
Antimicrobial, Cytotoxic, and Anti-Inflammatory Activities of Tigridia vanhouttei Extracts
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
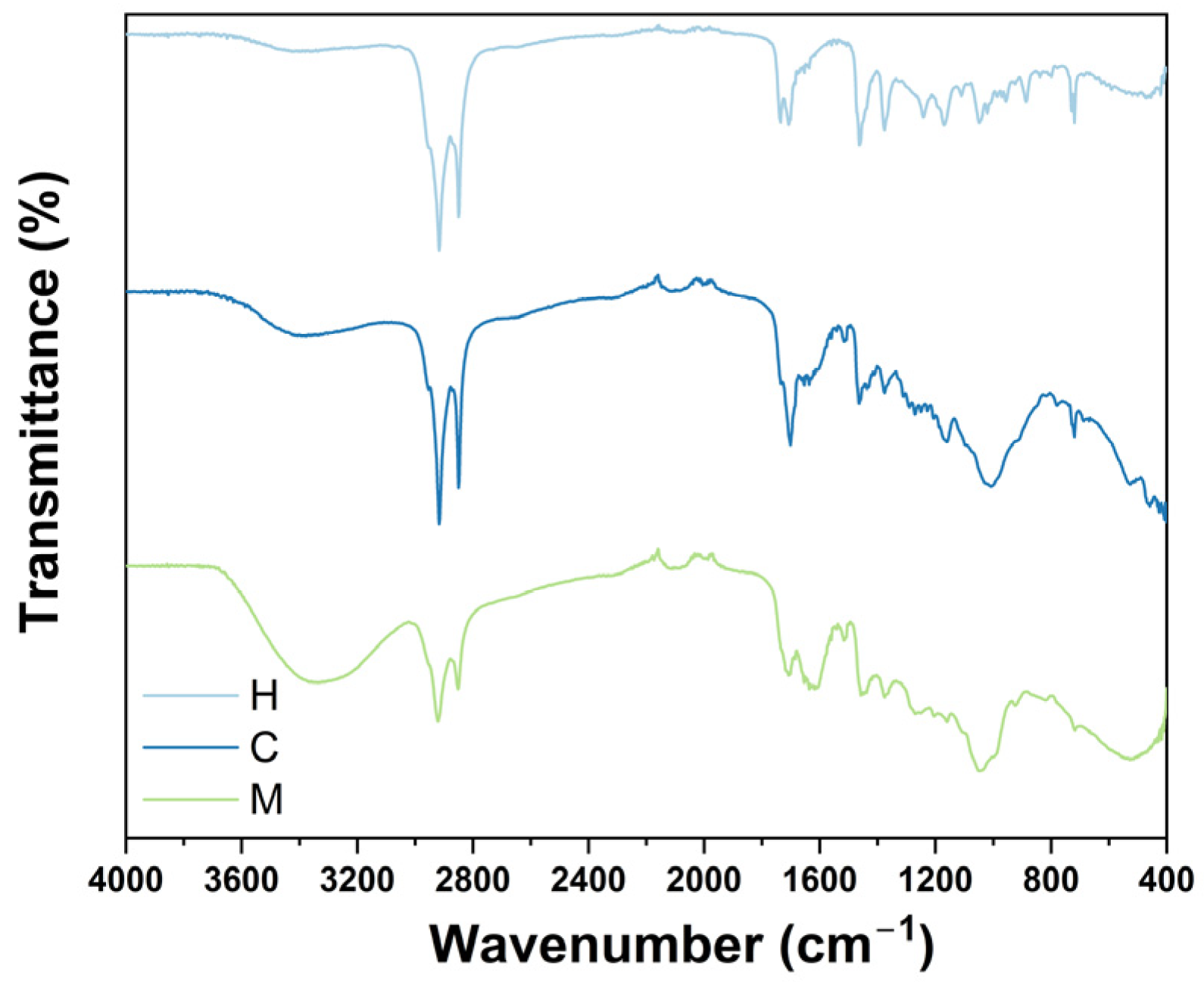
2.1. FTIR Analysis
2.2. GC-MS Analysis
2.3. Antimicrobial Activity
2.4. Cytotoxic Activity
2.5. Anti-Inflammatory Activity
3. Materials and Methods
3.1. Plant Material and Extract Preparation
3.2. FTIR Evaluation
3.3. Analysis of Phytoconstituents by GC-MS
3.4. Strains and Culture Media
3.5. Minimal Inhibitory Concentration (MIC) Assay
3.6. Cell Culture
3.7. Cytotoxicity Assay
3.8. Inflammatory Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deen, J.; Von Seidlein, L.; Clemens, J.D. Issues and Challenges of Public-Health Research in Developing Countries. Mansons Trop. Infect. Dis. 2014, 40–48.e1. [Google Scholar] [CrossRef]
- Cooper, L.A.; Purnell, T.S.; Showell, N.N.; Ibe, C.A.; Crews, D.C.; Gaskin, D.J.; Foti, K.; Thornton, R.L.J. Progress on Major Public Health Challenges: The Importance of Equity. Public Health Rep. 2018, 133, 15S–19S. [Google Scholar] [CrossRef]
- What Exactly is Antibiotic Resistance? Available online: https://www.cdc.gov/drugresistance/about.html (accessed on 1 July 2023).
