
Exogenous Easily Extractable Glomalin-Related Soil Protein Stimulates Plant Growth by Regulating Tonoplast Intrinsic Protein Expression in Lemon

Abstract
:1. Introduction
2. Results
2.1. Growth Status and Biomass
2.2. Root Morphology
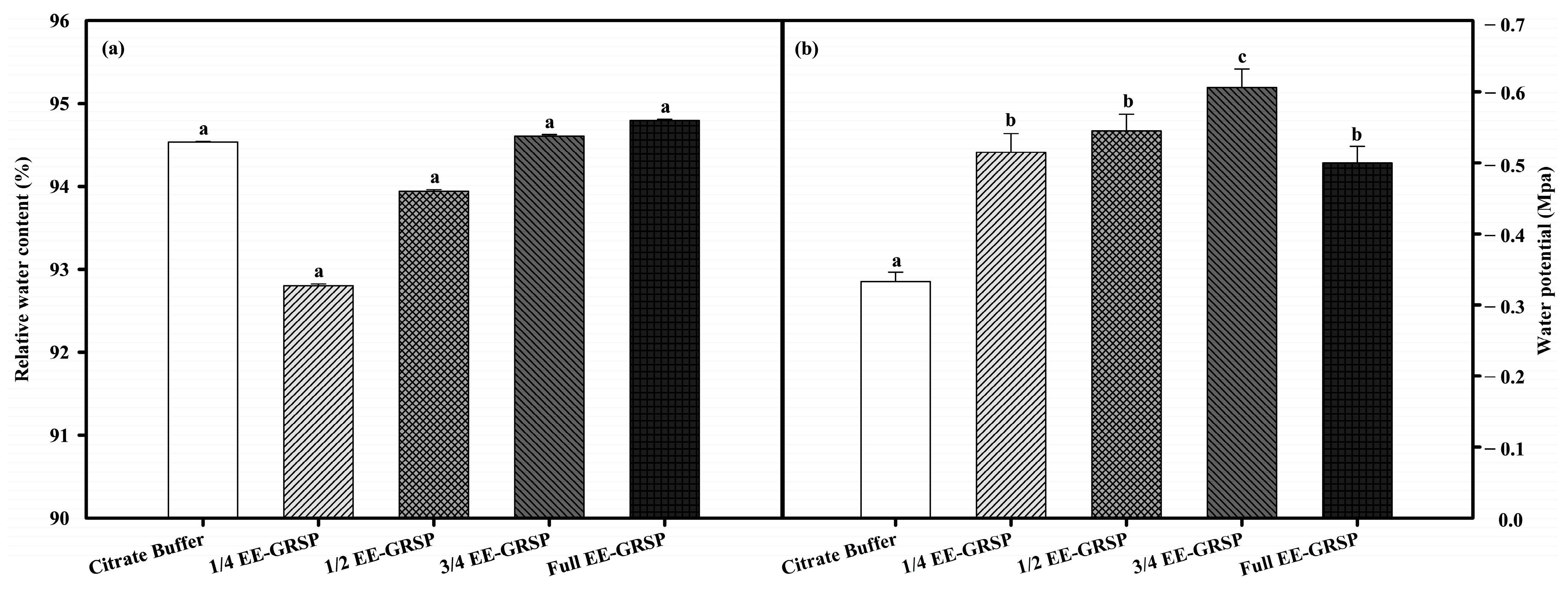
2.3. Leaf Water Status
2.4. Mineral Element
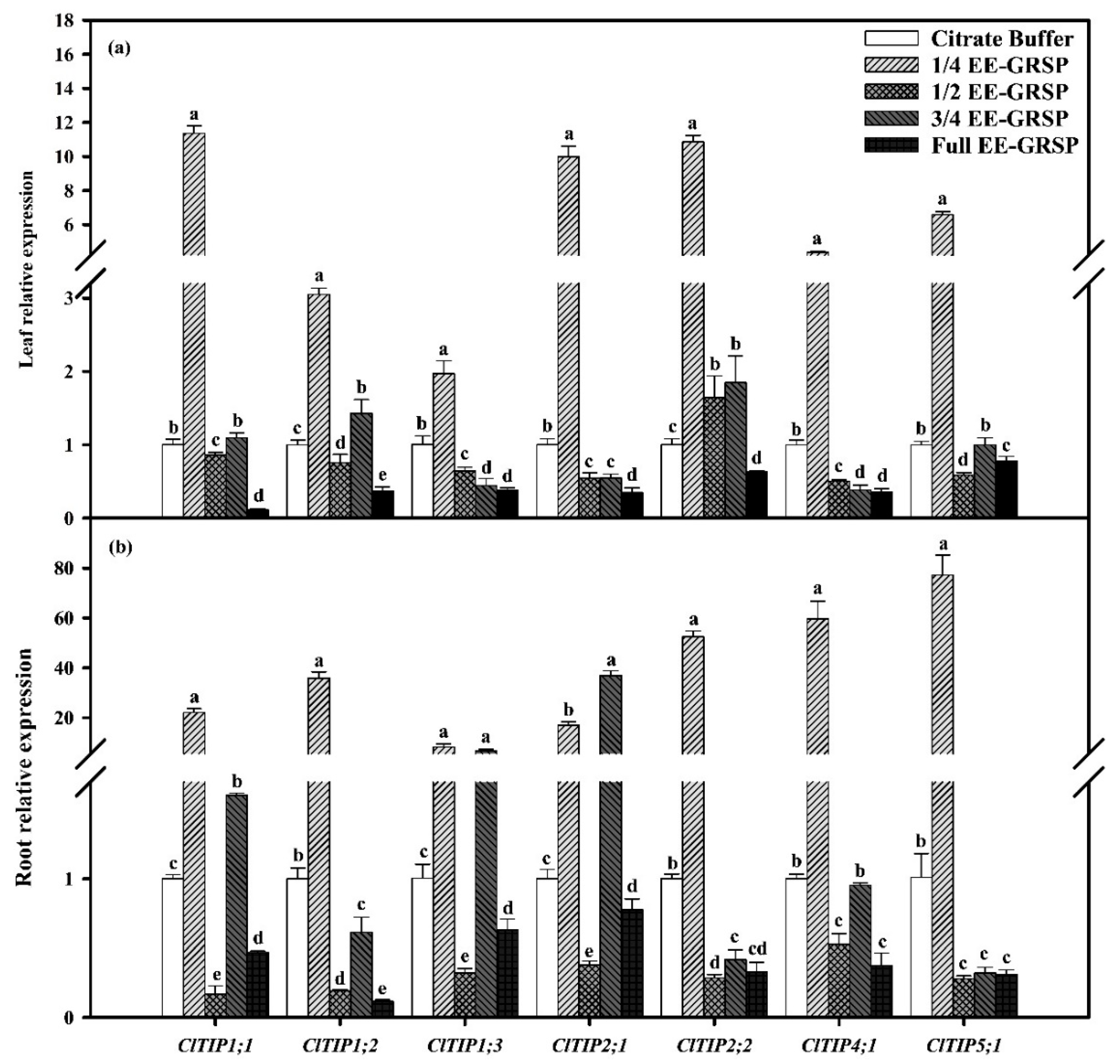
2.5. Relative Expression of TIPs
3. Discussion
4. Material and Methods
4.1. Experimental Material
4.2. Experimental Design
4.3. Variable Determinations
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wright, S.-F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Wright, S.-F.; Upadhyaya, A.; Buyer, J.-S. Comparison of N-linked oligosaccharides of glomalin from arbuscular mycorrhizal fungi and soils by capillary electrophoresis. Soil Biol. Biochem. 1998, 30, 1853–1857. [Google Scholar] [CrossRef]
- Wright, S.-F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Rillig, M.-C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.-S.; Cao, M.-Q.; Zou, Y.-N.; He, X.-H. Direct and indirect effects of glomalin, mycorrhizal hyphae and roots on aggregate stability in rhizosphere of trifoliate orange. Sci. Rep. 2014, 4, 5823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, Q.-S.; He, X.-H. Exogenous easily extractable glomalin-related soil protein promotes soil aggregation, relevant soil enzyme activities and plant growth in trifoliate orange. Plant Soil Environ. 2015, 61, 66–71. [Google Scholar] [CrossRef]
- Chi, G.-G.; Srivastava, A.-K.; Wu, Q.-S. Exogenous easily extractable glomalin-related soil protein improves drought tolerance of trifoliate orange. Arch. Agron. Soil Sci. 2018, 64, 1341–1350. [Google Scholar] [CrossRef]
- Liu, R.-C.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.-F.; Wu, Q.-S. Exogenous glomalin-related soil proteins differentially regulate soil properties in trifoliate orange. Agronomy 2021, 11, 1896. [Google Scholar] [CrossRef]
- Nichols, K.-A. Indirect contributions of AM fungi and soil aggregation to plant growth and protection. In Mycorrhizae: Sustainable Agriculture and Forestry; Springer: Dordrecht, The Netherlands, 2008; pp. 177–194. [Google Scholar] [CrossRef]
