperature and reflectance water index variation. Environ. Exp. Bot. 2010, 68, 165–174. [Google Scholar] [CrossRef]
- Ebadi, A.; Coombe, B.; May, P. Fruit-set on small Chardonnay and Shiraz vines grown under varying temperature regimes between budburst and flowering. Aust. J. Grape Wine Res. 1995, 1, 3–10. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Eisenhut, M.; Bräutigam, A.; Timm, S.; Florian, A.; Tohge, T.; Fernie, A.R.; Bauwe, H.; Weber, A.P. Photorespiration Is Crucial for Dynamic Response of Photosynthetic Metabolism and Stomatal Movement to Altered CO2 Availability. Mol. Plant 2017, 10, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Kržič, N.; Gaberščik, A. Photochemical efficiency of amphibious plants in an intermittent lake. Aquat. Bot. 2005, 83, 281–288. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 201–247. Available online: https://www.walz.com/files/downloads/pan/PAN078007.pdf (accessed on 17 July 2021).
- Yang, X.; Tang, J.; Mustard, J.F.; Lee, J.-E.; Rossini, M.; Joiner, J.; Munger, J.W.; Kornfeld, A.; Richardson, A.D. Solar-induced chlorophyll fluorescence that correlates with canopy photosynthesis on diurnal and seasonal scales in a temperate deciduous forest. Geophys. Res. Lett. 2015, 42, 2977–2987. [Google Scholar] [CrossRef]
- Helm, L.T.; Shi, H.; Lerdau, M.T.; Yang, X. Solar-induced chlorophyll fluorescence and short-term photosynthetic response to drought. Ecol. Appl. 2020, 30, e02101. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Frankenberg, C.; Jung, M.; Joiner, J.; Guanter, L.; Köhler, P.; Magney, T. Overview of Solar-Induced chlorophyll Fluorescence (SIF) from the Orbiting Carbon Observatory-2: Retrieval, cross-mission comparison, and global monitoring for GPP. Remote Sens. Environ. 2018, 209, 808–823. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Schimel, D.; Frankenberg, C.; Drewry, D.T.; Fisher, J.B.; Verma, M.; Berry, J.A.; Lee, J.-E.; Joiner, J. Application of satellite solar-induced chlorophyll fluorescence to understanding large-scale variations in vegetation phenology and function over northern high latitude forests. Remote Sens. Environ. 2017, 190, 178–187. [Google Scholar] [CrossRef]
- Chen, S.; Shen, Y.; Xie, P.; Lu, X.; He, Y.; Cen, H. In-situ diagnosis of citrus Huanglongbing using sun-induced chlorophyll flu-orescence. Trans. Chin. Soc. Agric. Eng. 2022, 38, 324–332. [Google Scholar] [CrossRef]
- Masarmi, A.G.; Solouki, M.; Fakheri, B.; Kalaji, H.M.; Mahgdingad, N.; Golkari, S.; Telesiński, A.; Lamlom, S.F.; Kociel, H.; Yousef, A.F. Comparing the salinity tolerance of twenty different wheat genotypes on the basis of their physiological and biochemical parameters under NaCl stress. PLoS ONE 2023, 18, e0282606. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef]
- Prananto, J.A.; Minasny, B.; Weaver, T. Near infrared (NIR) spectroscopy as a rapid and cost-effective method for nutrient analysis of plant leaf tissues. Adv. Agron. 2020, 164, 1–49. [Google Scholar] [CrossRef]
- Wei, T.; Yang, Z.; Wang, L.; Zhao, H.; Li, J. Simulation Model of Hourly Air Temperature inside Glass Greenhouse and Plastic Greenhouse. Chin. J. Agrometeorol. 2018, 39, 644–655. Available online: https://10.3969/j.issn.1000-6362.2018.10.003 (accessed on 13 April 2021).
- Evans, J.R.; Santiago, L.S. PrometheusWiki Gold Leaf Protocol: Gas exchange using LI-COR 6400. Funct. Plant Biol. 2014, 41, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Downton, W.J.S. Environmental regulation of photosynthesis. Photosynthesis 1982, 2, 263–343. Available online: http://hdl.handle.net/102.100.100/287736?index=1 (accessed on 17 March 2021).
