


Enhancing Phosphorus and Nitrogen Uptake in Maize Crops with Food Industry Biosolids and Azotobacter nigricans
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Properties of Soil and Biosolids
2.2. Activity of Azotobacter Nigricans
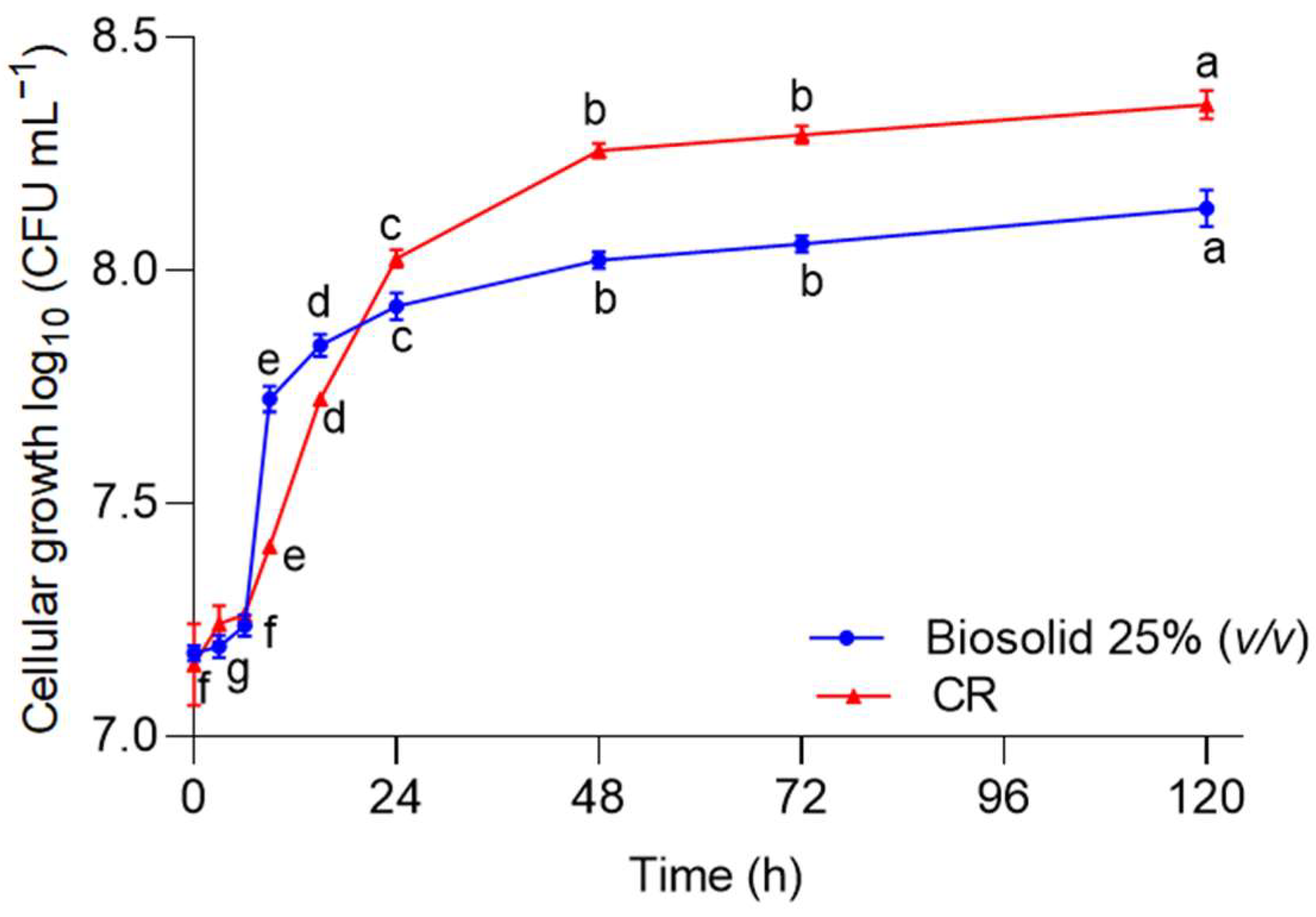
2.2.1. Inoculation Time Determination
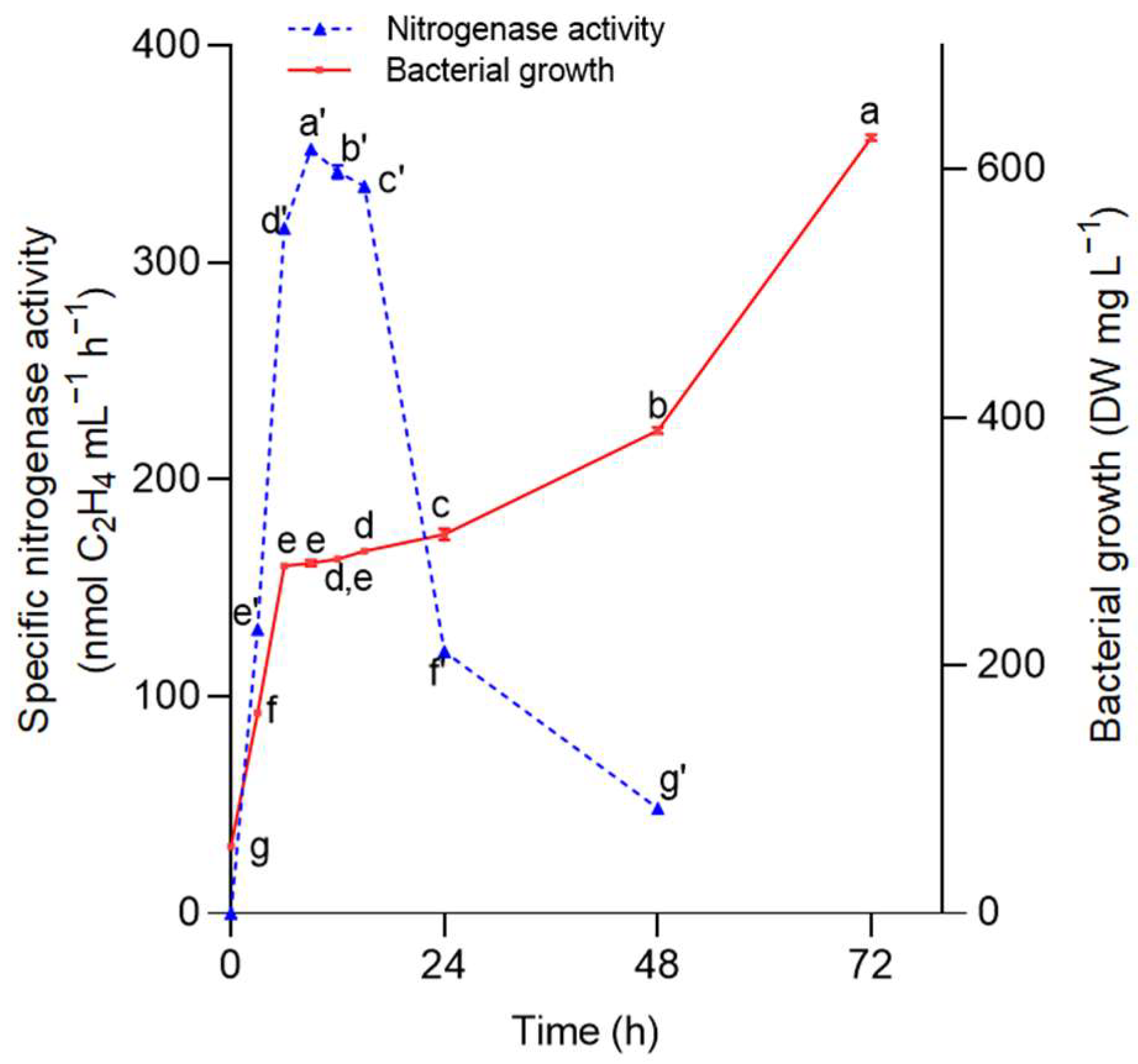
2.2.2. Nitrogenase Activity
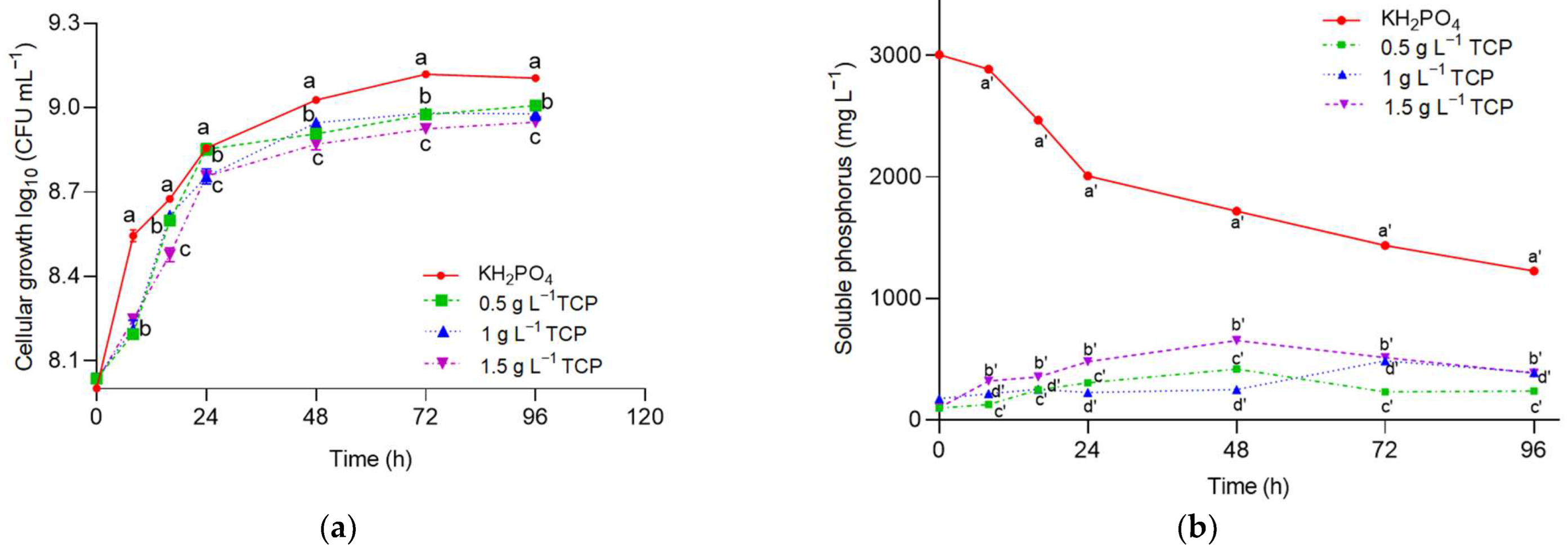
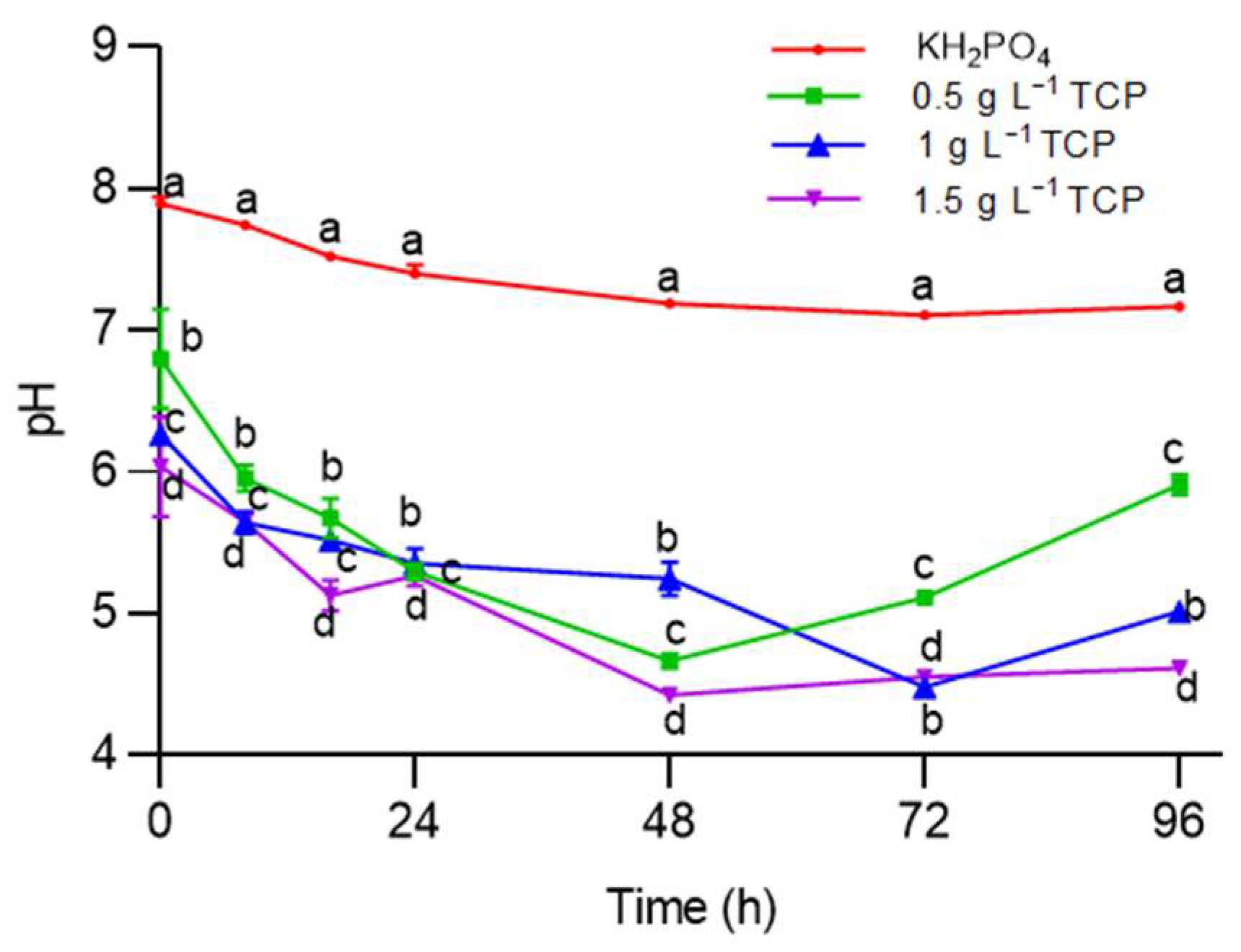
2.2.3. Phosphorus Solubilization
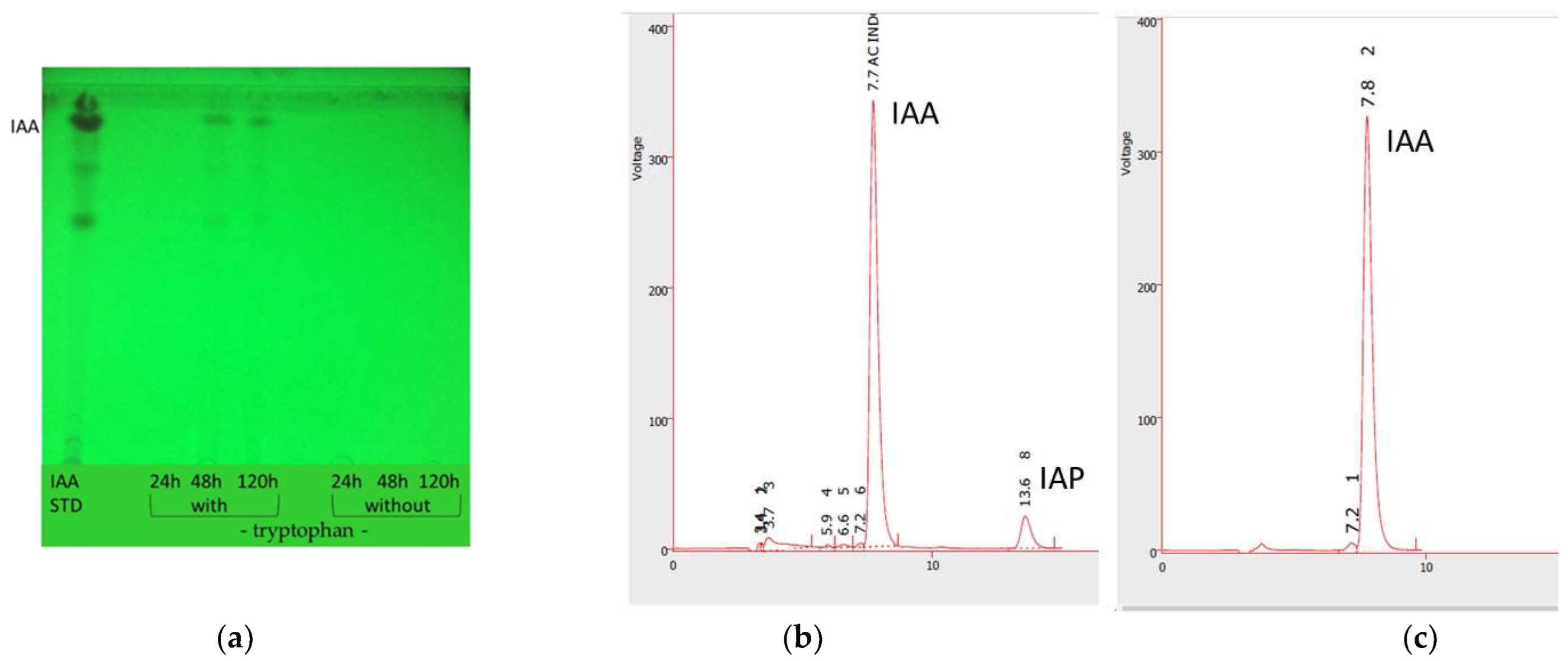
2.2.4. Indole Acetic Acid Production
2.3. Development of Zea mays Seedlings in Fähraeus System
2.3.1. Seed Germination of Zea mays
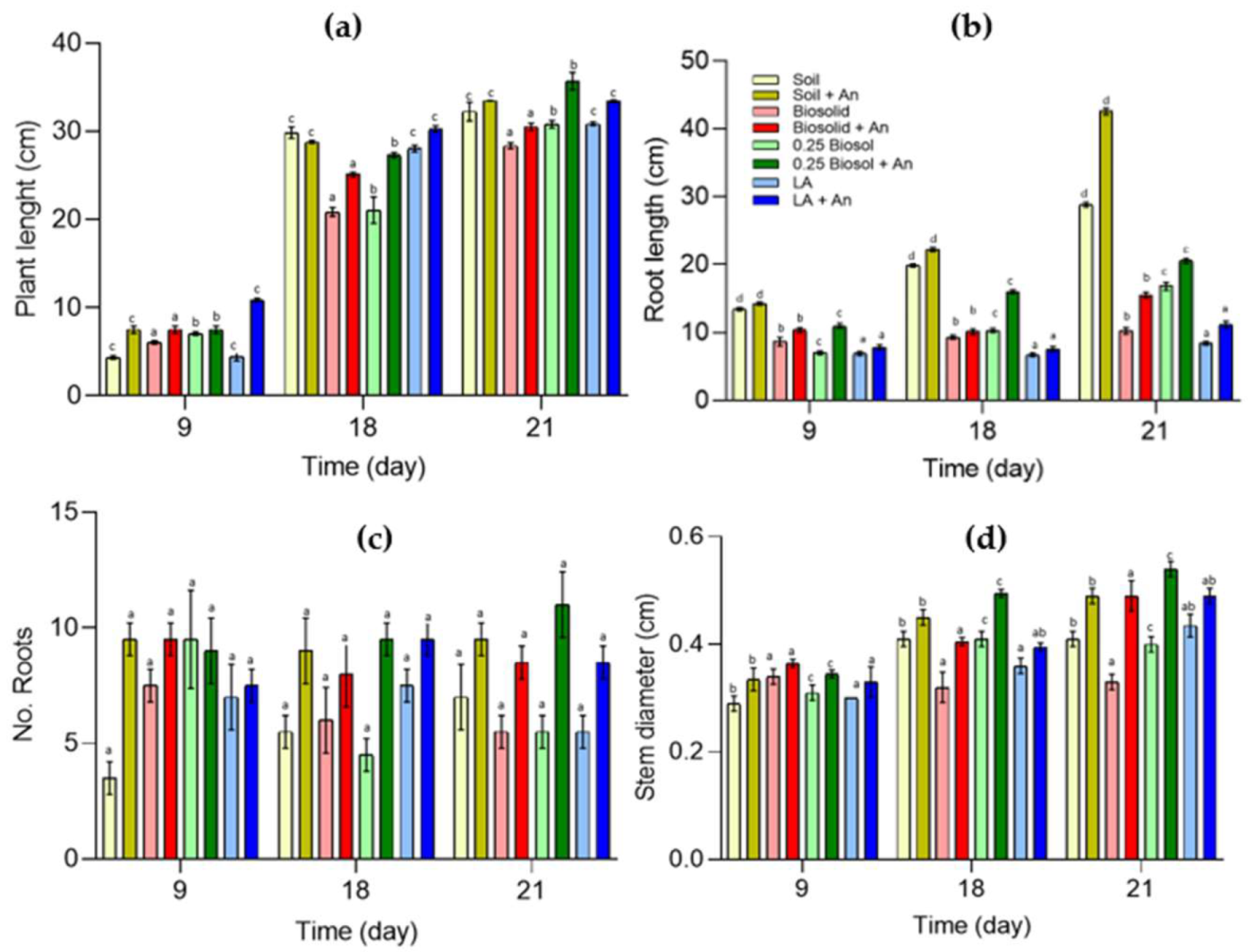
2.3.2. Effect of Biosolids and Azotobacter nigricans on the Development of Zea mays Seedlings
