

Salt Tolerance Evaluation of Cucumber Germplasm under Sodium Chloride Stress
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of NaCl Stress on Plant Growth Phenotype of Various Cucumber Seedlings
2.2. Effects of NaCl Stress on Salt Injury Index and Survival Rate of Various Cucumber Seedlings
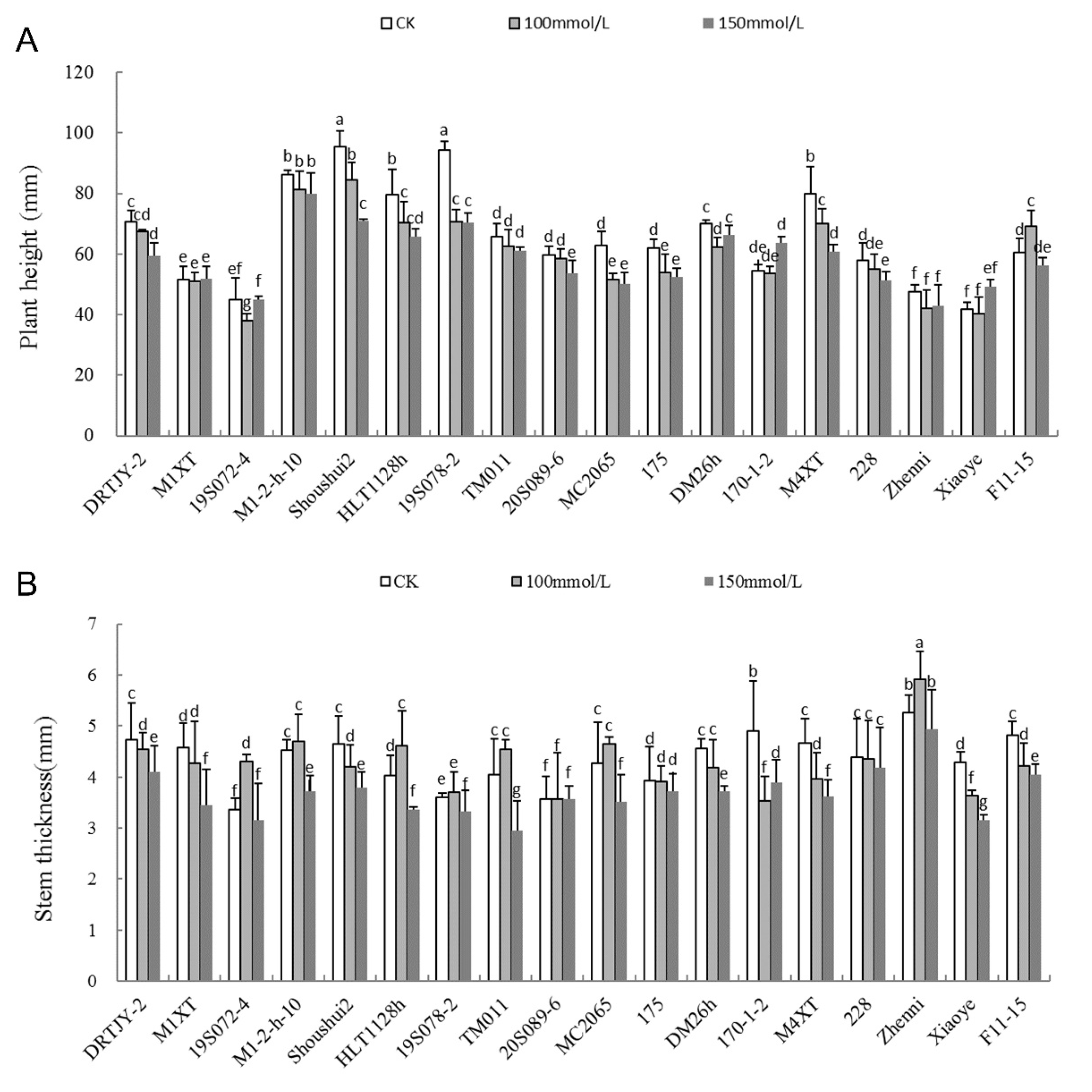
2.3. Effects of NaCl Stress on Plant Height and Stem Diameter
2.4. Effects of NaCl Stress on Chlorophyll Content
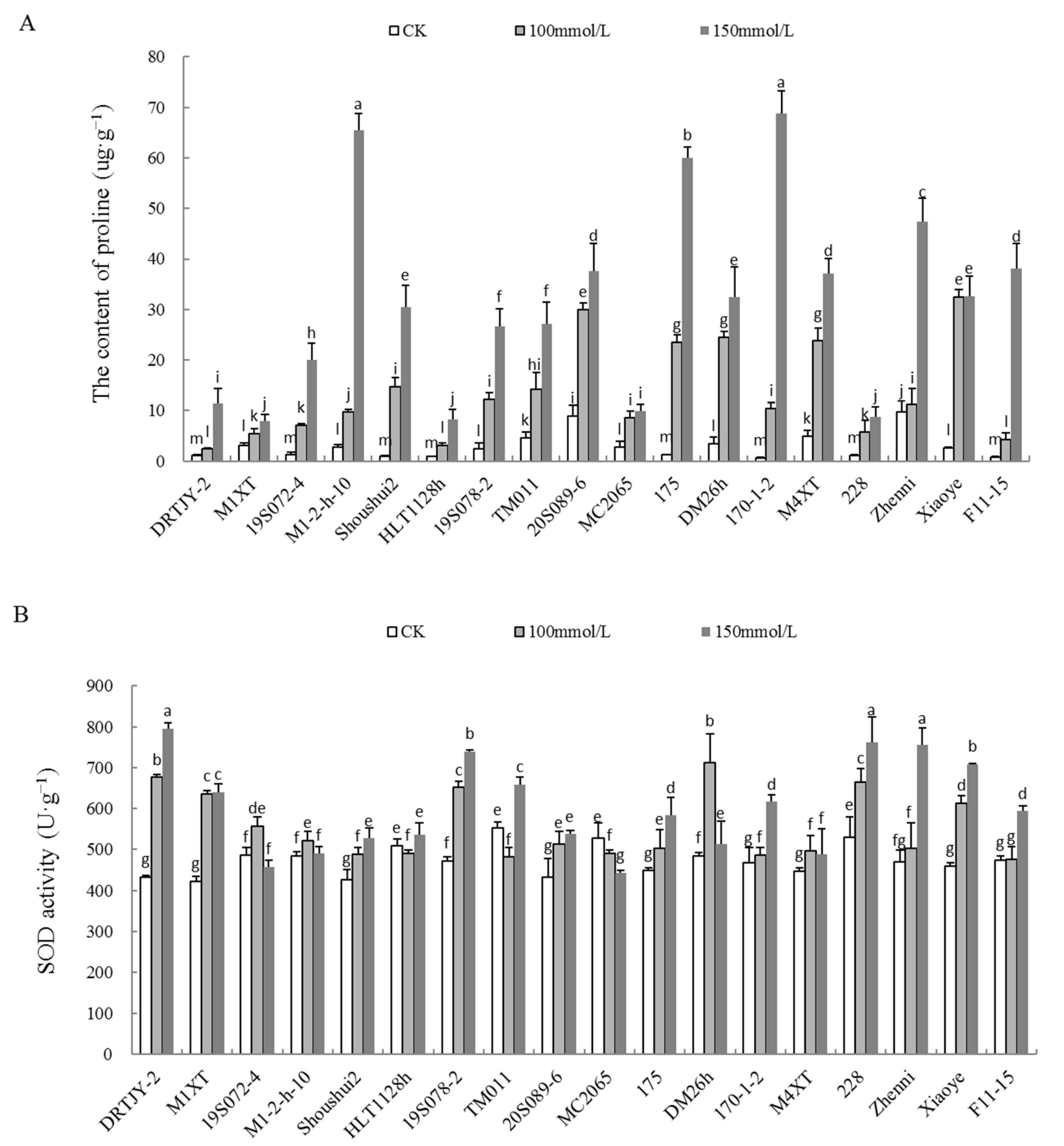
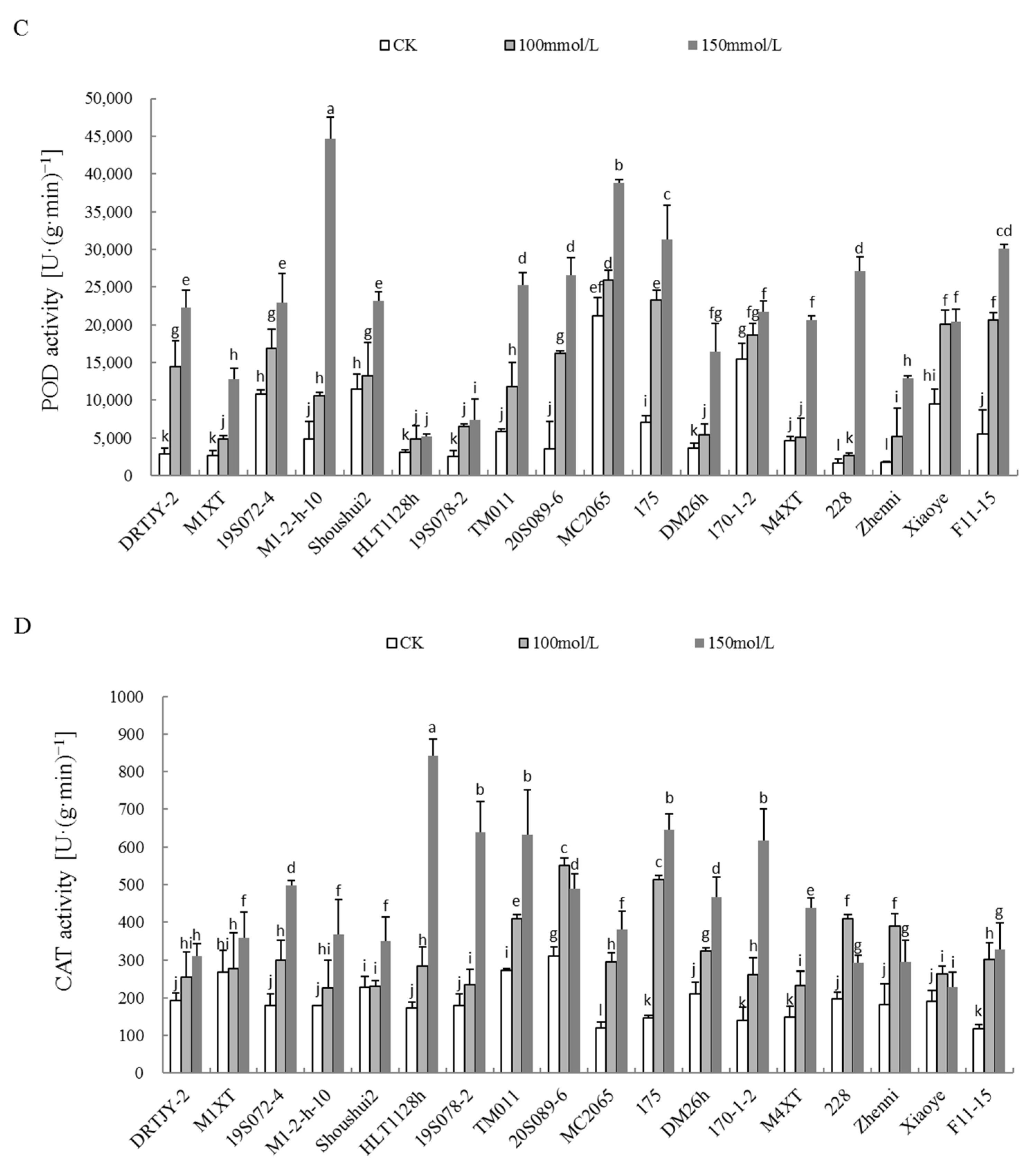
2.5. Effects of NaCl Stress on the Proline Content and POD, SOD, and CAT Activities
2.6. The Effect of NaCl Stress on the MDA Content of Various Cucumber Seedlings
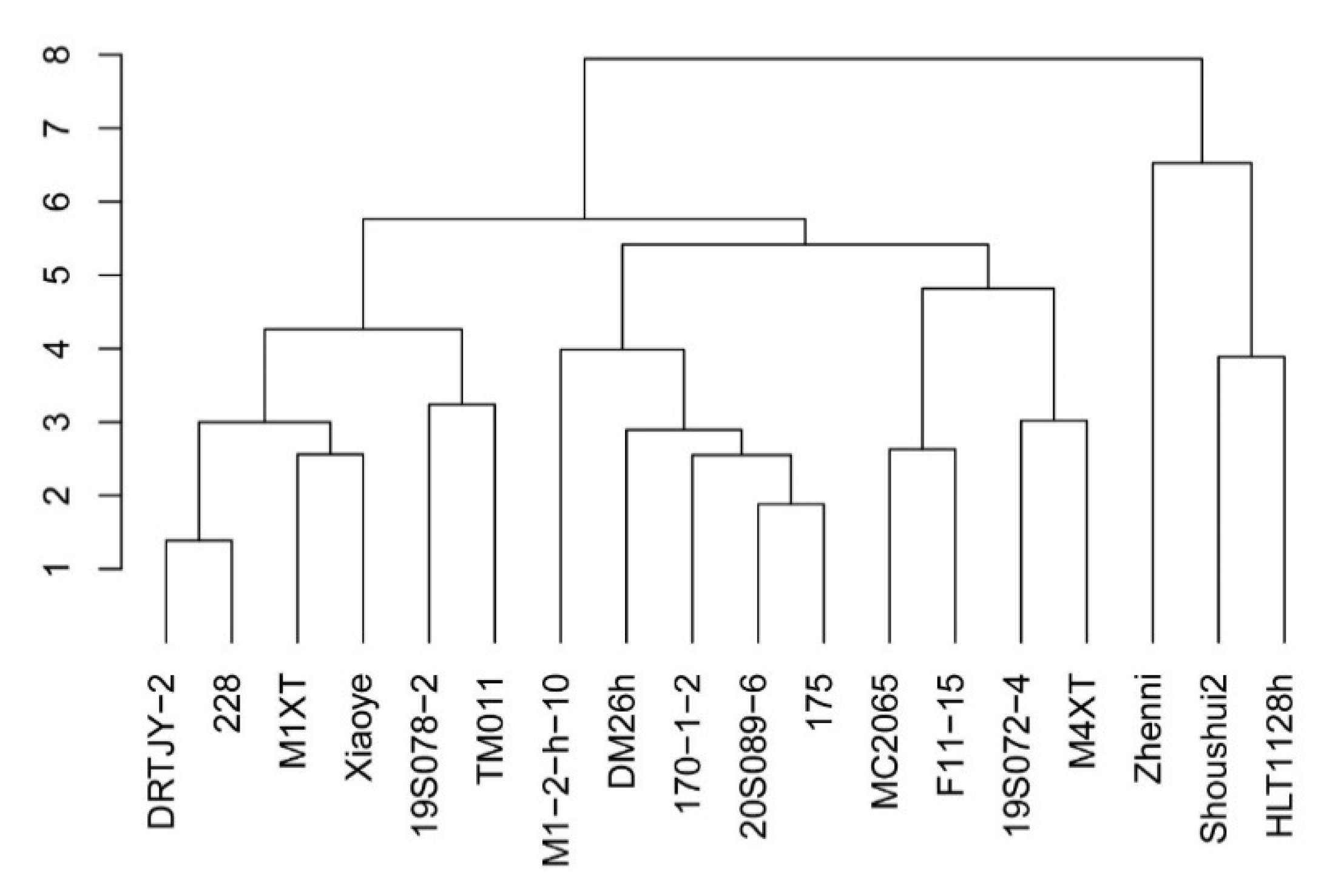
2.7. Cluster Analysis of Salt Tolerance of Cucumber
2.8. Principal Component Analysis and Comprehensive Evaluation of Cucumber Salt Tolerance at the Seedling Stage
3. Materials and Methods
3.1. Plant Materials
3.2. NaCl Treatment on Cucumber Seedlings
3.3. Classification of the Salt-Tolerance Level of Cucumbers
3.4. Determination of Growth Index
3.5. Quantification of Total Chlorophyll
3.6. Determination of Proline Content
3.7. Extraction and Assay of Antioxidant Enzymes
3.8. Determination of MDA Content
3.9. Data Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alfosea-Simón, M.; Zavala-Gonzalez, E.A.; Camara-Zapata, J.M.; Martínez-Nicolás, J.J.; Simón, I.; Simón-Grao, S.; García-Sánchez, F. Effect of foliar application of amino acids on the salinity tolerance of tomato plants cultivated under hydroponic system. Sci. Hortic. 2020, 272, 109509. [Google Scholar] [CrossRef]
- El-Mogy, M.M.; Garchery, C.; Stevens, R. Irrigation with salt water affects growth, yield, fruit quality, storability and marker-gene expression in cherry tomato. Acta Agric. Scand. B Soil Plant Sci. 2018, 68, 727–737. [Google Scholar] [CrossRef]
- Gul, H.; Ali, R.; Rauf, M.; Hamayun, M.; Arif, M.; Khan, S.A.; Parveen, Z.; Alrefaei, A.F.; Lee, I.J. Aspergillus welwitschiae BK isolate ameliorates the physicochemical characteristics and mineral profile of maize under salt stress. Plants 2023, 12, 1703. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Song, L.; Shu, Y.; Wang, S.; Niu, J.; Wang, Z.; Yu, T.; Gu, W.; Ma, H. Comparative proteomic analysis of seedling leaves of different salt tolerant soybean genotypes. J. Proteom. 2012, 75, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Li, L.; Yang, Z.; Wang, Y.; Li, J.; Chen, W.; Wang, J.; Meng, Z. Transcriptome analysis of genes in response to magnesium nitrate stress on cucumber leaf. Sci. Hortic. 2021, 288, 110391. [Google Scholar] [CrossRef]
