
Unveiling the Role of Sorghum RPAP3 in the Function of R2TP Complex: Insights into Protein Assembly in Plants
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Bioinformatic Analysis of S. bicolor RPAP3
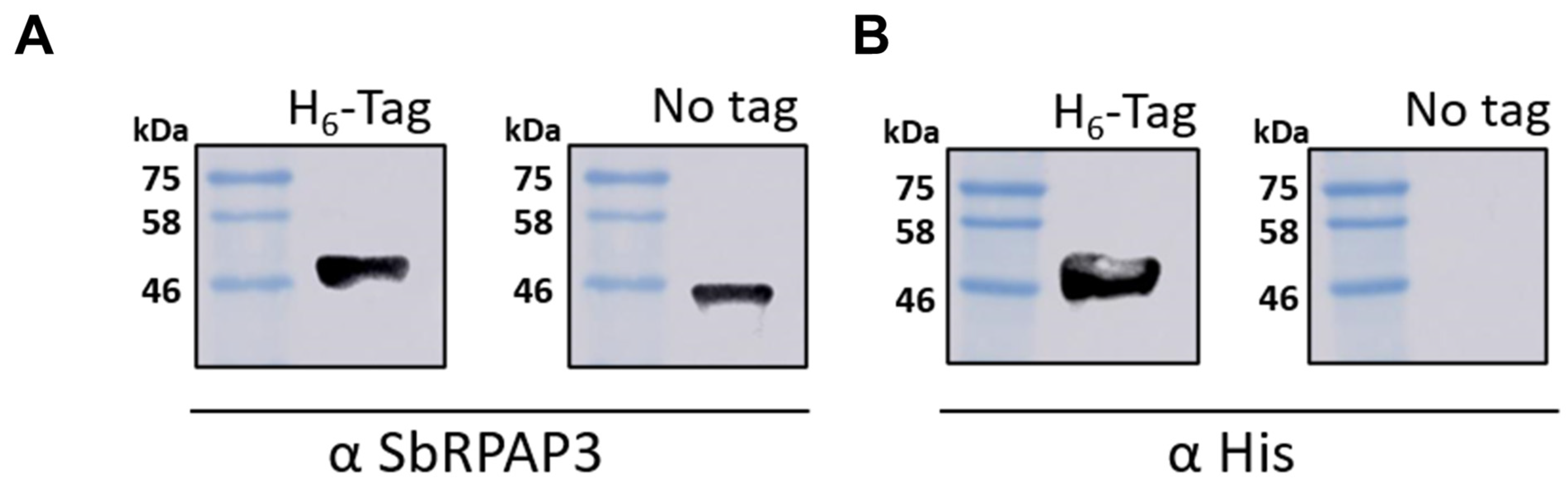
2.2. Recombinant SbRPAP3 Was Produced Pure and Folded as an Elongated Monomer in Solution
2.3. SbRPAP3 Interacts with SsHsp90, SsHsp70, and HsRUVBL1/2
3. Materials and Methods
3.1. Cloning, Expression, and Purification
3.2. Secondary Structure in Silico and Domain Prediction
3.3. Homology Model of SbRPAP3
3.4. Molecular modeling
3.5. Spectroscopy
3.6. Size Exclusion Chromatography Coupled to Multi-Angle and Quasi-Elastic Light Scattering (SEC-MALS-QELS)
3.7. Protein–Protein Interaction Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, R.; Davey, M.; Hsu, Y.-C.; Kaplanek, P.; Tong, A.; Parsons, A.B.; Krogan, N.; Cagney, G.; Mai, D.; Greenblatt, J.; et al. Navigating the Chaperone Network: An Integrative Map of Physical and Genetic Interactions Mediated by the Hsp90 Chaperone. Cell 2005, 120, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houry, W.A.; Bertrand, E.; Coulombe, B. The PAQosome, an R2TP-Based Chaperone for Quaternary Structure Formation. Trends Biochem. Sci. 2018, 43, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Marmier-Gourrier, N.; Pradet-Balade, B.; Wurth, L.; Verheggen, C.; Jády, B.E.; Rothé, B.; Pescia, C.; Robert, M.-C.; Kiss, T.; et al. The Hsp90 Chaperone Controls the Biogenesis of L7Ae RNPs through Conserved Machinery. J. Cell Biol. 2008, 180, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Pradet-Balade, B.; Verheggen, C.; Molle, D.; Boireau, S.; Georgieva, M.; Azzag, K.; Robert, M.-C.; Ahmad, Y.; Neel, H.; et al. HSP90 and Its R2TP/Prefoldin-like Cochaperone Are Involved in the Cytoplasmic Assembly of RNA Polymerase II. Mol. Cell 2010, 39, 912–924. [Google Scholar] [CrossRef] [Green Version]
- Hořejší, Z.; Stach, L.; Flower, T.G.; Joshi, D.; Flynn, H.; Skehel, J.M.; O’Reilly, N.J.; Ogrodowicz, R.W.; Smerdon, S.J.; Boulton, S.J. Phosphorylation-Dependent PIH1D1 Interactions Define Substrate Specificity of the R2TP Cochaperone Complex. Cell Rep. 2014, 7, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Cheung, K.L.Y.; Huen, J.; Houry, W.A.; Ortega, J. Comparison of the Multiple Oligomeric Structures Observed for the Rvb1 and Rvb2 Proteins. Biochem. Cell Biol. 2010, 88, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Henri, J.; Chagot, M.-E.; Bourguet, M.; Abel, Y.; Terral, G.; Maurizy, C.; Aigueperse, C.; Georgescauld, F.; Vandermoere, F.; Saint-Fort, R.; et al. Deep Structural Analysis of RPAP3 and PIH1D1, Two Components of the HSP90 Co-Chaperone R2TP Complex. Structure 2018, 26, 1196–1209.e8. [Google Scholar] [CrossRef] [Green Version]
- Martino, F.; Pal, M.; Muñoz-Hernández, H.; Rodríguez, C.F.; Núñez-Ramírez, R.; Gil-Carton, D.; Degliesposti, G.; Skehel, J.M.; Roe, S.M.; Prodromou, C.; et al. RPAP3 Provides a Flexible Scaffold for Coupling HSP90 to the Human R2TP Co-Chaperone Complex. Nat. Commun. 2018, 9, 1501. [Google Scholar] [CrossRef] [Green Version]