- Edwards, J.; Laskowski, M.; Baskin, T.-I.; Mitchell, N.; DeMeo, B. The role of water in fast plant movements. Integr. Comp. Biol. 2019, 59, 1525–1534. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, Z.; Liu, F.; Sun, L.; Hao, F. Versatile roles of aquaporins in plant growth and development. Int. J. Mol. Sci. 2020, 21, 9485. [Google Scholar] [CrossRef]
- Danielson, J.-Å.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol. 2008, 8, 45–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beebo, A.; Thomas, D.; Der, C.; Sanchez, L.; Leborgne-Castel, N.; Marty, F.; Schoefs, B.; Bouhidel, K. Life with and without AtTIP1; 1, an Arabidopsis aquaporin preferentially localized in the apposing tonoplasts of adjacent vacuoles. Plant Mol. Biol. 2009, 70, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-F.; Yamaji, N.; Mitani, N.; Xu, X.-Y.; Su, Y.-H.; McGrath, S.-P.; Zhao, F.-J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [PubMed]
- Uehlein, N.; Fileschi, K.; Eckert, M.; Bienert, G.-P.; Bertl, A.; Kaldenhoff, R. Arbuscular mycorrhizal symbiosis and plant aquaporin expression. Phytochemistry 2007, 68, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Porcel, R.; Aroca, R.; Azcon, R.; Ruiz-Lozano, J.-M. PIP aquaporin gene expression in arbuscular mycorrhizal Glycine max and Lactuca sativa plants in relation to drought stress tolerance. Plant Mol. Biol. 2006, 60, 389–404. [Google Scholar] [CrossRef]
- Zézé, A.; Brou, Y.-C.; Meddich, A.; Marty, F. Molecular identification of MIP genes expressed in the roots of an arbuscular mycorrhizal Trifolium alexandrium L. under water stress. Afr. J. Agric. Res. 2008, 3, 78–83. [Google Scholar] [CrossRef]
- He, J.-D.; Dong, T.; Wu, H.-H.; Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Mycorrhizas induce diverse responses of root TIP aquaporin gene expression to drought stress in trifoliate orange. Sci. Hortic-Amsterdam. 2019, 243, 64–69. [Google Scholar] [CrossRef]
- Lei, A.-Q.; Yang, Q.-H.; Zhang, Y.; Liao, W.-Y.; Xie, Y.-C.; Srivastava, A.-K.; Hashem, A.; Alqahtani, M.-D.; Abd_Allah, E.-F.; Wu, Q.-S.; et al. Agronomic Practices Alter Regulated Effects of Easily Extractable Glomalin-Related Soil Protein on Fruit Quality and Soil Properties of Satsuma Mandarin. Agronomy 2023, 13, 881. [Google Scholar] [CrossRef]