- Wang, M.; Yang, S.; Sun, L.; Feng, Z.; Gao, Y.; Zhai, X.; Dong, Y.; Wu, H.; Cui, Y.; Li, S.; et al. A CBL4-CIPK6 module confers salt tolerance in cucumber. Veg. Res. 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Jia Danli, Y.Z.; Junling, G.; Caixia, G.; Yongliang, W. Salt tolerance screening of six maize varieties. Chin. Agric. Sci. Bull. 2017, 33, 1–8. [Google Scholar]
- Atieno, J.; Colmer, T.D.; Taylor, J.; Li, Y.; Quealy, J.; Kotula, L.; Nicol, D.; Nguyen, D.T.; Brien, C.; Langridge, P.; et al. Novel salinity tolerance loci in chickpea identified in glasshouse and field environments. Front. Plant Sci. 2021, 12, 667910. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, P.; Kumar, A.; Kyriacou, M.; Colla, G.; Rouphael, Y. Grafting tomato as a tool to improve salt tolerance. Agronomy 2020, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Fahad, S.; Zamin, M.; Shah, S.; Mian, I.A.; Danish, S.; Zafar-Ul-Hye, M.; Battaglia, M.L.; Naz, R.M.M.; Saeed, B.; et al. Coupling phosphate-solubilizing bacteria with phosphorus supplements improve maize phosphorus acquisition and growth under Lime induced salinity stress. Plants 2020, 9, 900. [Google Scholar] [CrossRef]
- Dinneny, J.R. Traversing organizational scales in plant salt-stress responses. Curr. Opin. Plant Biol. 2015, 23, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-s.; Zhang, J.; Li, M.-x.; Shi, L.-x. Metabolomics analysis reveals the salt-tolerant mechanism in glycine soja. J. Plant Growth Regul. 2017, 36, 460–471. [Google Scholar] [CrossRef]
- Luo, H.; Zhou, Z.; Song, G.; Yao, H.; Han, L. Antioxidant enzyme activity and microRNA are associated with growth of Poa pratensis callus under salt stress. Plant Biotechnol. Rep. 2020, 14, 429–438. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, W.; Su, C.; Ma, H.; Pan, Y.; Zhang, X.; Li, J. Tandem 13-Lipoxygenase genes in a cluster confers yellow-green leaf in cucumber. Int. J. Mol. Sci. 2019, 20, 3102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Jiang, X.; Zhang, J.; He, Y.; Zhu, X.; Zhou, X.; Gong, H.; Yin, J.; Liu, Y. Silicon confers cucumber resistance to salinity stress through regulation of proline and cytokinins. Plant Physiol. Biochem. 2020, 156, 209–220. [Google Scholar] [CrossRef]
- Xu, Y.; Yuan, Y.H.; Du, N.S.; Wang, Y.; Shu, S.; Sun, J.; Guo, S.R. Proteomic analysis of heat stress resistance of cucumber leaves when grafted onto Momordica rootstock. Hortic. Res. 2018, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.P.; Li, H.; Jiang, C.D.; Tang, Y.D.; Lei, S. Ion micro-distribution in varying aged leaves in salt-treated cucumber seedlings. Plant Physiol. Biochem. 