
Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints

 , ,
, ,
Abstract
:1. Introduction
2. Bibliometric Analysis of Research on PEOs’ Biopesticidal Potential
PEOs as Biopesticides: Research Trends
3. PEOs as “New Age Biopesticides”
3.1. PEOs as Bioherbicides
Effect of PEO Supplementation on the Photosynthesis and Ultrastructure Integrity of Weeds
3.2. PEOs as Bioinsecticides
Current Status and Outlook for the Identification of Receptor Modulators That Enhance PEOs’ Insecticidal Effects
3.3. PEOs as Biofungicides
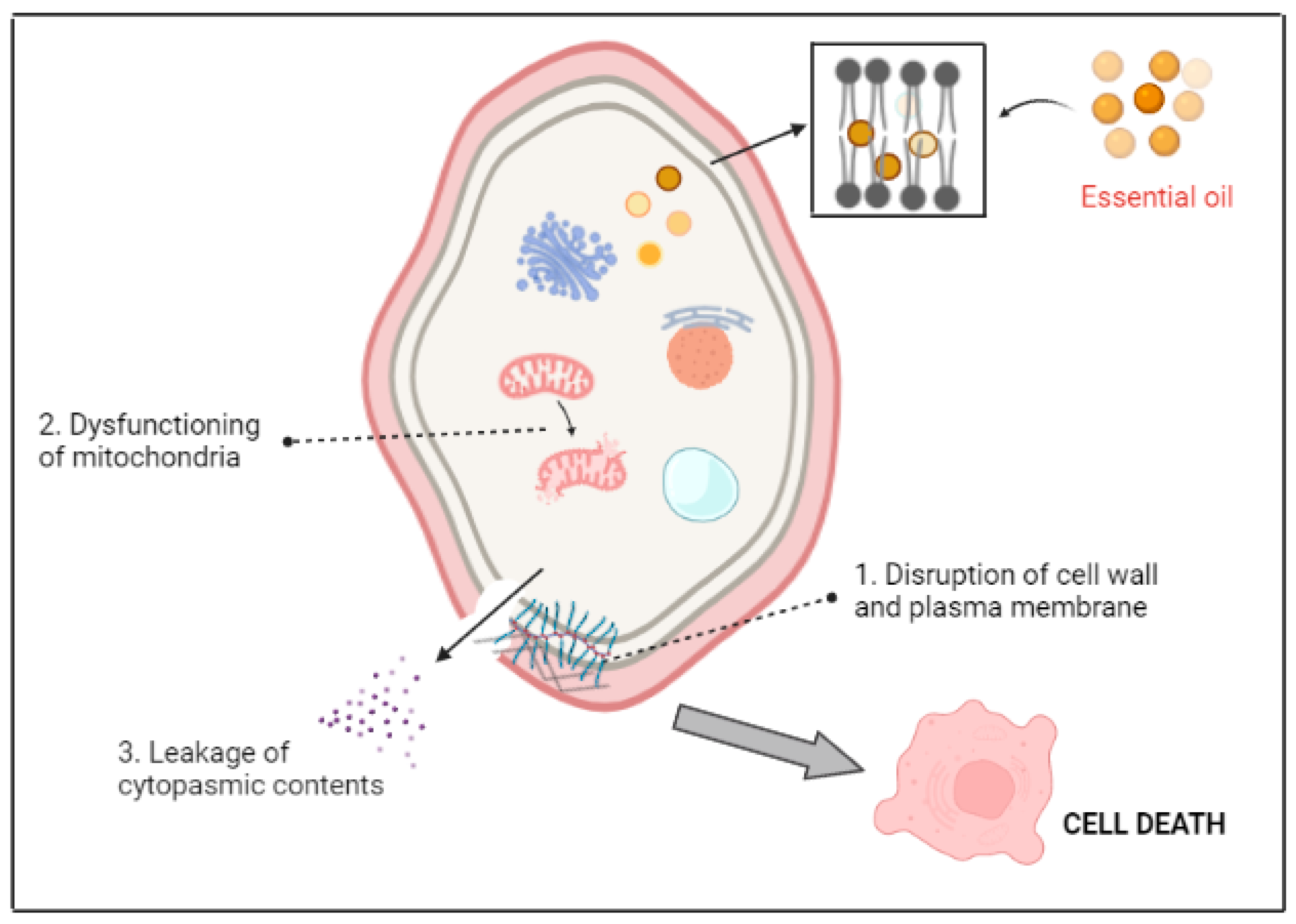
Mechanism of Action of PEOs in Fungal Cells
4. Technological Advancements in the Research on Biopesticides
4.1. Use of Nanoformulations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of PEO or Its Active Component | Encapsulation/ Emulsifying Agent | Method(s) Used for Encapsulation | Biological Target | Effects | Reference(s) |
|---|---|---|---|---|---|
| Bioherbicides | |||||
| Satureja hortensis L. | Carbohydrate and protein natural polymers (gum arabic /gelatin, apple pectin, and gelatin) and cross-linkers (citric acid and transglutaminase enzyme) | Complex coacervation | Amaranthus retroflexus L. and Solanum lycopersicum L. | (a) Inhibition in germination and growth of A. retroflexus (b) Inhibition was comparable to chemical herbicide (Metribuzin) | [157] |
| Foeniculum vulgare Mill. | Tween 80 | Ultrasonic emulsification | Phalaris minor Retz., Avena ludoviciana Durieu, Rumex dentatus L., and Medicago denticulata Willd | (a) Stability of nanoemulsions persistent after 30 days of storage (b) Inhibition of target weeds at as low as 0.05 wt.% dose of nanoemulsion (c) Individual components of PEO showed weed growth at high doses | [158] |
| Carum carvi L. and Mentha piperita L. | Commercial multifunctional adjuvant ATPOLAN BIO 80 EC | Emulsification | Echinochloa crus-galli (L.) P. Beauv. | (a) Foliar injuries were reported and photosynthetic efficiency was reduced in weed plant at 2.5% adjuvant dose (b) Crop plant was unaffected | [159] |
| Bioinsecticides | |||||
| Cymbopogon nardus (L.) Rendle | Tween 80 | Emulsification | Oryzaephilus surinamensis L. | (a) Nanoemulsions displayed greater adult mortality than pure oil in both male and female adults (b) LC50 value of nanoemulsions was significantly less than that of pure oil depicting 50% mortality at lower concentrations of nanoemulsions | [160] |
| Pimpinella anisum L., Foeniculum vulgare Mill., and Mentha piperita L. | Tween 80 | Self-emulsification | Bactrocera oleae Gmelin | (a) No residual toxicity reported (b) All nanoformulations were capable of reducing oviposition punctures (c) P. anisum displayed the highest percentage of repellent activity followed by F. vulgare and M. piperita | [161] |