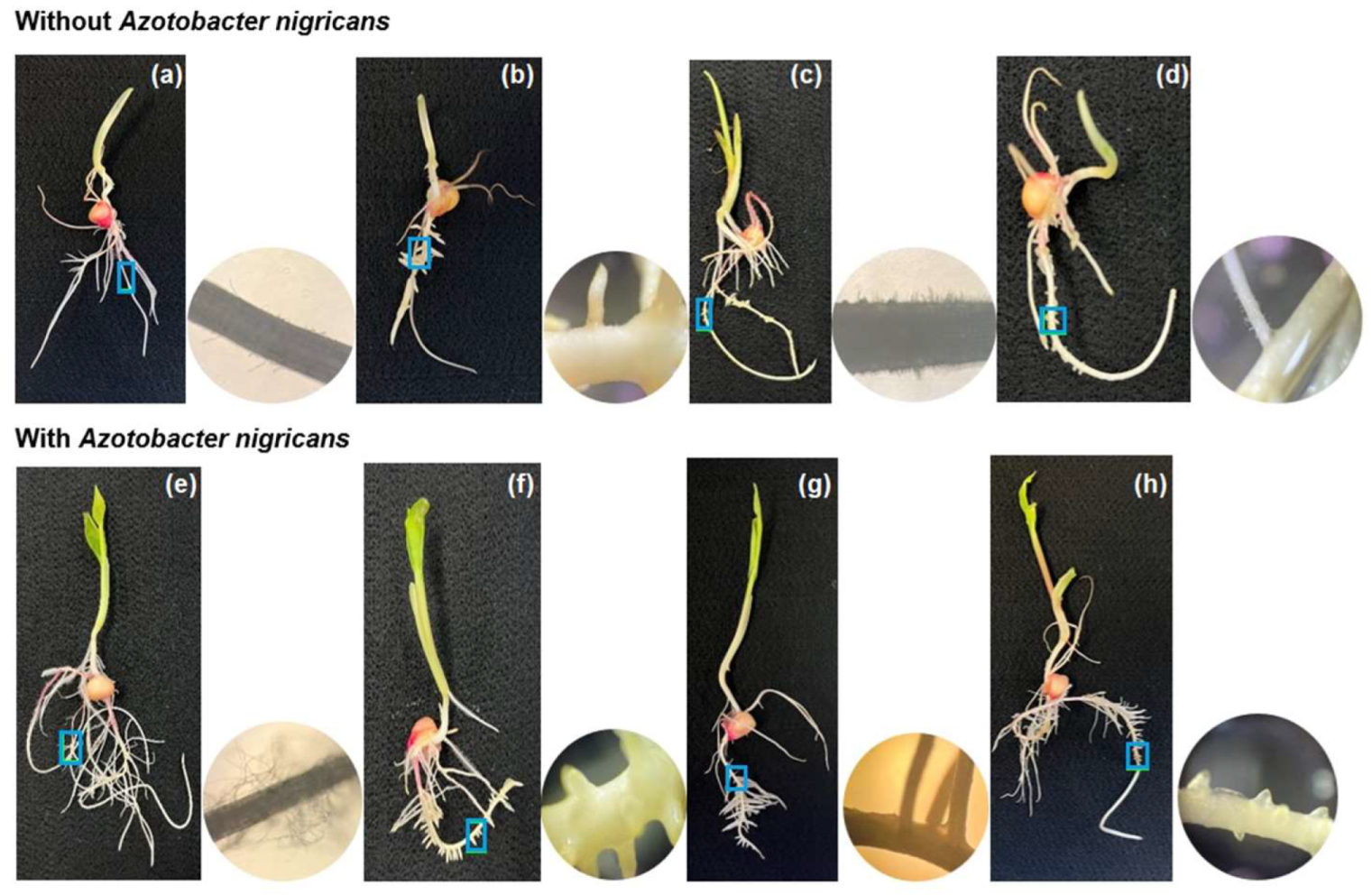
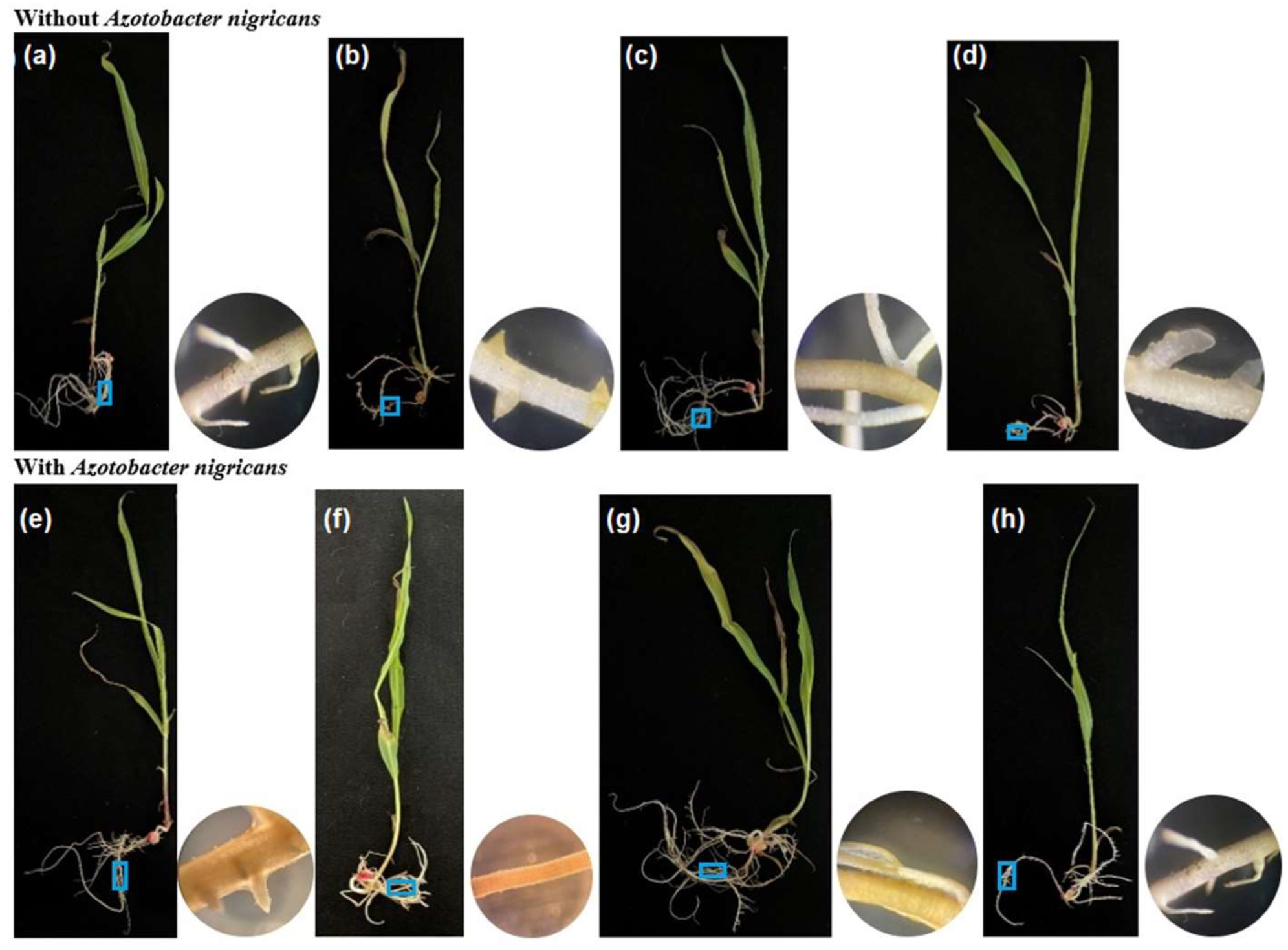
2.4. Colonization of Azotobacter nigricans in the Roots of the Zea mays Plants
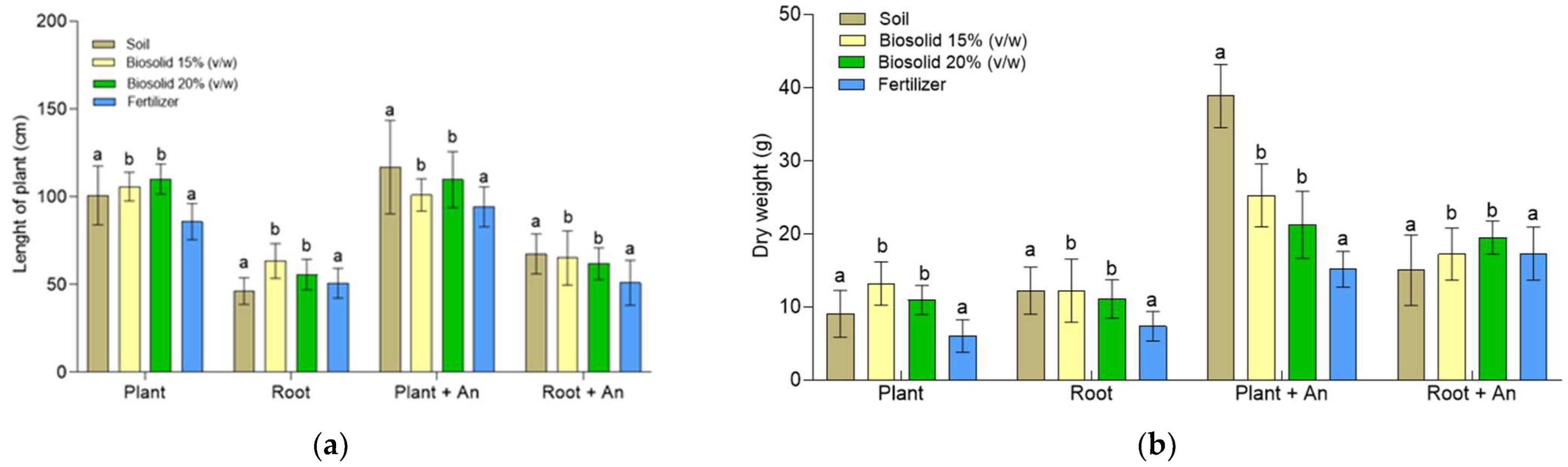
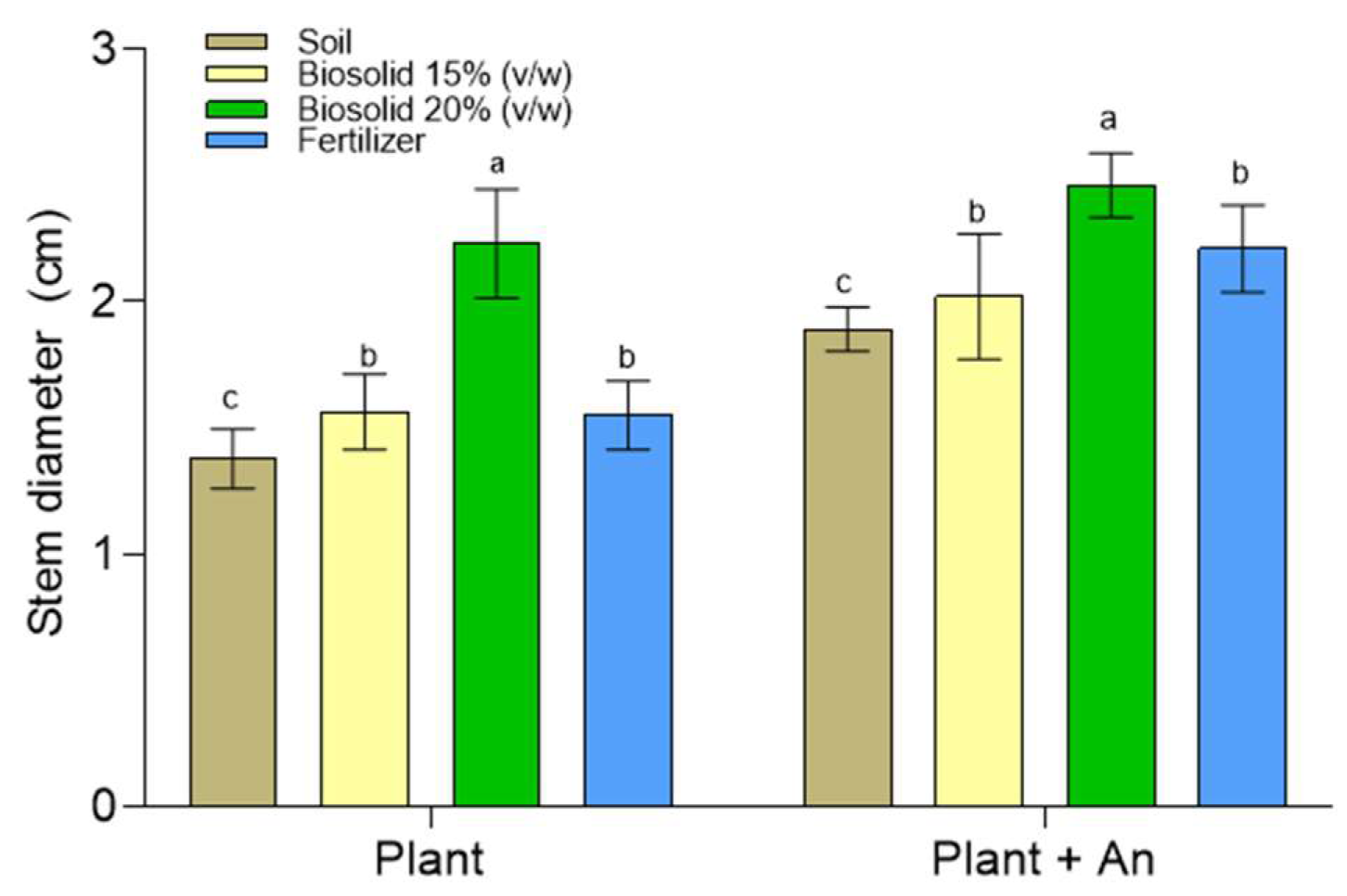
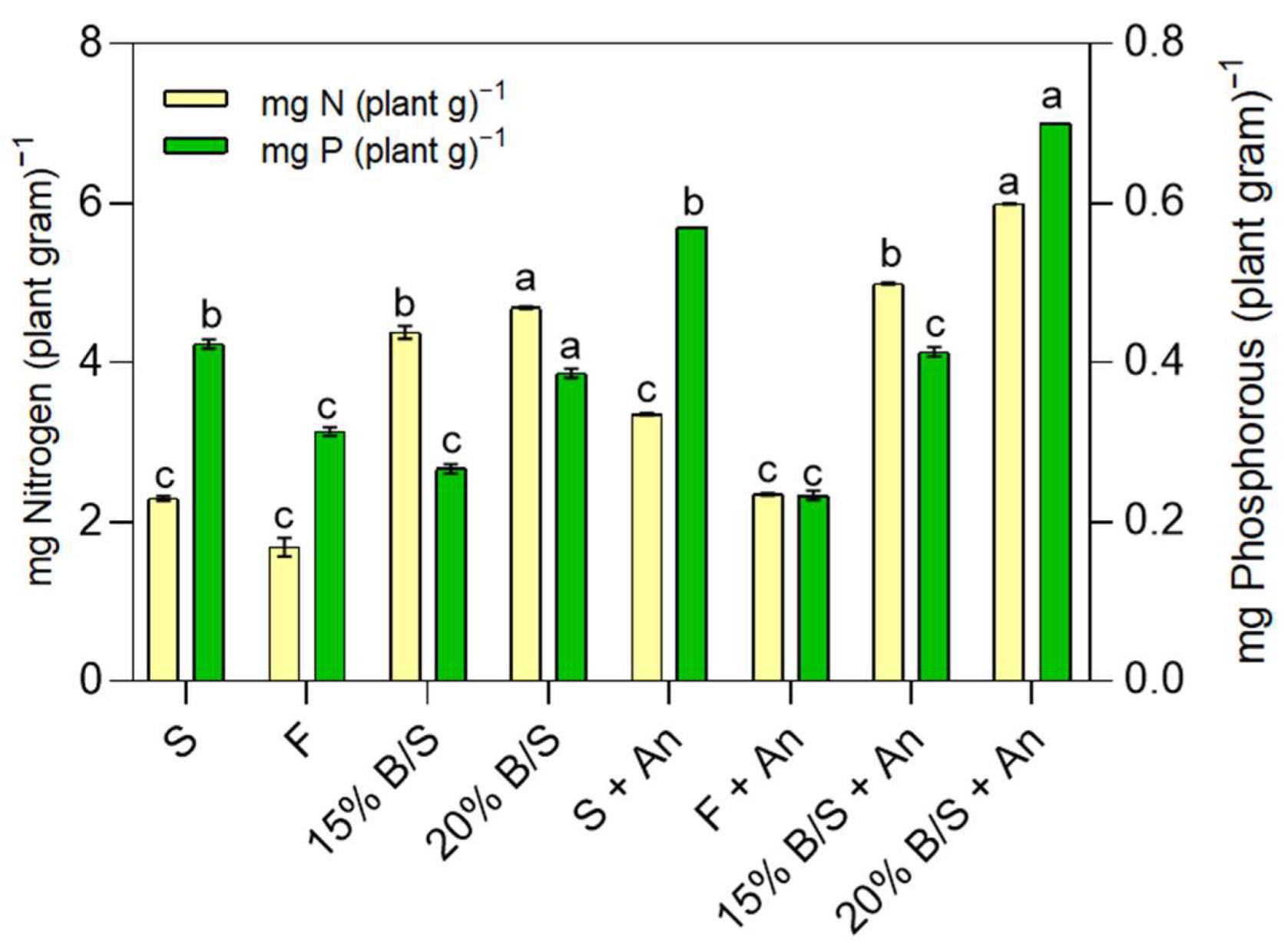
2.5. Growing Zea mays Plants in a Greenhouse
3. Discussion
4. Materials and Methods
4.1. Soil Collection
4.2. Biosolids Collection
4.3. Soil and Biosolids Characterization
4.3.1. pH
4.3.2. Humidity
4.3.3. Soil Texture
4.3.4. Organic Matter
4.3.5. Electrical Conductivity
4.3.6. Total Solids
4.3.7. Total Phosphorus and Total Nitrogen
4.3.8. Total Coliforms
4.3.9. Heavy Metals
4.4. Growth of Azotobacter nigricans in Soil Conditioned with Biosolids
4.5. Nitrogenase Activity in Azotobacter nigricans
4.6. Indole Acetic Acid (IAA) Production by Azotobacter nigricans