2018, 129, 71–76. [Google Scholar] [CrossRef]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Cao, Q.; Du, L.; Yang, Z.; Li, L.; Duan, X.; Yang, W.; Chen, W.; Meng, Z. Screening and evaluation of cucumber salt-tolerant germplasm. J. Nuclear Agric. Sci. 2022, 36, 865–875. [Google Scholar]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Kabała, K.; Reda, M.; Wdowikowska, A.; Janicka, M. Role of plasma membrane NADPH oxidase in response to salt stress in cucumber seedlings. Antioxidants 2022, 11, 1534. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.A.-O.; Liu, L.A.-O.; Rupasinghe, T.W.T.; Roessner, U.A.-O.; Barkla, B.A.-O. Salt stress alters membrane lipid content and lipid biosynthesis pathways in the plasma membrane and tonoplast. Plant Physiol. 2022, 189, 805–826. [Google Scholar] [CrossRef] [PubMed]
- Bin, J.; Zhu, M.; Ding, H.; Zai, Z.; Shi, T.; Wang, L.; Yang, X.; Yue, Y. New insights into the roles of osmanthus fragrans heat-shock transcription factors in cold and other stress responses. Horticulturae 2022, 8, 80. [Google Scholar] [CrossRef]
- Prisco, J.T.; Gomes-Filho, E.; Miranda, R. Physiology and biochemistry of plants growing under salt stress. In Manejo da Salinidade na Agricultura: Estudos Básicos e Aplicados, 2nd ed.; Hans Raj Gheyi, H.R., Dias, N.d.S., de Lacerda, C.F., Filho, E.G., Eds.; Expressão Gráfica e Editora: Fortaleza, Brazil, 2016; Chapter 12; pp. 163–180. [Google Scholar]
- Dash, M.; Panda, S. Salt Stress Induced Changes in Growth and Enzyme Activities in Germinating Phaseolus Mungo Seeds. Biol. Plant. 2001, 44, 587–589. [Google Scholar] [CrossRef]
- Benzarti, M.; Rejeb, K.B.; Messedi, D.; Mna, A.B.; Hessini, K.; Ksontini, M.; Abdelly, C.; Debez, A. Effect of high salinity on Atriplex portulacoides: Growth, leaf water relations and solute accumulation in relation with osmotic adjustment. S. Afr. J. Bot. 2014, 95, 70–77. [Google Scholar] [CrossRef]
- Yousfi, S.; Rabhi, M.; Hessini, K.; Abdelly, C.; Gharsalli, M. Differences in efficient metabolite management and nutrient metabolic regulation between wild and cultivated barley grown at high salinity. Plant Biol. 2010, 12, 650–658. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 45, 1096–1109. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G.; Zhang, H.; Liu, B.; Yang, Y.; Qin, L.; Chen, E.; Guan, Y. Identification of QTLs for salt tolerance at germination and seedling stage of Sorghum bicolor L. Moench. Euphytica 2014, 196, 117–127. [Google Scholar] [CrossRef]
- Guo, Y.; Jiang, Q.; Hu, Z.; Sun, X.; Fan, S.; Zhang, H. Function of the auxin-responsive gene TaSAUR75 under salt and drought stress. Crop J. 2018, 6, 181–190. [Google Scholar] [CrossRef]