| Rosmarinus officinalis L., Lavandula angustifolia Mill., Mentha piperita L. | Lipid: Softisan Surfactants: Kolliphor RH40 and Labrafil | Phase inversion temperature (PIT) method | Aphis gossypii (synonym of Aphis forbesi weed), Spodoptera littoralis Boisduval, and Tuta absoluta Meyrick | (a) Bioassay confirmed high mortality in A. gossypii treated with oil-loaded lipid carriers (b) Reduction in progeny was observed. With no mortality observed, the feeding activity of S. littoralis was reduced with L. angustifolia and R. officinalis carriers (c) No effect was observed on T. absoluta | [162] |
| Foeniculum vulgare Mill., Mentha piperita L., and Citrus sinensis (L.) Osbeck | 5% Tween | Spontaneous emulsification | Rhyzopetha dominica Fabricius | (a) All the PEOs displayed repellent activity against the tested storage pest. F. vulgare oil displayed the least activity (b) Insects recovered after 24 h of treatment and habituation was validated | [52] |
| Baccharis reticularia DC. and oil components (limonene, α-pinene, and β-pinene) | Nonionic surfactant (mixture of sorbitan monooleate, polysorbate 80, and/or polysorbate 20) | Low-energy titration | Tribolium castaneum Herbst | (a) Both the PEO and its components displayed good repellent activity at 8.8 μg cm−2 | [149] |
| Citrus sinensis (L.) Osbeck | Silica (SiO2) | Sol–gel microencapsulation | Spodoptera littoralis Boisduval and Aphis gossypii (synonym of Aphis forbesi Weed) | (a) Insecticidal activity against cotton leafworm (S. littoralis) (b) Reduction in the fertility and number of A. gossypii offsprings | [163] |
| Pimpinella anisum L., Artemisia vulgaris L., Foenicum vulgare Mill., Allium sativum L., Lavandula angustifolia Mill., Mentha piperita L., Rosmarinus officinalis L., and Salvia officinalis Pall. | 5% Tween 80 | Spontaneous emulsification | Tribolium confusum Duval | (a) All PEOs displayed repellent activity against the tested storage pest (b) With a RC50 value of 0.033 mg, the PEO of P. anisum exhibited maximum repellent activity | [164] |
| Cymbopogon citratus (DC.) Stapf and Eucalyptus globulus Labill. | Polysorbate 80 | High-energy emulsification | Musca domestica L. and Lucilia cuprina Wiedemann | (a) No adulticidal activity with E. globulus oil (b) Free C. citratus oil displayed better adulticial activity as compared to nanoemulsion; however, the use of nanoemulsion is suitable as it prevents the volatilization and degradation of oil (c) The concentration of nanoemulsion, time of treatment exposure, and encapsulation method play significant roles in mediating adulticidal activity | [165] |