- Aloke, C.; Achilonu, I. Coping with the ESKAPE pathogens: Evolving strategies, challenges and future prospects. Microb. Pathog. 2023, 175, 105963. [Google Scholar] [CrossRef]
- Arastehfar, A.; Gabaldón, T.; Garcia-Rubio, R.; Jenks, J.D.; Hoenigl, M.; Salzer, H.J.F.; Ilkit, M.; Lass-Flörl, C.; Perlin, D.S. Drug-Resistant Fungi: An Emerging Challenge Threatening Our Limited Antifungal Armamentarium. Antibiotics 2020, 9, 877. [Google Scholar] [CrossRef]
- Garvey, M.; Rowan, N.J. Pathogenic Drug Resistant Fungi: A Review of Mitigation Strategies. Int. J. Mol. Sci. 2023, 24, 1584. [Google Scholar] [CrossRef]
- Xie, X.; Li, X.; Tang, W.; Xie, P.; Tan, X. Primary tumor location in lung cancer: The evaluation and administration. Chin. Med. J. 2022, 135, 127–136. [Google Scholar] [CrossRef]
- Mejía-Méndez, J.L.; López-Mena, E.R.; Sánchez-Arreola, E. Activities against Lung Cancer of Biosynthesized Silver Nanoparticles: A Review. Biomedicines 2023, 11, 389. [Google Scholar] [CrossRef]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Saultz, J.N.; Garzon, R. Acute Myeloid Leukemia: A Concise Review. J. Clin. Med. 2016, 5, 33. [Google Scholar] [CrossRef]
- De Kouchkovsky, I.; Abdul-Hay, M. ‘Acute myeloid leukemia: A comprehensive review and 2016 update’. Blood Cancer J. 2016, 6, e441. [Google Scholar] [CrossRef]
- Nix, N.M.; Price, A. Acute Myeloid Leukemia: An Ever-Changing Disease. J. Adv. Pract. Oncol. 2019, 10, 4–8. [Google Scholar] [CrossRef]
- Nekhlyudov, L.; Campbell, G.B.; Schmitz, K.H.; Brooks, G.A.; Kumar, A.J.; Ganz, P.A.; Von Ah, D. Cancer-related impairments and functional limitations among long-term cancer survivors: Gaps and opportunities for clinical practice. Cancer 2022, 128, 222–229. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef]
- Singla, R.K.; De, R.; Efferth, T.; Mezzetti, B.; Uddin, M.S.; Sanusi; Ntie-Kang, F.; Wang, D.; Schultz, F.; Kharat, K.R.; et al. The International Natural Product Sciences Taskforce (INPST) and the power of Twitter networking exemplified through #INPST hashtag analysis. Phytomedicine 2023, 108, 154520. [Google Scholar] [CrossRef]
- Singab, A.N.B.; Ayoub, I.M.; El-Shazly, M.; Korinek, M.; Wu, T.-Y.; Cheng, Y.-B.; Chang, F.-R.; Wu, Y.-C. Shedding the light on Iridaceae: Ethnobotany, phytochemistry and biological activity. Ind. Crops Prod. 2016, 92, 308–335. [Google Scholar] [CrossRef]
- Khatib, S.; Faraloni, C.; Bouissane, L. Exploring the Use of Iris Species: Antioxidant Properties, Phytochemistry, Medicinal and Industrial Applications. Antioxidants 2022, 11, 526. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.I.M.; Hussain, F.H.S.; Najmaldin, S.K.; Thu, Z.M.; Ibrahim, M.F.; Gilardoni, G.; Vidari, G. Phytochemistry and Biological Activities of Iris Species Growing in Iraqi Kurdistan and Phenolic Constituents of the Traditional Plant Iris postii. Molecules 2021, 26, 264. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, P. Phylogeny and Classification of the Iridaceae and the Relationships of Iris. Ann. Bot. 2000, 58. [Google Scholar]
- Munguía-Lino, G.; Vargas-Ponce, O.; Rodríguez, A.; Munguía-Lino, G.; Vargas-Ponce, O.; Rodríguez, A. Tigridieae (Iridaceae) in North America: Floral diversity, flower preservation methods and keys for the identification of genera and species. Bot. Sci. 2017, 95, 473–502. [Google Scholar] [CrossRef]