- Liu, R.-C.; Gao, W.-Q.; Srivastava, A.-K.; Zou, Y.-N.; Kuča, K.; Hashem, A.; Abd_Allah, E.-F.; Wu, Q.-S. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Basha, D.-M.-A.; Hellal, F.; El-Sayed, S. Effects of Potassium and Humic Acid on Amelioration of Soil Salinity Hazardous on Pea Plants. Asian J. Soil Sci. Plant Nutr. 2020, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rajamani, K.; Reddy, K.-I.; Srinivas, A. Influence of Super Absorbent Polymer and Humic Acid on Maize Yield and Nutrient Uptake on Rainfed Alfisols. Int. J. Environ. Clim. Chang. 2021, 11, 97–106. [Google Scholar] [CrossRef]
- Schindler, F.-V.; Mercer, E.-J.; Rice, J.-A. Chemical characteristics of glomalin-related soil protein (GRSP) extracted from soils of varying organic matter content. Soil Biol. Biochem. 2007, 39, 320–329. [Google Scholar] [CrossRef]
- Funakoshi, Y.; Daimon, H.; Matsumura, A. Morphological and physiological studies on densely branched lateral roots triggered by localized phosphate in Sesbania cannabina. J. Plant Nutr. Soil Sci. 2018, 181, 336–344. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Hao, Y.; Wu, X.-L.; Dai, F.-J.; Abd-Allah, E.-F.; Wu, Q.-S.; Liu, S.-R. Arbuscular mycorrhizal fungi improve drought tolerance of tea plants via modulating root architecture and hormones. Plant Growth Regul. 2023. [Google Scholar] [CrossRef]
- Sanjeev Krishna, S.; Nanja Reddy, Y.-A. Root growth pattern in finger millet under well-watered and drought stress condition: Its relation to shoot mass production. Plant Physiol. Rep. 2021, 26, 393–401. [Google Scholar] [CrossRef]
- Browne, M.; Yardimci, N.-T.; Scoffoni, C.; Jarrahi, M.; Sack, L. Prediction of leaf water potential and relative water content using terahertz radiation spectroscopy. Plant Direct 2020, 4, e00197. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.-M.; Turner, N.-C. Osmotic adjustment in leaves of sorghum in response to water deficits. Plant Physiol. 1978, 61, 122–126. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.-S.; James, R.-A.; Munns, R.; Condon, T.-A.; Passioura, J.-B. Osmotic adjustment leads to anomalously low estimates of relative water content in wheat and barley. Funct. Plant Biol. 2008, 35, 1172–1182. [Google Scholar] [CrossRef]
- Yang, L.; Zou, Y.-N.; Tian, Z.-H.; Wu, Q.-S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Johnston, A.-E.; Poulton, P.-R.; Fixen, P.-E.; Curtin, D. Phosphorus: Its efficient use in agriculture. Adv. Agron. 2014, 123, 177–228. [Google Scholar] [CrossRef]