| Lippia multiflora Mold. | 89.75% Hydrolate and 0.25% Chitosan | Low-energy emulsification | Plutella xylostella L., Brevicoryne brassicae L., Hellula undalis Fabricius, Spodoptera exigua Hubner, and Bemisia tabaci Gennadius | (a) PEO nanoemulsions were tested against synthetic pesticide, i.e., Karate 5 EC (Lambda cyhalothrin 52 gL−1) in two regions of Ivory Coast (b) Treatment with nanoemulsion reduced the damage of cabbage head as compared to synthetic pesticide, thereby signifying better protection against selected insects | [166] |
| Biofungicides | |||||
| Cymbopogon martini (Roxb.) W.Watson | Lipids (cocoa butter and sesame oil) | Melt emulsification method | Aspergillus nomius Kurtzman, B.W.Horn and Hesselt. | (a) 100% inhibition of A. nomius was displayed by nanostructured lipid carriers | [153] |
| d-Limonene | Emulsifiers | Phase transition composition emulsification | Pyricularia oryzae Cavara, Rhizoctonia solani J.G.Kühn, Colletortrichum gloeosporiodes (Penz.) Penz. and Sacc., and Phomopsis amygdali (Delacr.) J.J.Tuset and M.T.Portilla. | (a) Stability of nanoemulsions was tested and found to increase with an increase in dose of emulsifier (b) Emulsifier (EL40) displayed the highest stability © Inhibition of growth of tested fungal pathogens was reported | [167] |
| Cinnamomum verum J.Presl, Thymus vulgaris L., and Melaleuca alternifolia (Maiden and Betche) Cheel | Crodamol GTCC and polysorbate 80 | Phase inversion composition method (PIC) and ultrasonication | Fusarium culmorum (Wm.G.Sm.) Sacc., Phytophthora cactorum (Lebert and Cohn) J.Schröt. Trichophyton mentagrophytes (C.P.Robin) Sabour., Microsporum gypseum (E.Bodin) Guiart and Grigoraki, Aspergillus niger Tiegh., and Scopulariopsis brevicaulis (Sacc.) Brainier | (a) Fungicidal activities of pure oil, macroemulsions (PIC-based), and nanoemulsions (ultrasonication-based) were reported (b) Nanoemulsions prepared using high-energy ultrasonication showed better fungicidal activities (c) Among the different PEOs studied, M. alternifolia oil displayed the best activity | [168] |
4.2. Use of Bioinformatics
4.3. Transcriptomic Profiling to Elucidate PEOs’ Insecticidal Efficacy
5. Efficiency of PEOs in Promoting Sustainable Agriculture
6. Constraints, Safety, and Commercialization Considerations
6.1. Constraints
- The increase in quantity and frequency of PEO application is a result of the degradation and volatilization of essential metabolites upon exposure to air, sunlight, moisture, and high temperatures. As was previously stated, the nanoencapsulation of PEOs can practically serve this purpose if used at the appropriate concentration. For example, Su et al. [191] developed several derivatives based on PEOs and β-methoxyacrylate to improve the stability of fungicidal oil formulations.