- Millson, S.H.; Vaughan, C.K.; Zhai, C.; Ali, M.M.U.; Panaretou, B.; Piper, P.W.; Pearl, L.H.; Prodromou, C. Chaperone Ligand-Discrimination by the TPR-Domain Protein Tah1. Biochem. J. 2008, 413, 261–268. [Google Scholar] [CrossRef]
- da Silva, V.C.H.; Ramos, C.H.I. The Network Interaction of the Human Cytosolic 90kDa Heat Shock Protein Hsp90: A Target for Cancer Therapeutics. J. Proteom. 2012, 75, 2790–2802. [Google Scholar] [CrossRef]
- Seraphim, T.V.; Nano, N.; Cheung, Y.W.S.; Aluksanasuwan, S.; Colleti, C.; Mao, Y.-Q.; Bhandari, V.; Young, G.; Höll, L.; Phanse, S.; et al. Assembly Principles of the Human R2TP Chaperone Complex Reveal the Presence of R2T and R2P Complexes. Structure 2022, 30, 156–171.e12. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.; Cloutier, P.; Poitras, C.; Gauthier, M.-S.; Coulombe, B. Unphosphorylated Form of the PAQosome Core Subunit RPAP3 Binds Ribosomal Preassembly Complexes to Modulate Ribosome Biogenesis. J. Proteom. Res. 2022, 21, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Espitia-Hernández, P.; Chávez González, M.L.; Ascacio-Valdés, J.A.; Dávila-Medina, D.; Flores-Naveda, A.; Silva, T.; Ruelas Chacón, X.; Sepúlveda, L. Sorghum (Sorghum bicolor L.) as a Potential Source of Bioactive Substances and Their Biological Properties. Crit. Rev. Food Sci. Nutr. 2022, 62, 2269–2280. [Google Scholar] [CrossRef] [PubMed]
- Pazderů, K.; Hodoval, J.; Urban, J.; Pulkrábek, J.; Pačuta, V.; Adamčík, J. The Influence of Sweet Sorghum Crop Stand Arrangement on Biomass and Biogas Production. Plant Soil. Environ. 2014, 60, 433–438. [Google Scholar] [CrossRef]
- Jiménez, B.; Ugwu, F.; Zhao, R.; Ortí, L.; Makhnevych, T.; Pineda-Lucena, A.; Houry, W.A. Structure of Minimal Tetratricopeptide Repeat Domain Protein Tah1 Reveals Mechanism of Its Interaction with Pih1 and Hsp90. J. Biol. Chem. 2012, 287, 5698–5709. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Kakihara, Y.; Gribun, A.; Huen, J.; Yang, G.; Khanna, M.; Costanzo, M.; Brost, R.L.; Boone, C.; Hughes, T.R.; et al. Molecular Chaperone Hsp90 Stabilizes Pih1/Nop17 to Maintain R2TP Complex Activity That Regulates SnoRNA Accumulation. J. Cell Biol. 2008, 180, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Chagot, M.-E.; Jacquemin, C.; Branlant, C.; Charpentier, B.; Manival, X.; Quinternet, M. 1H, 15N and 13C Resonance Assignments of the Two TPR Domains from the Human RPAP3 Protein. Biomol. NMR Assign. 2015, 9, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Itsuki, Y.; Saeki, M.; Nakahara, H.; Egusa, H.; Irie, Y.; Terao, Y.; Kawabata, S.; Yatani, H.; Kamisaki, Y. Molecular Cloning of Novel Monad Binding Protein Containing Tetratricopeptide Repeat Domains. FEBS Lett. 2008, 582, 2365–2370. [Google Scholar] [CrossRef] [Green Version]
- von Morgen, P.; Hořejší, Z.; Macurek, L. Substrate Recognition and Function of the R2TP Complex in Response to Cellular Stress. Front. Genet. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Reva, B.A.; Finkelstein, A.V.; Skolnick, J. What Is the Probability of a Chance Prediction of a Protein Structure with an Rmsd of 6 å? Fold. Des 1998, 3, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, C.F.; Llorca, O. RPAP3 C-Terminal Domain: A Conserved Domain for the Assembly of R2TP Co-Chaperone Complexes. Cells 2020, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Lim, F.-L.; Finkler, A.; Fromm, H.; Slabas, A.R.; Knight, M.R. Transcriptomic Analysis of Sorghum bicolor Responding to Combined Heat and Drought Stress. BMC Genom. 2014, 15, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarshad, A.; Talei, D.; Torabi, M.; Rafiei, F.; Nejatkhah, P. Morphological and Biochemical Responses of Sorghum bicolor (L.) Moench under Drought Stress. SN Appl. Sci. 2021, 3, 81. [Google Scholar] [CrossRef]