- Esfandiari Ghalati, R.; Shamili, M.; Homaei, A. Effect of putrescine on biochemical and physiological characteristics of guava (Psidium guajava L.) seedlings under salt stress. Sci. Hortic. 2020, 261, 108961. [Google Scholar] [CrossRef]
- Krishnaraj, C.; Jagan, E.G.; Ramachandran, R.; Abirami, S.M.; Mohan, N.; Kalaichelvan, P.T. Effect of biologically synthesized silver nanoparticles on Bacopa monnieri (Linn.) Wettst. plant growth metabolism. Process Biochem. 2012, 47, 651–658. [Google Scholar] [CrossRef]
- Naliwajski, M.R.; Skłodowska, M. The oxidative stress and antioxidant systems in cucumber cells during acclimation to salinity. Biol. Plant. 2014, 58, 47–54. [Google Scholar] [CrossRef]
- Yan, S.; Chong, P.; Zhao, M. Effect of salt stress on the photosynthetic characteristics and endogenous hormones, and: A comprehensive evaluation of salt tolerance in Reaumuria soongorica seedlings. Plant Signal. Behav. 2022, 17, 2031782. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, Y.; Wang, W.; Xu, K.; Niu, L.; Yu, L.; Zou, J.W.; Wang, Z.; Zhang, Y.J.; Wang, F.; et al. Evaluation of salt tolerance mechanism and study on salt tolerance relationship of different salt-tolerant wheat varieties. Comput. Intel. Neurosc. 2022, 2022, 2395568. [Google Scholar] [CrossRef]
- Zhang, R.; Hussain, S.; Wang, Y.; Liu, Y.; Li, Q.; Chen, Y.; Wei, H.; Gao, P.; Dai, Q. Comprehensive evaluation of salt tolerance in rice (Oryza sativa L.) germplasm at the germination stage. Agronomy 2021, 11, 1569. [Google Scholar] [CrossRef]
- Tang, H.; Xu, C.; Jiang, Y.; Wang, J.; Wang, Z.; Tian, L. Evaluation of physical characteristics of typical maize seeds in a cold area of North China based on principal component analysis. Processes 2021, 9, 1167. [Google Scholar] [CrossRef]
- Zheng, P.; Zhang, M.; Wang, Z.; Wang, T.; Tang, L.; Ma, E.; Liu, J.; Shi, F. Comprehensive evaluation of the fruit quality of the main cultivars of pear (Pyrus spp.) in North China. Erwerbs-Obstbau 2022, 64, 219–227. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, J.; Lu, Z.; Zhao, X.; Liu, H.; Zhao, K.; Cong, H. Salt tolerance evaluation of soybean germplasm. Xinjiang Agric. Sci. 2021, 58, 65–71. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 258–260. [Google Scholar]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Abreu, C.E.B.d.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.Z.; Peng, T.; Huang, X.S.; Fan, Q.J.; Liu, J.H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Havir, E.A.; McHale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, L.; Heath, L.P. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar]