- Due to technological barriers, these natural compounds are less economical and not as popular among farmers as synthetic pesticides. However, this problem can be overcome by providing subsidies to farmers and highlighting the long-term benefits of botanical pesticides by various governmental and nongovernmental agencies.
- In contrast to chemical pesticides that are usually lethal, biopesticides often function by preventing the pathogens from proliferating further. Thus, combined spraying of chemical and biological pesticides is being adopted to get rid of pre-existing pests. However, to achieve a complete phase-out of chemical pesticides, more research needs to be conducted to explore the broad spectrum of lethal natural products as biopesticides.
- The requirements of a large arable land area, source plants, and proper climatic conditions are additional barriers to the use of biopesticides. The collection of plant material needs to be conducted at the right time to preserve the quality of the raw materials required, thus making it seasonal. The purity of products is also restricted due to the differences in chemical composition of plants’ active ingredients in different geographical areas [192].
6.2. Safety
- Harmful organic solvents used for PEO solubilization, as well as toxic doses acquired by nonpesticidal active components of plant products, raise concerns about the safety of plant-based biopesticides [193].
- The growing body of research on nanoemulsions raises alarms about the safety of such formulations, partly due to the involvement of organic solvents in their lab-scale preparation and the use of spray drying on a commercial scale, which is potentially hazardous to the environment.
6.3. Commercialization
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations (UN). Department of economic and social affairs, population division. In World Population Prospects: The 2022 Revision; United Nations: New York, NY, USA, 2022. [Google Scholar]
- Kukal, M.S.; Irmak, S. Climate-driven crop yield and yield variability and climate change impacts on the US Great Plains agricultural production. Sci. Rep. 2018, 8, 3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World. Transforming Food Systems for Food Security, Improved Nutrition and Affordable Healthy Diets for All; FAO: Rome, Italy, 2021. [Google Scholar]
- Leila, B.; El-Hafid, N. Biofertilizers and biopesticides: Microbes for sustainable agriculture. Adv. Plant Microb. Sustain. Agric. 2020, 565, 257–279. [Google Scholar]
- Copplestone, J.F. The development of the WHO recommended classification of pesticides by hazard. Bull. World Health Organ. 1988, 66, 545–551. [Google Scholar]
- Marutescu, L.; Chifiriuc, M.C. Molecular mechanisms of pesticides toxicity. In New Pesticides and Soil Sensors; Grumezescu, A.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 393–435. [Google Scholar]
- Isman, M.B. Challenges of pest management in the twenty first century: New tools and strategies to combat old and new foes alike. Front. Agron. 2019, 1, 2. [Google Scholar] [CrossRef]
- Singh, R.; Singh, H.; Raghubanshi, A.S. Challenges and opportunities for agricultural sustainability in changing climate scenarios: A perspective on Indian agriculture. Trop. Ecol. 2019, 60, 167–185. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef]
- Chaudhary, S.; Kanwar, R.K.; Sehgal, A.; Cahill, D.M.; Barrow, C.J.; Sehgal, R.; Kanwar, J.R. Progress on Azadirachta indica based biopesticides in replacing synthetic toxic pesticides. Front. Plant Sci. 2017, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Dhuldhaj, U.P.; Singh, R.; Singh, V.K. Pesticide contamination in agro-ecosystems: Toxicity, impacts, and bio-based management strategies. Environ. Sci. Pollut. Res. 2023, 30, 9243–9270. [Google Scholar] [CrossRef]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. Fores. Ecol. Manage. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Duke, S.O.; Pan, Z.; Bajsa-Hirschel, J.; Boyette, C.D. The potential future roles of natural compounds and microbial bioherbicides in weed management in crops. Adv. Weed Sci. 2022, 40, e020210054. [Google Scholar] [CrossRef]