- Eckert, K.; Saliou, J.-M.; Monlezun, L.; Vigouroux, A.; Atmane, N.; Caillat, C.; Quevillon-Chéruel, S.; Madiona, K.; Nicaise, M.; Lazereg, S.; et al. The Pih1-Tah1 Cochaperone Complex Inhibits Hsp90 Molecular Chaperone ATPase Activity. J. Biol. Chem. 2010, 285, 31304–31312. [Google Scholar] [CrossRef] [Green Version]
- Araujo, S.A.; Martins, G.H.; Quel, N.G.; Aragão, A.Z.B.; Morea, E.G.O.; Borges, J.C.; Houry, W.A.; Cano, M.I.N.; Ramos, C.H.I. Purification and Characterization of a Novel and Conserved TPR-Domain Protein That Binds Both Hsp90 and Hsp70 and Is Expressed in All Developmental Stages of Leishmania Major. Biochimie 2021, 182, 51–60. [Google Scholar] [CrossRef]
- D’Andrea, L. TPR Proteins: The Versatile Helix. Trends Biochem. Sci. 2003, 28, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.d.C.; Pinheiro, G.M.S.; Dahlström, K.M.; Souto, D.E.P.; Kubota, L.T.; Barbosa, L.R.S.; Ramos, C.H.I. On the Structure and Function of Sorghum bicolor CHIP (Carboxyl Terminus of Hsc70-Interacting Protein): A Link between Chaperone and Proteasome Systems. Plant Sci. 2020, 296, 110506. [Google Scholar] [CrossRef]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor Genome and the Diversification of Grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Russell, L.C.; Whitt, S.R.; Chen, M.-S.; Chinkers, M. Identification of Conserved Residues Required for the Binding of a Tetratricopeptide Repeat Domain to Heat Shock Protein 90. J. Biol. Chem. 1999, 274, 20060–20063. [Google Scholar] [CrossRef] [Green Version]
- Scheufler, C.; Brinker, A.; Bourenkov, G.; Pegoraro, S.; Moroder, L.; Bartunik, H.; Hartl, F.U.; Moarefi, I. Structure of TPR Domain–Peptide Complexes. Cell 2000, 101, 199–210. [Google Scholar] [CrossRef]
- Zeytuni, N.; Zarivach, R. Structural and Functional Discussion of the Tetra-Trico-Peptide Repeat, a Protein Interaction Module. Structure 2012, 20, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Back, R.; Dominguez, C.; Rothé, B.; Bobo, C.; Beaufils, C.; Moréra, S.; Meyer, P.; Charpentier, B.; Branlant, C.; Allain, F.H.-T.; et al. High-Resolution Structural Analysis Shows How Tah1 Tethers Hsp90 to the R2TP Complex. Structure 2013, 21, 1834–1847. [Google Scholar] [CrossRef] [Green Version]
- da Silva, V.C.H.; Cagliari, T.C.; Lima, T.B.; Gozzo, F.C.; Ramos, C.H.I. Conformational and Functional Studies of a Cytosolic 90 KDa Heat Shock Protein Hsp90 from Sugarcane. Plant Physiol. Biochem. 2013, 68, 16–22. [Google Scholar] [CrossRef]
- Tiroli-Cepeda, A.O.; Lima, T.B.; Balbuena, T.S.; Gozzo, F.C.; Ramos, C.H.I. Structural and Functional Characterization of the Chaperone Hsp70 from Sugarcane. Insights into Conformational Changes during Cycling from Cross-Linking/Mass Spectrometry Assays. J. Proteom. 2014, 104, 48–56. [Google Scholar] [CrossRef]