- Wang, J.X.; Gong, H.Y.; Tu, X.J.; Guo, Z.X.; Zhao, J.N.; Shen, J.; Li, Z.Y.; Sun, J. Screening of phosphite-tolerant alfalfa varieties and identification of phosphite tolerance indicators. Acta Prataculturae Sin. 2021, 30, 186–199. [Google Scholar]
- Sithtisarn, S.; Theerawitaya, C.; Samphumphuang, T.; Takabe, T.; Singh, H.P.; Cha-um, S. Molecular and physiological responses of Egeria densa under salt stress. Flora 2023, 300, 152226. [Google Scholar] [CrossRef]
- Chatterjee, P.; Samaddar, S.; Anandham, R.; Kang, Y.; Kim, K.; Selvakumar, G.; Sa, T. Beneficial soil bacterium pseudomonas frederiksbergensis OS261 augments salt tolerance and promotes red pepper plant growth. Front. Plant Sci. 2017, 8, 705. [Google Scholar] [CrossRef] [PubMed]
- Hütsch, B.W.; He, W.; Schubert, S. Nitrogen nutritional status of young maize plants (Zea mays) is not limited by NaCl stress. J. Plant Nutr. Soil Sci. 2016, 179, 775–783. [Google Scholar] [CrossRef]
- Wang, Q.; Men, L.; Gao, L.; Tian, Y. Effect of grafting and gypsum application on cucumber (Cucumis sativus L.) growth under saline water irrigation. Agric. Water Manag. 2017, 188, 79–90. [Google Scholar] [CrossRef]
- Zhu, Y.; Gu, W.; Tian, R.; Li, C.; Ji, Y.; Li, T.; Wei, C.; Chen, Z. Morphological, physiological, and secondary metabolic responses of Taraxacum officinale to salt stress. Plant Physiol. Biochem. 2022, 189, 71–82. [Google Scholar] [CrossRef]
- Li, D.; Si, J.; Ren, X.; Li, J. Coordination in functional traits of Populus euphratica leaves under salt stress. Front. For. Glob. Change 2023, 6, 1144079. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2014, 75, 391–404. [Google Scholar] [CrossRef]
- Yu, X.; Shi, P.; Hui, C.; Miao, L.; Liu, C.; Zhang, Q.; Feng, C. Effects of salt stress on the leaf shape and scaling of Pyrus betulifolia bunge. Symmetry 2019, 11, 991. [Google Scholar] [CrossRef] [Green Version]
- Ellenberger, J.; Siefen, N.; Krefting, P.; Schulze Lutum, J.-B.; Pfarr, D.; Remmel, M.; Schröder, L.; Röhlen-Schmittgen, S. Effect of UV radiation and salt stress on the accumulation of economically relevant secondary metabolites in bell pepper plants. Agronomy 2020, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Niu, M.; Xie, J.; Sun, J.; Huang, Y.; Kong, Q.; Nawaz, M.A.; Bie, Z. A shoot based Na+ tolerance mechanism observed in pumpkin—An important consideration for screening salt tolerant rootstocks. Sci. Hortic. 2017, 218, 38–47. [Google Scholar] [CrossRef]
- Ren, L.; Li, F.; Jiang, J.; Li, H. BpTCP3 transcription factor improves salt tolerance of Betula platyphylla by reducing reactive oxygen species damage. Forests 2021, 12, 1633. [Google Scholar] [CrossRef]