- Effers, K.; Scholz, B.; Nickel, C.; Hanisch, B.; Marner, F.-J. Structure Determination of Tigridial, an Iridopentaene from Tigridia pavonia (Iridaceae). Eur. J. Org. Chem. 1999, 1999, 2793–2797. [Google Scholar] [CrossRef]
- Williams, C.A.; Harborne, J.B. Biflavonoids, Quinones and Xanthones as Rare Chemical Markers in the Family Iridaceae. Z. Für Naturforschung C 1985, 40, 325–330. [Google Scholar] [CrossRef]
- Rockwood, A.L.; Kushnir, M.M.; Clarke, N.J. 2—Mass Spectrometry. In Principles and Applications of Clinical Mass Spectrometry; Rifai, N., Horvath, A.R., Wittwer, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 33–65. ISBN 978-0-12-816063-3. [Google Scholar]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef]
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef]
- Zheng, C.J.; Yoo, J.-S.; Lee, T.-G.; Cho, H.-Y.; Kim, Y.-H.; Kim, W.-G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [PubMed]
- Bakrim, S.; Benkhaira, N.; Bourais, I.; Benali, T.; Lee, L.-H.; El Omari, N.; Sheikh, R.A.; Goh, K.W.; Ming, L.C.; Bouyahya, A. Health Benefits and Pharmacological Properties of Stigmasterol. Antioxidants 2022, 11, 1912. [Google Scholar] [CrossRef]
- McGaw, L.J.; Elgorashi, E.E.; Eloff, J.N. 8—Cytotoxicity of African Medicinal Plants Against Normal Animal and Human Cells. In Toxicological Survey of African Medicinal Plants; Kuete, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 181–233. ISBN 978-0-12-800018-2. [Google Scholar]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Pick, N.; Cameron, S.; Arad, D.; Av-Gay, Y. Screening of compounds toxicity against human Monocytic cell line-THP-1 by flow cytometry. Biol. Proced. Online 2004, 6, 220–225. [Google Scholar] [CrossRef]
- Smith, B.T. Cell line A549: A model system for the study of alveolar type II cell function. Am. Rev. Respir. Dis. 1977, 115, 285–293. [Google Scholar]
- Liu, M.; Chen, Y.-L.; Kuo, Y.-H.; Lu, M.-K.; Liao, C.-C. Aqueous extract of Sapindus mukorossi induced cell death of A549 cells and exhibited antitumor property in vivo. Sci. Rep. 2018, 8, 4831. [Google Scholar] [CrossRef]
- Olivito, F.; Amodio, N.; Di Gioia, M.L.; Nardi, M.; Oliverio, M.; Juli, G.; Tassone, P.; Procopio, A. Synthesis and preliminary evaluation of the anti-cancer activity on A549 lung cancer cells of a series of unsaturated disulfides †Electronic supplementary information (ESI) available: Experimental details and compound characterization. MedChemComm 2018, 10, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hameed, E.-S.S.; Bazaid, S.A.; Shohayeb, M.M.; El-Sayed, M.M.; El-Wakil, E.A. Phytochemical Studies and Evaluation of Antioxidant, Anticancer and Antimicrobial Properties of Conocarpus erectus L. Growing in Taif, Saudi Arabia. Eur. J. Med. Plants 2012, 2, 93–112. [Google Scholar] [CrossRef]
- Ayaz, M.; Sadiq, A.; Wadood, A.; Junaid, M.; Ullah, F.; Zaman Khan, N. Cytotoxicity and molecular docking studies on phytosterols isolated from Polygonum hydropiper L. Steroids 2019, 141, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Nath, N.; Rauf, A.; Emran, T.B.; Mitra, S.; Islam, F.; Chandran, D.; Barua, J.; Khandaker, M.U.; Idris, A.M.; et al. Multifunctional roles and pharmacological potential of β-sitosterol: Emerging evidence toward clinical applications. Chem. Biol. Interact. 2022, 365, 110117. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, J.-Q.; Sun, J.-X.; Li, H.-J. Psoralen and isopsoralen from Psoraleae Fructus aroused hepatotoxicity via induction of aryl hydrocarbon receptor-mediated CYP1A2 expression. J. Ethnopharmacol. 2022, 297, 115577. [Google Scholar] [CrossRef] [PubMed]