- Zanphorlin, L.M.; Lima, T.B.; Wong, M.J.; Balbuena, T.S.; Minetti, C.A.S.A.; Remeta, D.P.; Young, J.C.; Barbosa, L.R.S.; Gozzo, F.C.; Ramos, C.H.I. Heat Shock Protein 90 KDa (Hsp90) Has a Second Functional Interaction Site with the Mitochondrial Import Receptor Tom70. J. Biol. Chem. 2016, 291, 18620–18631. [Google Scholar] [CrossRef] [Green Version]
- Edelhoch, H. Spectroscopic Determination of Tryptophan and Tyrosine in Proteins. Biochemistry 2001, 6, 1948–1954. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Estimation of Protein Secondary Structure from Circular Dichroism Spectra: Comparison of CONTIN, SELCON, and CDSSTR Methods with an Expanded Reference Set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Sigrist, C.J.A.; De Castro, E.; Cerutti, L.; Cuche, B.A.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and Continuing Developments at PROSITE. Nucleic Acids Res. 2013, 41, D344–D347. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam Protein Families Database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L.; et al. InterPro: The Integrative Protein Signature Database. Nucleic Acids Res. 2009, 37, D211–D215. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Baek, M.; Dimaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate Prediction of Protein Structures and Interactions Using a Three-Track Neural Network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of Protein Structures: Patterns of Nonbonded Atomic Interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Delano, W.L. PyMOL: An Open-Source Molecular Graphics Tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Borges, J.C.; Ramos, C.H.I. Spectroscopic and Thermodynamic Measurements of Nucleotide-Induced Changes in the Human 70-KDa Heat Shock Cognate Protein. Arch. Biochem. Biophys. 2006, 452, 46–54. [Google Scholar] [CrossRef]
- Philo, J.S. SEDNTERP: A Calculation and Database Utility to Aid Interpretation of Analytical Ultracentrifugation and Light Scattering Data. Eur. Biophys. J. 2023. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Measured | Sphere |
|---|---|---|
| Molecular Mass (kDa) | 41.7 ± 0.3 | 44.1 a |
| D (10−7 cm2 s−1) | 7.5 ± 0.3 | 9.2 b |
| Rs (Å) | 38 ± 0.0 | 23 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonio, L.M.; Martins, G.H.; Aragão, A.Z.B.; Quel, N.G.; Zazeri, G.; Houry, W.A.; Ramos, C.H.I. Unveiling the Role of Sorghum RPAP3 in the Function of R2TP Complex: Insights into Protein Assembly in Plants. Plants 2023, 12, 2925. https://doi.org/10.3390/plants12162925
Antonio LM, Martins GH, Aragão AZB, Quel NG, Zazeri G, Houry WA, Ramos CHI. Unveiling the Role of Sorghum RPAP3 in the Function of R2TP Complex: Insights into Protein Assembly in Plants. Plants. 2023; 12(16):2925. https://doi.org/10.3390/plants12162925
Chicago/Turabian StyleAntonio, Larissa Machado, Gustavo Henrique Martins, Annelize Zambon Barbosa Aragão, Natália Galdi Quel, Gabriel Zazeri, Walid A. Houry, and Carlos Henrique Inacio Ramos. 2023. "Unveiling the Role of Sorghum RPAP3 in the Function of R2TP Complex: Insights into Protein Assembly in Plants" Plants 12, no. 16: 2925. https://doi.org/10.3390/plants12162925