- Yasar, F.; Ellialtioglu, S.; Yildiz, K. Effect of salt stress on antioxidant defense systems, lipid peroxidation, and chlorophyll content in green bean. Russ. J. Plant Physiol. 2008, 55, 782–786. [Google Scholar] [CrossRef]
- Pietrzykowska, M.; Suorsa, M.; Semchonok, D.A.; Tikkanen, M.; Boekema, E.J.; Aro, E.-M.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Lu, T.; Zhang, G.; Qi, M.; Tang, W.; Li, L.; Liu, Y.; Li, T. Photosystem inhibition and protection in tomato leaves under low light. Sci. Hortic. 2017, 217, 145–155. [Google Scholar] [CrossRef]
- Sevengor, S.; Yasar, F.; Kusvuran, S.; Ellialtioglu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Raza, M.A.; Rizwan, M.; Tariq, R.; Ali, S.; Huang, L. Silver nanoparticles improved the plant growth and reduced the sodium and chlorine accumulation in pearl millet: A life cycle study. Environ. Sci. Pollut. Res. 2021, 28, 13712–13724. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; Ganie, S.A.; Hossain, M.S.; Gomaa, A.M. Seed priming with cypress leaf extract enhances photosynthesis and antioxidative defense in zucchini seedlings under salt stress. Sci. Hortic. 2022, 293, 110707. [Google Scholar] [CrossRef]
- Sehar, Z.; Gautam, H.; Masood, A.; Khan, N.A. Ethylene- and proline-dependent regulation of antioxidant enzymes to mitigate heat stress and boost photosynthetic efficacy in wheat plants. J. Plant Growth Regul. 2023, 42, 2683–2697. [Google Scholar] [CrossRef]
- Kamali-Andani, N.; Fallah, S.; Peralta-Videa, J.R.; Golkar, P. A comprehensive study of selenium and cerium oxide nanoparticles on mung bean: Individual and synergistic effect on photosynthesis pigments, antioxidants, and dry matter accumulation. Sci. Total Environ. 2022, 830, 154837. [Google Scholar] [CrossRef]
- Avestan, S.; Ghasemnezhad, M.; Esfahani, M.; Byrt, C.S. Application of nano-silicon dioxide improves salt stress tolerance in strawberry plants. Agronomy 2019, 9, 246. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Hou, L.; Xiao, P.; Guo, Y.; Deyholos, M.K.; Liu, X. VvWRKY30, a grape WRKY transcription factor, plays a positive regulatory role under salinity stress. Plant Sci. 2019, 280, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhou, K.; Liu, Y.; Yang, S.; Zhang, J.; Gong, X.; Ma, F. Overexpression of MdMIPS1 enhances salt tolerance by improving osmosis, ion balance, and antioxidant activity in transgenic apple. Plant Sci. 2020, 301, 110654. [Google Scholar] [CrossRef]
- Zaidi, I.; Ebel, C.; Belgaroui, N.; Ghorbel, M.; Amara, I.; Hanin, M. The wheat MAP kinase phosphatase 1 alleviates salt stress and increases antioxidant activities in Arabidopsis. J. Plant Physiol. 2016, 193, 12–21. [Google Scholar] [CrossRef]