- Moravcová, A.; Červinková, Z.; Kučera, O.; Mezera, V.; Rychtrmoc, D.; Lotková, H. The effect of oleic and palmitic acid on induction of steatosis and cytotoxicity on rat hepatocytes in primary culture. Physiol. Res. 2015, 64, S627–S636. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Okin, D.; Medzhitov, R. Evolution of Inflammatory Diseases. Curr. Biol. CB 2012, 22, R733–R740. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Balto, K.; Sasaki, H.; Stashenko, P. Interleukin-6 Deficiency Increases Inflammatory Bone Destruction. Infect. Immun. 2001, 69, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, Y.; Ye, J. IL-6: A Potential Role in Cardiac Metabolic Homeostasis. Int. J. Mol. Sci. 2018, 19, 2474. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α Signaling in Macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of Interleukin 10 Transcriptional Regulation in Inflammation and Autoimmune Disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef]
- Rodríguez-Yoldi, M.J. Anti-Inflammatory and Antioxidant Properties of Plant Extracts. Antioxidants 2021, 10, 921. [Google Scholar] [CrossRef]
- Morgan, L.V.; Petry, F.; Scatolin, M.; de Oliveira, P.V.; Alves, B.O.; Zilli, G.A.L.; Volfe, C.R.B.; Oltramari, A.R.; de Oliveira, D.; Scapinello, J.; et al. Investigation of the anti-inflammatory effects of stigmasterol in mice: Insight into its mechanism of action. Behav. Pharmacol. 2021, 32, 640–651. [Google Scholar] [CrossRef]
- Gabay, O.; Sanchez, C.; Salvat, C.; Chevy, F.; Breton, M.; Nourissat, G.; Wolf, C.; Jacques, C.; Berenbaum, F. Stigmasterol: A phytosterol with potential anti-osteoarthritic properties. Osteoarthr. Cartil. 2010, 18, 106–116. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, F.; Shen, M.; Jia, S.; Xie, J. Phytosterols Suppress Phagocytosis and Inhibit Inflammatory Mediators via ERK Pathway on LPS-Triggered Inflammatory Responses in RAW264.7 Macrophages and the Correlation with Their Structure. Foods 2019, 8, 582. [Google Scholar] [CrossRef]
- Kasirzadeh, S.; Ghahremani, M.H.; Setayesh, N.; Jeivad, F.; Shadboorestan, A.; Taheri, A.; Beh-Pajooh, A.; Azadkhah Shalmani, A.; Ebadollahi-Natanzi, A.; Khan, A.; et al. β-Sitosterol Alters the Inflammatory Response in CLP Rat Model of Sepsis by Modulation of NFκB Signaling. BioMed Res. Int. 2021, 2021, 5535562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Durazzo, A.; Kiefer, J.; Lucarini, M.; Camilli, E.; Marconi, S.; Gabrielli, P.; Aguzzi, A.; Gambelli, L.; Lisciani, S.; Marletta, L. Qualitative Analysis of Traditional Italian Dishes: FTIR Approach. Sustainability 2018, 10, 4112. [Google Scholar] [CrossRef]
- Cuevas-Cianca, S.I.; Leal, A.C.L.; Hernández, L.R.; Arreola, E.S.; Bach, H. Antimicrobial, toxicity, and anti-inflammatory activities of Buddleja perfoliata Kunth. Phytomedicine Plus 2022, 2, 100357. [Google Scholar] [CrossRef]
- Cruz Paredes, C.; Bolívar Balbás, P.; Gómez-Velasco, A.; Juárez, Z.N.; Sánchez Arreola, E.; Hernández, L.R.; Bach, H. Antimicrobial, Antiparasitic, Anti-Inflammatory, and Cytotoxic Activities of Lopezia racemosa. Sci. World J. 2013, 2013, e237438. [Google Scholar] [CrossRef] [PubMed]