- Singh, D.; Alam, A.; Jha, S.-K.; Kumar, S.; Pandey, R.; Chinnusamy, V.; Tripathi, S.; Sathee, L. Impact of single and dual deficiency of nitrogen and iron on photosynthesis and fluorescence parameters in hydroponically and field grown bread wheat. Plant Physiol. Rep. 2022, 27, 632–640. [Google Scholar] [CrossRef]
- Meng, X.; Bai, S.; Wang, S.; Pan, Y.; Chen, K.; Xie, K.; Wang, M.; Guo, S. The sensitivity of photosynthesis to magnesium deficiency differs between rice (Oryza sativa L.) and cucumber (Cucumis sativus L.). Front. Plant Sci. 2023, 14, 1164866. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Chavez, M.-C.; Carrillo-Gonzalez, R.; Wright, S.-F.; Nichols, K.-A. The role of glomalin, a protein produced by arbuscular mycorrhizal fungi, in sequestering potentially toxic elements. Environ. Pollut. 2004, 130, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Rehman, M.-Z.-U.; Maqbool, A. A critical review on the effects of zinc at toxic levels of cadmium in plants. Environ. Sci. Pollut. Res. 2019, 26, 6279–6289. [Google Scholar] [CrossRef]
- Ketehouli, T.; Quoc, V.-H.-N.; Dong, J.; Do, H.; Li, X.; Wang, F. Overview of the roles of calcium sensors in plants’ response to osmotic stress signalling. Funct. Plant Biol. 2022, 49, 589–599. [Google Scholar] [CrossRef]
- Schmidt, S.-B.; Jensen, P.-E.; Husted, S. Manganese deficiency in plants: The impact on photosystem II. Trends Plant Sci. 2016, 21, 622–632. [Google Scholar] [CrossRef]
- Chen, W.; Mou, X.; Meng, P.; Chen, J.; Tang, X.; Meng, G.; Xin, K.; Zhang, Y.; Wang, C. Effects of arbuscular mycorrhizal fungus inoculation on the growth and nitrogen metabolism of Catalpa bungei CA Mey. under different nitrogen levels. Front. Plant Sci. 2023, 14, 1138184. [Google Scholar] [CrossRef]
- Khalvati, M.-A.; Hu, Y.; Mozafar, A.; Schmidhalter, U. Quantification of water uptake by arbuscular mycorrhizal hyphae and its significance for leaf growth, water relations, and gas exchange of barley subjected to drought stress. Plant Biol. 2005, 7, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Zou, Y.-N.; Wu, Q.-S. Quantitative estimation of water uptake by mycorrhizal extraradical hyphae in citrus under drought stress. Sci. Hortic. 2018, 229, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Sade, N.; Vinocur, B.-J.; Diber, A.; Shatil, A.; Ronen, G.; Nissan, H.; Wallach, R.; Karchi, H.; Moshelion, M. Improving plant stress tolerance and yield production: Is the tonoplast aquaporin SlTIP2; 2 a key to isohydric to anisohydric conversion? New Phytol. 