- Sheikh-Mohamadi, M.-H.; Etemadi, N.; Nikbakht, A.; Arab, M.; Majidi, M.; Pessarakli, M. Antioxidant defence system and physiological responses of Iranian crested wheatgrass (Agropyron cristatum L.) to drought and salinity stress. Acta Physiol. Plant. 2017, 39, 245. [Google Scholar] [CrossRef]
- Farsaraei, S.; Mehdizadeh, L.; Moghaddam, M. Seed priming with putrescine alleviated salinity stress during germination and seedling growth of medicinal pumpkin. J. Soil Sci. Plant Nutr. 2021, 21, 1782–1792. [Google Scholar] [CrossRef]
- Xue, L.; Ma, J.; Hu, Q.; Ma, J. Superoxide dismutase plays an important role in maize resistance to soil CO2 stress. Acta Geol. Sin. Engl. 2023, 97, 995–1001. [Google Scholar] [CrossRef]
- Samadi, S.; Habibi, G.; Vaziri, A. Exogenous trehalose alleviates the inhibitory effects of salt stress in strawberry plants. Acta Physiol. Plant. 2019, 41, 112. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, D.; Wang, L.; Xue, Z.; Guo, M.; Duan, L.; Zheng, Y. Comparison and discrimination of Artemisia argyi and Artemisia lavandulifolia by gas chromatography-mass spectrometry-based metabolomic approach. J. AOAC Int. 2019, 102, 1814–1821. [Google Scholar] [CrossRef]
- Wang, J.; Liao, X.; Zheng, P.; Xue, S.; Peng, R. Classification of Chinese herbal medicine by laser-induced breakdown spectroscopy with principal component analysis and artificial neural network. Anal. Lett. 2018, 51, 575–586. [Google Scholar] [CrossRef]
- Chen, S.; Shen, J.; Fan, K.; Qian, W.; Gu, H.; Li, Y.; Zhang, J.; Han, X.; Wang, Y.; Ding, Z. Hyperspectral machine-learning model for screening tea germplasm resources with drought tolerance. Front. Plant Sci. 2022, 13, 1048442. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Kaur, N.; Sharma, A.; Kumar, A. Evaluation and screening of elite wheat germplasm for salinity stress at the seedling phase. Physiol. Plant. 2021, 173, 237593616. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germplasm | Salt Treatment Concentration/(mmol·L−1) | Salt Damage Index/(%) | Survival Rate/(%) |
|---|---|---|---|
| JY-2 | 100 | 10.00 | |
| 150 | 40.00 | 27.27 | |
| M1XT | 100 | 0.00 | |
| 150 | 35.00 | 20.00 | |
| 19S072-4 | 100 | 25.00 | |
| 150 | 70.00 | 9.09 | |
| M1-2-h-10 | 100 | 5.00 | |
| 150 | 50.00 | 0.00 | |
| Shoushui2 | 100 | 0.00 | |
| 150 | 15.00 | 100.00 | |
| HLT1128h | 100 | 5.00 | |
| 150 | 15.00 | 100.00 | |
| 19S078-2 | 100 | 25.00 | |
| 150 | 35.00 | 23.08 | |
| TM011 | 100 | 5.00 | |
| 150 | 70.00 | 16.67 | |
| 20S098-6 | 100 | 10.00 | |
| 150 | 45.00 | 11.11 | |
| MC2065 | 100 | 10.00 | |
| 150 | 20.00 | 70.00 | |
| 175 | 100 | 5.00 | |
| 150 | 40.00 | 0.00 | |
| DM26h | 100 | 0.00 | |
| 150 | 30.00 | 8.33 | |