4.7. Solubilization of Tricalcium Phosphate by Azotobacter nigricans
4.8. Germination of Zea mays Seeds
4.9. Growth of Zea mays Seedlings in the Fähraeus System
Scanning Electron Microscopy Observation
4.10. Growth of Zea mays Plants in a Greenhouse
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vogel, C.; Sekine, R.; Huang, J.; Steckenmesser, D.; Steffens, D.; Huthwelker, T.; Borca, C.N.; Pradas del Real, A.E.; Castillo-Michel, H.; Adam, C. Effects of a nitrification inhibitor on nitrogen species in the soil and the yield and phosphorus uptake of maize. Sci. Total Environ. 2020, 715, 136895. [Google Scholar] [CrossRef] [PubMed]
- Torri, S.; Correa, R.; Renella, G. Biosolid Application to Agricultural Land—A Contribution to Global Phosphorus Recycle: A Review. Pedosphere 2017, 27, 1–16. [Google Scholar] [CrossRef]
- Sharma, B.; Sarkar, A.; Singh, P.; Singh, R.P. Agricultural utilization of biosolids: A review on potential effects on soil and plant grown. Waste Manag. 2017, 64, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Koutroubas, S.D.; Antoniadis, V.; Damalas, C.A.; Fotiadis, S. Municipal Sewage Sludge Effects on Maize Yield, Nitrogen Use Efficiency, and Soil Properties. J. Soil Sci. Plant Nutr. 2023, 23, 1209–1221. [Google Scholar] [CrossRef]
- Scaci, S. Investigation of effect of chemical fertilizers on environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef]
- Pahalvi, H.; Rafiya, L.; Rashid, S.; Nisar, B.; Kamili, A. Chemical fertilizers and their impact on soil health. In Microbiota and Biofertilizers; Dar, G.H., Bhat, R.A., Mehmood, M.A., Hakeem, K.R., Eds.; Springer: Cham, Switzerland, 2021; Volume 2, pp. 1–20. [Google Scholar] [CrossRef]
- Sullivan, D.M.; Cogger, C.G.; Bary, A. Fertilizing with Biosolids; Pacific Northwest Extension Publication: Portland, OR, USA; Pullman, WA, USA; Bois, IA, USA, 2015; pp. 1–19. [Google Scholar]
- Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). Diagnóstico Básico para la Gestión Integral de los Residuos Primera Edición; Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT): Ciudad de Mexico, Mexico, 2020; pp. 12–28. [Google Scholar]
- Uribe-Montes, H.R.; Chávez-Sánchez, N.; Orozco-Hernández, G.; Espino-Valdez, M.S. Biosólidos digeridos anaeróbicamente en la producción de maíz forrajero. Agric. Técnica México 2003, 29, 25–34. Available online: http://www.redalyc.org/articulo.oa?id=60829103 (accessed on 22 August 2023).