2009, 181, 651–661. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.-M.; Aroca, R. Modulation of Aquaporin Genes by the Arbuscular Mycorrhizal Symbiosis in Relation to Osmotic Stress Tolerance: Aquaporin in AM Plants Under Osmotic Stress. In Symbioses and Stress: Joint Ventures in Biology; Springer: Dordrecht, The Netherlands, 2010; pp. 357–374. [Google Scholar] [CrossRef]
- Šurbanovski, N.; Sargent, D.-J.; Else, M.-A.; Simpson, D.-W.; Zhang, H.; Grant, O.-M. Expression of Fragaria vesca PIP aquaporins in response to drought stress: PIP down-regulation correlates with the decline in substrate moisture content. PLoS ONE 2013, 8, e74945. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-Y.; Guo, X.-N.; Wu, X.-L.; Dai, F.-J.; Wu, Q.-S. The Comprehensive Effects of Rhizophagus intraradices and P on Root System Architecture and P Transportation in Citrus limon L. Agriculture 2022, 12, 317. [Google Scholar] [CrossRef]
- Koide, R.-T.; Peoples, M.-S. Behavior of Bradford-reactive substances is consistent with predictions for glomalin. Appl. Soil Ecol. 2013, 63, 8–14. [Google Scholar] [CrossRef]
- Bradford, M.-M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Sade, N.; Galkin, E.; Moshelion, M. Measuring Arabidopsis, tomato and barley leaf relative water content (RWC). Bio-protocol 2015, 5, e1451. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.-B. Measurement of leaf water potential by the dye method. Ecology 1967, 48, 1038–1041. [Google Scholar] [CrossRef]
- Parkinson, J.-A.; Allen, S.-E. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun. Soil Sci. Plan. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Medveckienė, B.; Kulaitienė, J.; Vaitkevičienė, N.; Levickienė, D.; Bunevičienė, K. Effect of harvesting in different ripening stages on the content of the mineral elements of rosehip (Rosa spp.) fruit flesh. Horticulturae 2022, 8, 467. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Stem Diameter (mm) | Leaf Number (#/plant) | Biomass (g FW/plant) | ||
|---|---|---|---|---|---|---|
| Leaf | Shoot | Root | ||||
| Citrate buffer | 8.42 ± 0.75 c | 0.30 ± 0.04 b | 8.60 ± 1.14 b | 0.39 ± 0.05 d | 0.38 ± 0.04 e | 1.06 ± 0.07 c |
| 1/4 EE-GRSP | 9.40 ± 0.91 b | 0.32 ± 0.02 b | 9.40 ± 1.52 b | 0.57 ± 0.05 c | 0.58 ± 0.01 d | 0.87 ± 0.09 c |