- Ye, Z.; Suggett, D.J.; Robakowski, P.; Kang, H. A mechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef]
- Ye, Z. A review on modeling of responses of photosynthesis to light and CO2. J. Plant Ecol. 2010, 34, 727–740. (In Chinese) [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Springer: Dordrecht, The Netherlands, 2004; pp. 279–319. [Google Scholar] [CrossRef]
- Maier, S.W.; Günther, K.P.; Stellmes, M. Sun-Induced Fluorescence: A New Tool for Precision Farming. In Digital Imaging and Spectral Techniques: Applications to Precision Agriculture and Crop Physiology; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2004; Volume 66, pp. 207–222. [Google Scholar] [CrossRef]
- Pitas, I.; Venetsanopoulos, A. Nonlinear mean filters in image processing. IEEE Trans. Acoust. Speech Signal Process. 1986, 34, 573–584. [Google Scholar] [CrossRef]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Z.; Zhang, F.; Li, C. Effect of nitrogen application on enhancing high-temperature stress tolerance of tomato plants during the flowering and fruiting stage. Front. Plant Sci. 2023, 14, 1172078. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yuan, L.; Pu, R.; Loraamm, R.W.; Yang, G.; Wang, J. Comparison between wavelet spectral features and conventional spectral features in detecting yellow rust for winter wheat. Comput. Electron. Agric. 2014, 100, 79–87. [Google Scholar] [CrossRef]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Burrus, C.S. Wavelets and Wavelet Transforms. 2015. Available online: https://scholarship.rice.edu/bitstream/handle/1911/112342/col11454-FINAL.pdf?sequence=1 (accessed on 6 May 2023).
- Goh, A. Back-propagation neural networks for modeling complex systems. Artif. Intell. Eng. 1995, 9, 143–151. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Smola, A.J.; Schölkopf, B. A tutorial on support vector regression. Stat. Comput. 2004, 14, 199–222. [Google Scholar] [CrossRef]
- Hoerl, A.E.; Kennard, R.W. Ridge Regression: Applications to Nonorthogonal Problems. Technometrics 1970, 12, 69–82. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principal Components | PC1 | PC2 | PC3 | |
|---|---|---|---|---|
| Eigenvalue | 3.43 | 1.072 | 0.621 | |
| Variance contribution (%) | 57.167 | 17.874 | 10.356 | |
| Cumulative variance contribution (%) | 57.167 | 75.041 | 85.397 | |
| Loadings | 0.047 | 0.926 | −0.278 | |
| −0.506 | 0.012 | 0.043 | ||
| 0.359 | 0.281 | 0.800 | ||
| −0.396 | 0.047 | 0.526 | ||
| 0.456 | −0.247 | −0.024 | ||
| 0.498 | −0.013 | −0.067 | ||
| Variables | Formulas |
|---|---|
| PC1 Score | |
| PC2 Score | |
| PC3 Score | |
| Composite Score |
| Comprehensive Score for Photosynthetic System Chilling Injury | Strawberry Chilling Stress Severity Level |
|---|---|
| 0.54 ≤ CSPC | Level 0 |
| 0 ≤ CSPC < 0.54 | Level 1 |
| −0.54 ≤ CSPC < 0 | Level 2 |
| −1.08 ≤ CSPC < −0.54 | Level 3 |
| −1.62 ≤ CSPC < −1.08 | Level 4 |
| CSPC < −1.62 | Level 5 |
| Strawberry Chilling Injury Severity | |||||||