- Cogger, C.G.; Forge, T.A.; Neilsen, G.H. Biosolids recycling: Nitrogen management and soil ecology. Can. J. Soil Sci. 2006, 86, 613–620. [Google Scholar] [CrossRef]
- Badza, T.; Tesfamariam, E.H.; Cogger, C.G. Agricultural use suitability assessment and characterization of municipal liquid sludge: Based on South Africa survey. Sci. Total Environ. 2020, 721, 137658. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; Peach, D. Biosolids in Vegetable Production Systems. HortTechnology 2002, 12, 336–340. [Google Scholar] [CrossRef]
- Melo, W.; Delarica, D.; Guedes, A.; Lavezzo, L.; Donha, R.; de Araújo, A.; de Melo, G.; Macedo, F. Ten years of application of sewage sludge on tropical soil. A balance sheet on agricultural crops and environmental quality. Sci. Total Environ. 2018, 643, 1493–1501. [Google Scholar] [CrossRef]
- Walter, I.; Martínez, F.; Cala, V. Heavy metal speciation and phytotoxic effects of three representative sewage sludges for agricultural uses. Environ. Pollut. 2006, 139, 507–514. [Google Scholar] [CrossRef]
- Kidd, P.S.; Domínguez-Rodríguez, M.J.; Díez, J.; Monterroso, C. Bioavailability and plant accumulation of heavy metals and phosphorus in agricultural soils amended by long-term application of sewage sludge. Chemosphere 2007, 66, 1458–1467. [Google Scholar] [CrossRef]
- Yang, G.H.; Zhu, G.Y.; Li, H.L.; Han, X.M.; Li, J.M.; Ma, Y.B. Accumulation and bioavailability of heavy metals in a soil-wheat/maize system with long-term sewage sludge amendments. J. Integr. Agric. 2018, 17, 1861–1870. [Google Scholar] [CrossRef]
- de Amorim Júnior, S.S.; de Souza Pereira, M.A.; Morishigue, M.; da Costa, R.B.; de Oliveira Guilherme, D.; Magalhães Filho, F.J.C. Circular Economy in the Biosolids Management by Nexus Approach: A View to Enhancing Safe Nutrient Recycling—Pathogens, Metals, and Emerging Organic Pollutants Concern. Sustainability 2022, 14, 14693. [Google Scholar] [CrossRef]
- EUR Lex. Directiva 86/278/CEE del Consejo de 12 de junio de 1986 relativa a la protección del medio ambiente y, en particular, de los suelos, en la utilización de los lodos de depuradora en agricultura. Diario Oficial de Las Comunidades Europeas; pp. 6–12. 1984. Available online: https://eur-lex.europa.eu/ES/legal-content/summary/using-sewage-sludge-in-farming.html (accessed on 22 August 2023).
- USEPA Part 503, Standards for the Use or Disposal of Sewage Sludge. Title 40. Code of Federal Regulations. 1994. Available online: https://www.ecfr.gov/current/title-40/chapter-I/subchapter-O/part-503 (accessed on 8 May 2023).
- Secretaria de Medio Ambiente y Recursos Naturales (SEMARNAT). Protección ambiental. Lodos y Biosólidos. Especificaciones y límites máximos permisibles de contaminantes para su aprovechamiento y disposición final (NOM-004-SEMARNAT-2002): México, D.F., Diario Oficial de la Federación, viernes 15 de agosto del. 2002. Available online: https://dof.gob.mx/nota_detalle.php?codigo=691939&fecha=15/08/2003#gsc.tab=0 (accessed on 22 August 2023).
- Bıyıklı, M.; Dorak, S.; Aşık, B.B. Effects of Food Industry Wastewater Treatment Sludge on Corn Plant Development and Soil Properties. Pol. J. Environ. Stud. 2020, 29, 2565–2578. [Google Scholar] [CrossRef]
- Santacoloma-Londoño, S.P.; Buitrago-González, M.E.; Colorado-Molina, K.; Suárez-Pineda, I.; Martínez-Martina, M.A.; Villegas-Méndez, L.C. Agricultural Use of Biosolids Generated in Wastewater Treatment of a Food Industry. Rev. Fac. Ing. 2020, 29, e10666. [Google Scholar] [CrossRef]
- Marchuk, S.; Tait, S.; Sinha, P.; Harris, P.; Antille, D.L.; McCabe, B.K. Biosolids-derived fertilisers: A review of challenges and opportunities. Sci. Total Environ. 2023, 875, 162555. [Google Scholar] [CrossRef] [PubMed]
- Abdenaceur, R.; Farida, B.T.; Mourad, D.; Rima, H.; Zahia, O.; Fatma, S.-H. Effective biofertilizer Trichoderma spp. isolates with enzymatic activity and metabolites enhancing plant growth. Int. Microbiol. 2022, 25, 817–829. [Google Scholar] [CrossRef]
- dos Santos Lopes, M.; Dias-Filho, M.; Gurgel Cajueiro, E. Successful Plant Growth-Promoting Microbes: Inoculation Methods and Abiotic Factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Ansari, R.A.; Rizvi, R.; Sumbul, A.; Mahmood, I. PGPR: Current Vogue in Sustainable Crop Production. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; Volume 1, pp. 455–472. [Google Scholar] [CrossRef]
- Glick, B.R.; Gamalero, E. Recent developments in the study of plant microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.A.; Chand, S.; Ali, T. Potential Use of Azotobacter chroococcum in Crop Production: An Overview. Curr. Agric. Res. 2013, 1, 35–38. [Google Scholar] [CrossRef]
- Das, H. Azotobacters as biofertilizer. In Advances in Applied Microbiology; Gadd, G.M., Sariaslani, S., Eds.; Academic Press: London, UK, 2019; Volume 108, pp. 1–43. [Google Scholar] [CrossRef]
- Patil, S.V.; Mohite, B.; Patil, C.D.; Koli, S.H.; Borase, H.P.; Patil, V.S. Azotobacter. In Beneficial Microbes in Agro-Ecology; Amaresan, N., Senthil Kumar, M., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: London, UK, 2020; pp. 397–426. [Google Scholar] [CrossRef]
- Sagar, A.; Sayyed, R.Z.; Ramteke, P.W.; Ramakrishna, W.; Poczai, P.; Al Obaid, S.; Ansari, M.J. Synergistic Effect of Azotobacter nigricans and Nitrogen Phosphorus Potassium Fertilizer on Agronomic and Yieldtraits of Maize (Zea mays L.). Front. Plant Sci. 2022, 13, 952212. [Google Scholar] [CrossRef]
- FAOSTAT. Maize Production in 2020. Food and Agriculture Organization of the United Nations. 2021. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 8 May 2023).