| 1/2 EE-GRSP | 13.60 ± 0.53 a | 0.39 ± 0.04 a | 13.40 ± 1.92 a | 1.22 ± 0.11 b | 1.12 ± 0.04 b | 2.24 ± 0.30 a |
| 3/4 EE-GRSP | 14.32 ± 0.73 a | 0.40 ± 0.04 a | 13.80 ± 1.67 a | 1.07 ± 0.05 b | 0.91 ± 0.01 c | 1.80 ± 0.14 b |
| Full EE-GRSP | 13.80 ± 0.57 a | 0.37 ± 0.03 a | 13.80 ± 2.59 a | 1.70 ± 0.16 a | 1.18 ± 0.05 a | 1.95 ± 0.12 ab |
| Treatment | Total Length (cm) | Projected Area (cm²) | Surface Area (cm²) | Average Diameter (mm) | Volume (cm³) | Taproot Length (cm) | Lateral Root Numbers (#/plant) | ||
|---|---|---|---|---|---|---|---|---|---|
| 1st-Order | 2nd-Order | 3rd-Order | |||||||
| Citrate Buffer | 156.6 ± 9.4 a | 11.79 ± 0.65 a | 12.20 ± 0.53 c | 0.71 ± 0.01 a | 0.73 ± 0.02 c | 9.33 ± 1.53 c | 52 ± 3 c | 54 ± 3 d | 23 ± 3 a |
| 1/4EE-GRSP | 160.3 ± 12.7 a | 10.18 ± 0.34 b | 12.49 ± 1.05 bc | 0.72 ± 0.09 a | 0.86 ± 0.09 c | 11.83 ± 1.61 b | 43 ± 3 c | 63 ± 7 d | 12 ± 3 b |
| 1/2EE-GRSP | 164.5 ± 6.6 a | 11.47 ± 0.68 ab | 13.19 ± 0.43 bc | 0.71 ± 0.05 a | 1.61 ± 0.15 a | 16.67 ± 0.58 a | 80 ± 9 ab | 223 ± 12 b | 13 ± 3 b |
| 3/4EE-GRSP | 180.7 ± 14.9 a | 12.82 ± 1.13 a | 13.97 ± 0.33 a | 0.65 ± 0.06 a | 1.37 ± 0.08 b | 18.67 ± 1.53 a | 75 ± 7 b | 155 ± 12 c | 12 ± 4 b |
| Full EE-GRSP | 168.5 ± 15.2 a | 12.34 ± 0.56 a | 13.58 ± 0.46 ab | 0.64 ± 0.03 a | 1.40 ± 0.04 b | 12.33 ± 1.15 b | 86 ± 4 a | 256 ± 13 a | 19 ± 2 a |
| Treatment | N (g/kg) | P (g/kg) | K (g/kg) | Ca (g/kg) | Mg (g/kg) | Fe (g/kg) | Mn (mg/kg) | Zn (mg/kg) |
|---|---|---|---|---|---|---|---|---|
| Leaf | ||||||||
| Citrate Buffer | 16.91 ± 0.32 a | 1.15 ± 0.09 e | 14.72 ± 1.20 a | 22.32 ± 0.53 a | 2.74 ± 0.21 a | 0.11 ± 0.01 d | 26.74 ± 1.45 a | 0.25 ± 0.02 a |
| 1/4 EE-GRSP | 17.16 ± 0.42 a | 1.96 ± 0.12 d | 15.21 ± 1.62 a | 21.95 ± 0.89 a | 2.89 ± 0.15 a | 0.14 ± 0.01 c | 19.20 ± 1.33 bc | 0.17 ± 0.00 c |
| 1/2 EE-GRSP | 8.88 ± 0.37 b | 3.94 ± 0.09 b | 16.24 ± 1.62 a | 21.76 ± 0.48 a | 2.80 ± 0.17 a | 0.14 ± 0.01 c | 21.00 ± 0.66 b | 0.19 ± 0.01 b |
| 3/4 EE-GRSP | 9.37 ± 0.19 b | 5.05 ± 0.08 a | 15.91 ± 1.48 a | 22.00 ± 1.07 a | 3.07 ± 0.23 a | 0.21 ± 0.01 a | 18.35 ± 1.13 c | 0.15 ± 0.01 cd |
| Full EE-GRSP | 9.51 ± 1.18 b | 3.62 ± 0.15 c | 16.83 ± 1.27 a | 21.70 ± 0.72 a | 2.77 ± 0.19 a | 0.19 ± 0.01 b | 18.77 ± 0.23 c | 0.14 ± 0.00 d |
| Root | ||||||||