| 170-1-2 | 100 | 5.00 | |
| 150 | 35.00 | 0.00 | |
| M4XT | 100 | 5.00 | |
| 150 | 50.00 | 11.11 | |
| 228 | 100 | 15.00 | |
| 150 | 40.00 | 22.22 | |
| Zhenni | 100 | 15.00 | |
| 150 | 50.00 | 77.78 | |
| Xiaoye | 100 | 5.00 | |
| 150 | 55.00 | 12.50 | |
| F11-15 | 100 | 0.00 | |
| 150 | 20.00 | 66.67 |
| Main Ingredient | Eigenvalues | Variance Contribution Rate/(%) | Cumulative Contribution Rate/(%) |
|---|---|---|---|
| 1 | 2.939 | 29.386 | 29.386 |
| 2 | 1.985 | 19.852 | 49.237 |
| 3 | 1.537 | 15.372 | 64.609 |
| 4 | 1.121 | 11.211 | 75.820 |
| 5 | 1.071 | 10.706 | 86.526 |
| 6 | 0.562 | 5.623 | 92.150 |
| 7 | 0.320 | 3.196 | 95.345 |
| 8 | 0.222 | 2.218 | 97.564 |
| 9 | 0.174 | 1.741 | 99.304 |
| 10 | 0.070 | 0.696 | 100.000 |
| Germplasm | Overall Ratings | Sort |
|---|---|---|
| DRTJY-2 | 0.22285 | 2 |
| M1XT | −0.25308 | 15 |
| 19S072-4 | −0.35987 | 17 |
| M1-2-h-10 | 0.03461 | 9 |
| Shoushui2 | −0.04529 | 12 |
| HLT1128h | −0.36740 | 18 |
| 19S078-2 | 0.09732 | 6 |
| TM011 | −0.03740 | 11 |
| 20S089-6 | −0.00371 | 10 |
| MC2065 | −0.33354 | 16 |
| 175 | 0.03754 | 8 |
| DM26h | −0.08068 | 13 |
| 170-1-2 | 0.18252 | 4 |
| M4XT | −0.16228 | 14 |
| 228 | 0.19509 | 3 |
| Zhenni | 0.67673 | 1 |
| Xiaoye | 0.09631 | 7 |
| F11-15 | 0.10027 | 5 |
| Serial Number | Germplasm Name | Ecotype |
|---|---|---|
| 1 | DRTJY-2 | North China type |
| 2 | M1XT | North China type |
| 3 | 19S072-4 | North China type |
| 4 | M1-2-h-10 | South China type |
| 5 | Shoushui2 | Eurasian type |
| 6 | HLT1128h | Eurasian type |
| 7 | 19S078-2 | North China type |
| 8 | TM011 | Japanese Long |
| 9 | 20S098-6 | North China type |
| 10 | MC2065 | North China type |
| 11 | 175 | North China type |
| 12 | DM26h | North China type |
| 13 | 170-1-2 | North China type |
| 14 | M4XT | North China type |
| 15 | 228 | North China type |
| 16 | Zhenni | Eurasian type |
| 17 | Xiaoye | Eurasian type |
| 18 | F11-15 | North China type |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Du, L.; Cao, Q.; Yang, Z.; Liu, Y.; Yang, H.; Duan, X.; Meng, Z. Salt Tolerance Evaluation of Cucumber Germplasm under Sodium Chloride Stress. Plants 2023, 12, 2927. https://doi.org/10.3390/plants12162927
Li L, Du L, Cao Q, Yang Z, Liu Y, Yang H, Duan X, Meng Z. Salt Tolerance Evaluation of Cucumber Germplasm under Sodium Chloride Stress. Plants. 2023; 12(16):2927. https://doi.org/10.3390/plants12162927
Chicago/Turabian StyleLi, Libin, Lianda Du, Qiwei Cao, Zonghui Yang, Yihan Liu, Hua Yang, Xi Duan, and Zhaojuan Meng. 2023. "Salt Tolerance Evaluation of Cucumber Germplasm under Sodium Chloride Stress" Plants 12, no. 16: 2927. https://doi.org/10.3390/plants12162927