- Sithole, N.J.; Magwaza, L.S.; Mafongoya, P.L. Conservation agriculture and its impact on soil quality and maize yield: A South African perspective. Soil Tillage Res. 2016, 162, 55–67. [Google Scholar] [CrossRef]
- Bozkurt, M.A.; Akdeniz, H.; Keskin, B.; Yilmaz, I.H. Possibilities of using sewage sludge as nitrogen fertilizer for maize. Acta Agric. Scand. B Soil Plant Sci. 2006, 56, 143–149. [Google Scholar] [CrossRef]
- Li, X.; Lu, Q.; Li, D.; Wang, D.; Ren, X.; Yan, J.; Ahmed, T.; Li, B. Effects of Two Kinds of Commercial Organic Fertilizers on Growth and Rhizosphere Soil Properties of Corn on New Reclamation Land. Plants 2022, 11, 2553. [Google Scholar] [CrossRef]
- Battini, F.; Grønlund, M.; Agnolucci, M.; Giovannetti, M.; Jakobsen, I. Facilitation of phosphorus uptake in maize plants by mycorrhizosphere bacteria. Sci. Rep. 2017, 7, 4686. [Google Scholar] [CrossRef]
- Benbrik, B.; Elabed, A.; El Modafar, C.; Douira, A.; Amir, S.; Filali-Maltouf, A.; El Abed, S.; El Gachtouli, N.; Mohammed, I.; Koraichi, S.I. Reusing phosphate sludge enriched by phosphate solubilizing bacteria as biofertilizer: Growth promotion of Zea Mays. Biocatal. Agric. Biotechnol. 2020, 30, 101825. [Google Scholar] [CrossRef]
- Jameson, M.; White, J.G.; Osmond, D.L.; Aziz, T. Determination of Biosolids Phosphorus Solubility and Its Relationship to Wastewater Treatment. Water Environ. Res. 2016, 88, 602–610. [Google Scholar] [CrossRef]
- Ahmad, A.; Ugya, A.; Isah, H.; Imam, T. Mineralization and mobilization of biosolids phosphorus in soil: A concise review. J. Appl. Biol. Biotechnol. 2019, 7, 98–106. [Google Scholar] [CrossRef]
- Peck, T.R.; Soltanpour, P.M. Principles of soil testing. In Soil Testing and Plant Analysis, 3rd ed.; Westerman, R.L., Ed.; SSSA: Madison, WI, USA, 1990; pp. 80–85. [Google Scholar] [CrossRef]
- Pattison, T.; Moody, P.; Bagshaw, J. Soil Health for Vegetable Production in Australia; The State of Queensland, Department of Employment, Economic Development and Innovation: Brisbane, Australia, 2010; pp. 5–45. [Google Scholar]
- Anagnostopolus, A.; Vaiopoulou, E. Assessment of Chemical Oxygen Demand/Total Organic Carbon (COD/TOC) Ratios in Refinery Effluents (Issue 16); Concawe: Brussels, Belgium, 2022; pp. 5–14. [Google Scholar]
- Wang, X.; Sun, C.; Gao, S.; Wang, L.; Shuokui, H. Validation of germination rate and root elongation as indicator to assess phytotoxicity with Cucumis sativus. Chemosphere 2001, 44, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Freeman, S. Plant Nutrition. In Biological Science, 3rd ed.; Pearson/Benjamin Cummings: San Francisco, CA, USA, 2008; pp. 837–850. [Google Scholar]
- Kacprzak, M.; Neczaj, E.; Fijałkowski, K.; Grobelak, A.; Grosser, A.; Worwag, M.; Rorat, A.; Brattebo, H.; Almås, Å.; Singh, B.R. Sewage sludge disposal strategies for sustainable development. Environ. Res. 2017, 156, 39–46. [Google Scholar] [CrossRef]
- Bloom, P.R.; Skyllberg, U.L.; Sumner, M.E. Soil acidity. In Chemical Processes in Soils; Tabatabai, M.A., Sparks, D.L., Eds.; SSSA: Madison, WI, USA, 2005; pp. 411–459. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Corwin, D.L.; Lesch, S.M. Apparent soil electrical conductivity measurements in agriculture. Comput. Electron. Agric. 2005, 46, 11–43. [Google Scholar] [CrossRef]
- Saxton, K.E.; Rawls, W.J. Soil Water Characteristic Estimates by Texture and Organic Matter for Hydrologic Solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef]
- Al-Kaisi, M.M.; Lal, R. Conservation Agriculture Systems to Mitigate Climate Variability Effects on Soil Health. In Soil Health and Intensification of Agroecosytems; Al-Kaisi, M.M., Lowery, B., Eds.; Academic Press: Amsterdam, The Netherlands, 2017; pp. 79–107. [Google Scholar] [CrossRef]
- Saha-Pal, S.; Bhattacharyya, S.; Chakraborty, H. Solubilization of tricalcium phosphate by P(3HB) accumulating Azotobacter chroococcum MAL-201. World J. Microbiol. Biotechnol. 2014, 30, 1575–1582. [Google Scholar] [CrossRef]
- Havlin, J.L. Fertility. In Reference Module in Earth Systems and Environmental Sciences; Elsevier North Carolina State University: Raleigh, NC, USA, 2013; pp. 1–12. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; pp. 67–81. [Google Scholar]
- Roberts, D.A.; Cole, A.J.; Whelan, A.; de Nys, R.; Paul, N.A. Slow pyrolysis enhances the recovery and reuse of phosphorus and reduces metal leaching from biosolids. Waste Manag. 2017, 64, 133–139. [Google Scholar] [CrossRef]
- López-Mosquera, M.E.; Moirón, C.; Carral, E. Use of dairy-industry sludge as fertiliser for grasslands in northwest Spain: Heavy metal levels in the soil and plants. Resour. Conserv. Recycl. 2000, 30, 95–109. [Google Scholar] [CrossRef]
- Nosrati, R.; Owlia, P.; Saderi, H.; Rasooli, I.; Malboobi, M.A. Phosphate solubilization characteristics of efficient nitrogen fixing soil Azotobacter strains. Iran. J. Microbiol. 2014, 6, 285–295. [Google Scholar]
- Haahtela, K.; Kari, K.; Sundman, V. Nitrogenase activity (acetylene reduction) of root-associated, cold-climate Azospirillum, Enterobacter, Klebsiella and Pseudomonas species during growth on various carbon sources and at various partial pressures of oxygen. Appl. Environ. Microbiol. 1983, 45, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- García-Esquivel, G.; Calva-Calva, G.; Ferrera-Cerrato, R.; Fernández-Linares, L.C.; Rodríguez-Vázquez, R.; Esparza-García, F.J. Encystment of Azotobacter nigricans grown diazotrophically on kerosene as sole carbon source. Arch. Microbiol. 2009, 191, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Bhatnagar, A.; Kalla, A.; Narula, N. In vitro nitrogen fixation, phosphate solubilization, survival and nutrient release by Azotobacter strains in an aquatic system. Bioresour. Technol. 2001, 80, 101–109. [Google Scholar] [CrossRef]
- Singh, D.; Thapa, S.; Geat, N.; Mehriya, M.L.; Rajawat, M.V. Biofertilizers: Mechanisms and application. Biofertilizers 2021, 1, 151–166. [Google Scholar] [CrossRef]
- Paredes-Mendoza, M.; Espinosa-Victoria, D. Organic Acids Produced by Phosphate Solubilizing Rhizobacteria: A Critical Review. Terra Latinoam. 2010, 28, 61–70. [Google Scholar]
- Frankenberger, W.T.; Brunner, W. Method of Detection of Auxin-Indole-3-Acetic Acid in Soils by High Performance Liquid Chromatography. Soil Sci. Soc. Am. J. 1983, 47, 237–241. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Indole Acetic Acid Production by the Indigenous Isolates of Azotobacter and Fluorescent Pseudomonas in the Presence and Absence of Tryptophan. Turk. J. Biol. 2005, 29, 29–34. Available online: https://journals.tubitak.gov.tr/biology/vol29/iss1/5 (accessed on 22 August 2023).