- Juárez, Z.N.; Bach, H.; Sánchez-Arreola, E.; Bach, H.; Hernández, L.R. Protective antifungal activity of essential oils extracted from Buddleja perfoliata and Pelargonium graveolens against fungi isolated from stored grains. J. Appl. Microbiol. 2016, 120, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Maycotte, P.; Acosta-Casique, A.; Rodríguez-Rodríguez, S.; Moreno, D.A.; Ferreres, F.; Flores-Alonso, J.C.; Delgado-López, M.G.; Pérez-Santos, M.; Anaya-Ruiz, M. Bursera copallifera Extracts Have Cytotoxic and Migration-Inhibitory Effects in Breast Cancer Cell Lines. Integr. Cancer Ther. 2018, 17, 654–664. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Extract | Rt (min) | R Match | Match | % | Compound |
|---|---|---|---|---|---|
| Hexane | 42.493 | 851 | 869 | 3.62 | Palmitic acid |
| 46.386 | 864 | 811 | 0.04 | Myristic acid | |
| 46.717 | 791 | 783 | 0.54 | Ascorbic acid | |
| 54.295 | 939 | 895 | 13.29 | Linolelaidic acid | |
| 54.664 | 920 | 908 | 12.27 | Oleic acid | |
| 101.676 | 875 | 814 | 1.34 | Octacosane | |
| 115.255 | 873 | 844 | 2.58 | Nonacosane | |
| 120.443 | 779 | 757 | 0.77 | Isopsoralen | |
| 122.133 | 730 | 721 | 0.53 | β-Stigmasterol | |
| 126.025 | 813 | 799 | 1.17 | ε-Sitosterol | |
| 127.875 | 866 | 811 | 2.22 | Untriacontane | |
| 133.475 | 773 | 703 | 0.42 | Sitostenone | |
| Chloroform | 67.704 | 878 | 869 | 5.26 | Hexadecanoic acid |
| 74.340 | 862 | 809 | 8.19 | Tetradecanoic acid | |
| 88.372 | 872 | 820 | 5.01 | Linolelaidic acid | |
| 90.104 | 831 | 800 | 1.73 | Oleic acid | |
| 95.842 | 790 | 764 | 6.11 | Stearic acid | |
| 129.154 | 771 | 739 | 2.16 | Arachidic acid | |
| 168.336 | 829 | 794 | 1.21 | Methyl lignocerate | |
| 180.281 | 730 | 707 | 1.11 | Campesterol | |
| 181.254 | 747 | 742 | 1.58 | Stigmasterol | |
| Methanol | 43.475 | 927 | 843 | 27.51 | Tridecanoic acid methyl ester |
| 46.164 | 827 | 794 | 33.26 | Pentadecanoic acid | |
| 55.765 | 871 | 854 | 9.93 | Linoleic acidmethyl ester | |
| 56.264 | 805 | 787 | 2.86 | Myristoleic acid | |
| 57.959 | 877 | 801 | 2.18 | Methyl tetradecanoate | |
| 58.412 | 853 | 801 | 7.35 | Stearolic acid | |
| 58.728 | 749 | 703 | 5.03 | Oleic acid | |
| 100.201 | 782 | 756 | 1.93 | β-Sitosterol |
| Extract | Bacteria | Fungi | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AB | EC | MRSA | PA | LM | SA | ABc | PAc | CA | CN | TM | |
| Hexane | R | R | R | R | R | R | R | 200 | R | R | R |
| Chloroform | R | R | R | R | R | R | R | R | R | R | R |
| Methanol | R | R | R | R | R | R | R | R | R | R | R |
| Extract | THP-1 | A549 |
|---|---|---|
| Hexane | 90.16 | 294.77 |
| Chloroform | 46.42 | 1472.37 |
| Methanol | 443.12 | 843.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mejía-Méndez, J.L.; Lorenzo-Leal, A.C.; Bach, H.; López-Mena, E.R.; Navarro-López, D.E.; Hernández, L.R.; Juárez, Z.N.; Sánchez-Arreola, E. Antimicrobial, Cytotoxic, and Anti-Inflammatory Activities of Tigridia vanhouttei Extracts. Plants 2023, 12, 3136. https://doi.org/10.3390/plants12173136
Mejía-Méndez JL, Lorenzo-Leal AC, Bach H, López-Mena ER, Navarro-López DE, Hernández LR, Juárez ZN, Sánchez-Arreola E. Antimicrobial, Cytotoxic, and Anti-Inflammatory Activities of Tigridia vanhouttei Extracts. Plants. 2023; 12(17):3136. https://doi.org/10.3390/plants12173136
Chicago/Turabian StyleMejía-Méndez, Jorge L., Ana C. Lorenzo-Leal, Horacio Bach, Edgar R. López-Mena, Diego E. Navarro-López, Luis R. Hernández, Zaida N. Juárez, and Eugenio Sánchez-Arreola. 2023. "Antimicrobial, Cytotoxic, and Anti-Inflammatory Activities of Tigridia vanhouttei Extracts" Plants 12, no. 17: 3136. https://doi.org/10.3390/plants12173136