|---|---|---|---|---|---|---|---|
| Varieties | Days of Duration | 3 d | 6 d | 9 d | |||
| Daily Maximum Temperature/Daily Minimum Temperature | Score | Level | Score | Level | Score | Level | |
| Toyonoka | 25 °C/15 °C | 0.947 | 0 | 1.202 | 0 | 1.009 | 0 |
| 20 °C/10 °C | 0.704 | 0 | −0.250 | 1 | 1.234 | 0 | |
| 15 °C/5 °C | 0.403 | 1 | −0.339 | 2 | −0.842 | 3 | |
| 10 °C/0 °C | −0.525 | 2 | −1.231 | 4 | −1.813 | 5 | |
| Selva | 25 °C/15 °C | 1.323 | 0 | 1.218 | 0 | 1.172 | 0 |
| 20 °C/10 °C | 0.876 | 0 | 0.397 | 1 | −0.094 | 2 | |
| 15 °C/5 °C | −0.294 | 2 | −0.470 | 3 | −1.045 | 4 | |
| 10 °C/0 °C | −1.024 | 3 | −1.593 | 4 | −2.157 | 5 | |
| Characteristic Spectral Information | Mean Filter | Envelope Removal | First-Order Derivative | ||||
|---|---|---|---|---|---|---|---|
| Wavelength | Correlation Coefficient | Wavelength | Correlation Coefficient | Wavelength | Correlation Coefficient | ||
| Origin | Ba | 954.82 | 0.417 | 589.22 | 0.516 | 882.18 | 0.492 |
| Bb | 573.91 | 0.115 | 630.25 | −0.480 | 558.64 | −0.333 | |
| Ba − Bb | 0.440 | 0.521 | 0.472 | ||||
| Ba/Bb | 0.264 | −0.314 | −0.083 | ||||
| (Ba − Bb)/(Ba + Bb) | 0.273 | 0.052 | 0.060 | ||||
| CD1 | Ba | 882.18 | 0.534 | 882.18 | 0.495 | 884.86 | 0.391 |
| Bb | 879.51 | −0.508 | 581.56 | −0.468 | 887.54 | −0.417 | |
| Ba − Bb | 0.527 | 0.532 | 0.409 | ||||
| Ba/Bb | −0.030 | −0.053 | −0.016 | ||||
| (Ba − Bb)/(Ba + Bb) | 0.032 | −0.059 | −0.071 | ||||
| CD2 | Ba | 884.86–887.54 | 0.510 | 581.56–584.12 | 0.492 | 731.40–734.02 | 0.411 |
| Bb | 879.51–882.18 | −0.481 | 576.46–579.01 | −0.498 | 700.11–702.71 | −0.376 | |
| Ba − Bb | 0.499 | 0.497 | 0.436 | ||||
| Ba/Bb | −0.069 | 0.010 | −0.187 | ||||
| (Ba − Bb)/(Ba + Bb) | 0.108 | 0.051 | −0.009 | ||||
| CD3 | Ba | 710.52–718.34 | 0.398 | 648.29–656.03 | 0.451 | 720.95–728.79 | 0.424 |
| Bb | 700.11–707.92 | −0.378 | 576.46–584.12 | −0.474 | 731.40–739.25 | −0.485 | |
| Ba − Bb | 0.391 | 0.485 | 0.462 | ||||
| Ba/Bb | 0.277 | 0.046 | −0.213 | ||||
| (Ba − Bb)/(Ba + Bb) | −0.011 | 0.040 | −0.092 | ||||
| CD4 | Ba | 890.21–908.99 | 0.504 | 720.95–739.25 | 0.469 | 576.46–594.34 | 0.237 |
| Bb | 868.82–887.53 | −0.473 | 495.40–513.04 | −0.445 | 596.90–614.83 | −0.187 | |
| Ba − Bb | 0.497 | 0.499 | 0.217 | ||||
| Ba/Bb | 0.033 | 0.032 | 0.067 | ||||
| (Ba − Bb)/(Ba + Bb) | 0.020 | 0.076 | 0.264 | ||||
| CD5 | Ba | 617.40–656.03 | 0.498 | 576.46–614.83 | 0.471 | 576.46–614.83 | 0.461 |
| Bb | 658.62–697.50 | −0.396 | 535.79–573.91 | −0.411 | 826.23–866.15 | −0.405 | |
| Ba − Bb | 0.412 | 0.459 | 0.532 | ||||
| Ba/Bb | −0.498 | 0.023 | 0.048 | ||||
| (Ba − Bb)/(Ba + Bb) | −0.499 | −0.060 | 0.327 | ||||
| CD6 | Ba | 658.62–749.25 | 0.414 | 415.44–492.89 | 0.235 | 658.62–739.25 | 0.375 |
| Bb | 576.46–656.03 | −0.401 | 410.49–412.97 | −0.196 | 576.46–656.03 | −0.344 | |
| Ba − Bb | 0.407 | 0.232 | 0.360 | ||||
| Ba/Bb | −0.119 | 0.013 | −0.417 | ||||
| (Ba − Bb)/(Ba + Bb) | −0.015 | 0.151 | −0.368 | ||||