- Carvalhais, L.C.; Dennis, P.G.; Fedoseyenko, D.; Hajirezaei, M.R.; Borriss, R.; Von Wirén, N. Root exudation of sugars, amino acids, and organic acids by maize as affected by nitrogen, phosphorus, potassium, and iron deficiency. J. Plant Nutr. Soil Sci. 2011, 174, 3–11. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, J.R. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Pardo-García, J.M.; Martínez-Salgado, M.M.; Montaña-Lara, J.S. Bio-fertilizer production from an isolate of Azotobacter nigricans obtained from a plantation of Stevia rebaudiana Bert. Univ. Sci. 2009, 14, 71–78. [Google Scholar] [CrossRef]
- Cui, H.; Cheng, Z.; Li, P.; Miao, A. Prediction of Sweet Corn Seed Germination Based on Hyperspectral Image Technology and Multivariate Data Regression. Sensors 2020, 20, 4744. [Google Scholar] [CrossRef] [PubMed]
- Khaeim, H.; Kende, Z.; Jolánkai, M.; Kovács, G.P.; Gyuricza, C.; Tarnawa, Á. Impact of Temperature and Water on Seed Germination and Seedling Growth of Maize (Zea mays L.). Agronomy 2022, 12, 397. [Google Scholar] [CrossRef]
- Fähraeus, G. The Infection of Clover Root Hairs by Nodule Bacteria Studied by a Simple Glass Slide Technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar Behl, R.; Narula, N. Establishment of phosphate-solubilizing strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat cultivars under greenhouse conditions. Microbiol. Res. 2001, 156, 87–93. [Google Scholar] [CrossRef]
- Nadira, U.A.; Ahmed, I.M.; Wu, F.; Zhang, G. The regulation of root growth in response to phosphorus deficiency mediated by phytohormones in a Tibetan wild barley accession. Acta Physiol. Plant. 2016, 38, 105. [Google Scholar] [CrossRef]
- López-Bucio, J.; Cruz-Ramírez, A.; Herrera-Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 2003, 6, 280–287. [Google Scholar] [CrossRef]
- Bibikova, T.; Gilroy, S. Root Hair Development. J. Plant Growth Regul. 2002, 21, 383–415. [Google Scholar] [CrossRef]
- Ditengou, F.; Béguiristain, T.; Lapeyrie, F. Root hair elongation is inhibited by hypaphorine, the indole alkaloid from the ectomycorrhizal fungus Pisolithus tinctorius, and restored by indole-3-acetic acid. Planta 2000, 211, 722–728. [Google Scholar] [CrossRef]
- Costa, C.; Dwyer, L.M.; Zhou, X.; Dutilleul, P.; Hamel, C.; Reid, L.M.; Smith, D.L. Root Morphology of Contrasting Maize Genotypes. Agron. J. 2002, 94, 96–101. [Google Scholar] [CrossRef]
- Gurikar, C.; Sreenivasa, M.Y.; Gowda, N.A.N.; Lokesh, A.C. Chapter 6—Azotobacter—A potential symbiotic rhizosphere engineer. In Rhizosphere Engineering; Dubey, R.C., Kumar, P., Eds.; Academic Press: Cambridge, MA, USA, 2022; Volume 1, pp. 97–112. [Google Scholar] [CrossRef]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.L.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef]
- Gu, R.; Chen, F.; Long, L.; Cai, H.; Liu, Z.; Yang, J.; Wang, L.; Li, H.; Li, J.; Liu, W.; et al. Enhancing phosphorus uptake efficiency through QTL-based selection for root system architecture in maize. J. Genet. Genom. 2016, 43, 663–672. [Google Scholar] [CrossRef]
- González-Flores, E.; Ramos-Barragán, J.E.; Tornero-Campante, M.A.; Murillo-Murillo, M. Evaluación de dosis de biosólidos urbanos en maíz bajo condiciones de invernadero. Rev. Mex. Cienc. Agrícolas 2017, 8, 119–132. [Google Scholar] [CrossRef]
- Elizondo-Barrón, J.; Castillo-Tovar, H.; Cano-Pineda, A.; Loera-Gallardo, J.; Rodríguez-Guerra, R.; Terán-Vargas, A.P.; Maldonado-Moreno, N.; Patishtan-Pérez, J. Muestreo de Suelos y Preparación de Muestras. 2012. Available online: http://www.inifapcirne.gob.mx/Biblioteca/Publicaciones/935.pdf (accessed on 13 May 2023).
- Zagal, E.; Sadzawka, A. Protocolo de Métodos de Análisis Para Suelos y Lodos; Comisión de Normalización y Acreditación de la Sociedad Chilena de la Ciencia del Suelo: Concepción, Chile, 2007. [Google Scholar]
- González-Naranjo, V.; Leal, M.L.J.; De Bustamante, I. Guía de Caracterización Edáfica Para Actividades de Regeneración de Aguas Residuales en Usos Ambientales; Consolider Tragua: Madrid, España, 2012. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. (Eds.) APHA Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association (APHA), American Water Works Association (AWWA) and Water Environment Federation (WEF): Washington, DC, USA, 2012; pp. 154–165, 202–206, 216–218. [Google Scholar]
- Hach. Total nitrogen, method 10072, and total phosphate, method 10127. In Water Analysis Handbook, 9th ed.; Hach: Ames, IA, USA, 2014. [Google Scholar]
- SEMARNAT. NOM-113-SSA1-1994, Bienes y Servicios. Método para la Cuenta de Microorganismos Coliformes Totales en placa; SEMARNAT: Ciudad de Mexico, Mexico, 1994. [Google Scholar]
- García-Esquivel, G. Papel Fisiológico de Azotobacter Durante la Remoción de Queroseno en la Rizósfera. Ph.D. Thesis, Centro de Investigación y Estudios Avanzados del IPN, Departamento de Biotecnología, Ciudad de Mexico, Mexico, 2008. [Google Scholar]
- Postgate, J.R. The acetylene reduction test for nitrogen Wxation. In Methods in Microbiology, 6B ed.; Norris, J.R., Ribbons, D.W., Eds.; Academic Press: London, UK, 1971; pp. 343–356. [Google Scholar]
- Smith, G.S.; Johnston, C.M.; Cornforth, I.S. Comparison of Nutrient Solutions for Growth of Plants in Sand Culture. New Phytol. 1983, 94, 537–548. [Google Scholar] [CrossRef]
- Hu, Y.; Barker, A.V. A single plant tissue digestion for macronutrient analysis. Commun. Soil Sci. Plant Anal. 1999, 30, 677–687. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Agricultural Soil | Biosolids from the Food Industry | Ceiling Concentrations [19] |
|---|---|---|---|
| pH | 8.10 ± 0.12 | 6.80 ± 0.03 | |
| Electric conductivity (EC), dS m−1 | 1.05 ± 0.30 | - | |
| Humidity (H), % | 27.5 ± 0.1 | - | |
| Organic matter (OM), % | 8.70 ± 0.02 | 0.87 ± 0.20 | |
| Cation exchange (CEC) cmol kg−1 | 32.00 ± 0.01 | - | |
| Texture | sandy loam | - | |
| Clay, % | 0.17 | - | |
| Slime, % | 28.74 | - | |
| Sand, % | 71.09 | - | |
| Chemical parameters | |||
| NO3-, mg kg−1 | 11.32 ± 0.10 | 320.10 ± 0.03 | |
| Total nitrogen (N), mg kg−1 | 609.42 ± 0.10 | 1723.20 ± 0.03 | |
| PO4−3, mg kg−1 | 4.44 ± 0.02 | 27.58 ± 0.60 | |
| Total phosphorus (P), mg kg−1 | 13.62 ± 0.02 | 84.58 ± 0.60 | |
| Metals | |||
| As, mg kg−1 | 1.78 ± 0.12 | 1.12 ± 0.08 | 75 |
| Cd, mg kg−1 | <4 ± N.A. | 4.19 ± 0.32 | 85 |
| Cr, mg kg−1 | <20 ± N.A. | <20 ± N.A. | - |
| Cu, mg kg−1 | 17.14 ± 0.91 | <10 ± N.A. | 4300 |
| Pb, mg kg−1 | 23.61 ± 1.71 | 48.12 ± 3.49 | 840 |
| Hg, mg kg−1 | 0.39 ± 0.04 | 1.98 ± 0.79 | 57 |
| Ni, mg kg−1 | <20 ± N.A. | <20 ± N.A. | 420 |
| Zn, mg kg−1 | 65.96 ± 3.12 | 34.74 ± 1.64 | 7500 |
| Total solids (TSs), g L−1 | - | 15.90 ± 0.04 | - |
| Total volatile solids (TVSs), g L−1 | - | 9.99 ± 0.50 | - |
| Total organic carbon (TOC), mg L−1 | - | 631.00 ± 0.01 | - |
| Chemical oxygen demand (COD), mg L−1 | - | 6050 ± 4 | - |
| Total coliforms as colony-forming units (CFU), CFU mL−1 | - | 1.19 × 10−5 ± 0.01 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vera-García, S.-L.; Rodríguez-Casasola, F.-N.; Barrera-Cortés, J.; Albores-Medina, A.; Muñoz-Páez, K.M.; Cañizares-Villanueva, R.-O.; Montes-Horcasitas, M.-C. Enhancing Phosphorus and Nitrogen Uptake in Maize Crops with Food Industry Biosolids and Azotobacter nigricans. Plants 2023, 12, 3052. https://doi.org/10.3390/plants12173052
Vera-García S-L, Rodríguez-Casasola F-N, Barrera-Cortés J, Albores-Medina A, Muñoz-Páez KM, Cañizares-Villanueva R-O, Montes-Horcasitas M-C. Enhancing Phosphorus and Nitrogen Uptake in Maize Crops with Food Industry Biosolids and Azotobacter nigricans. Plants. 2023; 12(17):3052. https://doi.org/10.3390/plants12173052
Chicago/Turabian StyleVera-García, Sara-Luz, Felipe-Neri Rodríguez-Casasola, Josefina Barrera-Cortés, Arnulfo Albores-Medina, Karla M. Muñoz-Páez, Rosa-Olivia Cañizares-Villanueva, and Ma.-Carmen Montes-Horcasitas. 2023. "Enhancing Phosphorus and Nitrogen Uptake in Maize Crops with Food Industry Biosolids and Azotobacter nigricans" Plants 12, no. 17: 3052. https://doi.org/10.3390/plants12173052