| Citrate Buffer | 5.31 ± 0.35 a | 1.92 ± 0.06 d | 10.76 ± 0.59 a | 14.13 ± 0.19 a | 1.56 ± 0.05 c | 5.27 ± 0.25 d | 140.46 ± 1.37 a | 0.40 ± 0.02 d |
| 1/4 EE-GRSP | 5.06 ± 0.14 ab | 1.96 ± 0.25 d | 9.76 ± 0.71 a | 11.59 ± 0.42 b | 1.64 ± 0.06 c | 5.59 ± 0.04 d | 105.56 ± 5.17 b | 0.43 ± 0.02 cd |
| 1/2 EE-GRSP | 4.83 ± 0.10 b | 2.51 ± 0.02 b | 10.28 ± 0.61 a | 10.71 ± 0.42 c | 2.22 ± 0.13 ab | 6.15 ± 0.24 c | 85.27 ± 1.17 c | 0.86 ± 0.01 b |
| 3/4 EE-GRSP | 4.89 ± 0.07 b | 2.79 ± 0.01 a | 11.04 ± 0.17 a | 11.70 ± 0.40 b | 2.42 ± 0.16 a | 9.44 ± 0.50 a | 65.50 ± 0.71 d | 1.00 ± 0.10 a |
| Full EE-GRSP | 5.03 ± 0.09 ab | 2.20 ± 0.03 c | 9.79 ± 1.13 a | 10.05 ± 0.07 d | 2.13 ± 0.11 b | 7.13 ± 0.13 b | 56.58 ± 0.85 e | 0.49 ± 0.02 c |
| Genes | Gene ID | Sequence of Forward Primer (5′→3′) | Sequence of Reverse Primer (5′→3′) |
|---|---|---|---|
| ClTIP 2;1 | Cs1g15440.1 | CCTTCAAGGCCTATCTTGCTGAGTT | CCTGATGGATCAAGTGCTGCATCTG |
| ClTIP 4;1 | Cs4g19580 | TCTTCATCGTCAGGTCTCATGTTCC | ATAGTGACCCACAGATAACACCGTG |
| ClTIP 2;2 | Cs5g08710.1 | TTGAATGCCGCTGAAGGTTTGGTTA | AGCAATGGGCGCAATTGTTCCTAGT |
| ClTIP 1;3 | Cs7g28650 | GATCGTGGAGAGAATTGAAGTTGTG | GCAAGACATAAATCCATCCACTCCT |
| ClTIP 1;2 | Cs8g17900 | TAAAAAATGCCCGGAATTGCTATCG | TCCATTGTCCGTGAGCTTGCTGTAA |
| ClTIP 5;1 | Cs9g14770 | AGTCCAGACGCAGCATCAAATACAT | ACGGTGATGTGTCCACTCACTGCCT |
| ClTIP 1;1 | Orange1.1t03005.1 | TCGCTAATCCACTGCTAACCAACTT | TGGTCTCCACTTCTCAACTAGGAGC |
| β-Actin | Cs1g05000 | CCGACCGTATGAGCAAGGAAA | TTCCTGTGGACAATGGATGGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.-N.; Hao, Y.; Wu, X.-L.; Chen, X.; Liu, C.-Y. Exogenous Easily Extractable Glomalin-Related Soil Protein Stimulates Plant Growth by Regulating Tonoplast Intrinsic Protein Expression in Lemon. Plants 2023, 12, 2955. https://doi.org/10.3390/plants12162955
Guo X-N, Hao Y, Wu X-L, Chen X, Liu C-Y. Exogenous Easily Extractable Glomalin-Related Soil Protein Stimulates Plant Growth by Regulating Tonoplast Intrinsic Protein Expression in Lemon. Plants. 2023; 12(16):2955. https://doi.org/10.3390/plants12162955
Chicago/Turabian StyleGuo, Xiao-Niu, Yong Hao, Xiao-Long Wu, Xin Chen, and Chun-Yan Liu. 2023. "Exogenous Easily Extractable Glomalin-Related Soil Protein Stimulates Plant Growth by Regulating Tonoplast Intrinsic Protein Expression in Lemon" Plants 12, no. 16: 2955. https://doi.org/10.3390/plants12162955