| Treatment Temperature (Daily Maximum Temperature/Daily Minimum Temperature) | Duration of Treatment | Marker | Batch of Treatment (Start Date–End Date) |
|---|---|---|---|
| 25 °C/15 °C (Control temperature) | 3 d | CKD3 | 14/12/2021–17/12/2021, 15/12/2021–18/12/2021, 16/12/2021–19/12/2021 10/12/2022–13/12/2022, 11/12/2022–14/12/2022, 12/12/2022–15/12/2022 |
| 6 d | CKD6 | 14/12/2021–20/12/2021, 15/12/2021–21/12/2021, 16/12/2021–22/12/2021 10/12/2022–16/12/2022, 11/12/2022–17/12/2022, 12/12/2022–18/12/2022 | |
| 9 d | CKD9 | 14/12/2021–23/12/2021, 15/12/2021–24/12/2021, 16/12/2021–25/12/2021 10/12/2022–19/12/2022, 11/12/2022–20/12/2022, 12/12/2022–21/12/2022 | |
| 20 °C/10 °C (Slightly chilling) | 3 d | T1D3 | 23/12/2021–26/12/2021, 24/12/2021–27/12/2021, 25/12/2021–28/12/2021 19/12/2022–22/12/2022, 20/12/2022–23/12/2022, 21/12/2022–24/12/2022 |
| 6 d | T1D6 | 23/12/2021–29/12/2021, 24/12/2021–30/12/2021, 25/12/2021–31/12/2021 19/12/2022–25/12/2022, 20/12/2022–26/12/2022, 21/12/2022–27/12/2022 | |
| 9 d | T1D9 | 23/12/2021–1/1/2022, 24/12/2021–2/1/2022, 25/12/2021–3/1/2022 19/12/2022–28/12/2022, 20/12/2022–29/12/2022, 21/12/2022–30/12/2022 | |
| 15 °C/5 °C (Moderate chilling) | 3 d | T2D3 | 23/12/2021–26/12/2021, 24/12/2021–27/12/2021, 25/12/2021–28/12/2021 19/12/2022–22/12/2022, 20/12/2022–23/12/2022, 21/12/2022–24/12/2022 |
| 6 d | T2D6 | 23/12/2021–29/12/2021, 24/12/2021–30/12/2021, 25/12/2021–31/12/2021 19/12/2022–25/12/2022, 20/12/2022–26/12/2022, 21/12/2022–27/12/2022 | |
| 9 d | T2D9 | 23/12/2021–1/1/2022, 24/12/2021–2/1/2022, 25/12/2021–3/1/2022 19/12/2022–28/12/2022, 20/12/2022–29/12/2022, 21/12/2022–30/12/2022 | |
| 10 °C/0 °C (Severe chilling) | 3 d | T3D3 | 14/12/2021–17/12/2021, 15/12/2021–18/12/2021, 16/12/2021–19/12/2021 10/12/2022–13/12/2022, 11/12/2022–14/12/2022, 12/12/2022–15/12/2022 |
| 6 d | T3D6 | 14/12/2021–20/12/2021, 15/12/2021–21/12/2021, 16/12/2021–22/12/2021 10/12/2022–16/12/2022, 11/12/2022–17/12/2022, 12/12/2022–18/12/2022 | |
| 9 d | T3D9 | 14/12/2021–23/12/2021, 15/12/2021–24/12/2021, 16/12/2021–25/12/2021 10/12/2022–19/12/2022, 11/12/2022–20/12/2022, 12/12/2022–21/12/2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, N.; Yang, Z.; Luo, J.; Wang, C. Quantifying Chilling Injury on the Photosynthesis System of Strawberries: Insights from Photosynthetic Fluorescence Characteristics and Hyperspectral Inversion. Plants 2023, 12, 3138. https://doi.org/10.3390/plants12173138
Jiang N, Yang Z, Luo J, Wang C. Quantifying Chilling Injury on the Photosynthesis System of Strawberries: Insights from Photosynthetic Fluorescence Characteristics and Hyperspectral Inversion. Plants. 2023; 12(17):3138. https://doi.org/10.3390/plants12173138
Chicago/Turabian StyleJiang, Nan, Zaiqiang Yang, Jing Luo, and Canyue Wang. 2023. "Quantifying Chilling Injury on the Photosynthesis System of Strawberries: Insights from Photosynthetic Fluorescence Characteristics and Hyperspectral Inversion" Plants 12, no. 17: 3138. https://doi.org/10.3390/plants12173138