- Glare, T.R. Types of biopesticides. In Biopesticides Handbook; Nollet, L.M.L., Rathore, H.S., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 7–25. [Google Scholar]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibn, H.; Tyrwhitt, T. Orpheus, Peri Lithôn De Lapidibus, Poema Orpheo a Quibusdam Adscriptum; Payne, White, and Elmsly: London, UK, 1781. [Google Scholar]
- Birch, E.A.N.; Begg, G.S.; Squire, G.R. How agro-ecological research helps to address food security issues under new IPM and pesticide reduction policies for global crop production systems. J. Exp. Bot. 2011, 62, 3251–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duke, S.O.; Evidente, A.; Vurro, M. Natural products in pest management: Innovative approaches for increasing their use. Pest Manag. Sci. 2019, 75, 2299–2300. [Google Scholar] [CrossRef] [PubMed]
- Abbey, L.; Abbey, J.; Leke-Aladekoba, A.; Iheshiulo, E.M.A.; Ijenyo, M. Biopesticides and biofertilizers: Types, production, benefits, and utilization. In Byproducts from Agriculture and Fisheries: Adding Value for Food, Feed, Pharma, and Fuels; Simpson, B.K., Aryee, A.N.A., Toldrá, F., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 479–500. [Google Scholar]
- Alliance, D.; Santos, C.B.R.D.; Barbosa, L.M.C.; Gomes, J.S.; Lobato, C.C.; Viana, J.C.; Ramos, R.D.S.; Ferreira, R.M.D.A.; Souto, R.N.P. Chemical study, predictions in silico and larvicide activity of the essential oil of root Philodendron deflexum Poepp. J. Comput. Theor. Nanosci. 2017, 14, 3330–3337. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, K.; Kumar, A.; Sharma, S.; Pathak, R.; Jangir, M. Ocimum sp.: Source of biorational pesticides. Ind. Crops Prod. 2018, 122, 686–701. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial plant compounds, extracts and essential oils: An updated review on their effects and putative mechanisms of action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Kaur, S.; Negi, K.; Kumari, S.; Saini, V.; Batish, D.R.; Kohli, R.K. Assessment of in vitro antioxidant activity of essential oil of Eucalyptus citriodora (lemon-scented Eucalypt; Myrtaceae) and its major constituents. LWT 2012, 48, 237–241. [Google Scholar] [CrossRef]
- Grewal, K.; Joshi, J.; Rathee, S.; Kaur, S.; Singh, H.P.; Batish, D.R. Chemical composition and potential of Eucalyptus camaldulensis Dehnh. essential oil and its major components as anti-inflammatory and anti-leishmanial agent. J. Essent. Oil-Bear. Plants 2022, 25, 419–429. [Google Scholar] [CrossRef]
- Fagodia, S.K.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phytotoxicity and cytotoxicity of Citrus aurantiifolia essential oil and its major constituents: Limonene and citral. Ind. Crops Prod. 2017, 108, 708–715. [Google Scholar] [CrossRef]
- Sharma, M.; Grewal, K.; Jandrotia, R.; Batish, D.R.; Singh, H.P.; Kohli, R.K. Essential oils as anticancer agents: Potential role in malignancies, drug delivery mechanisms, and immune system enhancement. Biomed. Pharmacother. 2022, 146, 112514. [Google Scholar] [CrossRef]
- Cadena, M.B.; Preston, G.M.; Van der Hoorn, R.A.; Townley, H.E.; Thompson, I.P. Species-specific antimicrobial activity of essential oils and enhancement by encapsulation in mesoporous silica nanoparticles. Ind. Crops Prod. 2018, 122, 582–590. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Chemical characterization and allelopathic potential of volatile oil of Eucalyptus tereticornis against Amaranthus viridis. J. Plant Interact. 2011, 6, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Maria, G.A.; Riccardo, N. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Em. J. Food Agric. 2020, 22, 522–532. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, B.; Wani, M.S.; Gupta, R.C.; Habeeb, T.H. Determination of the volatile composition in essential oil of Azadirachta indica A. Juss from different areas of north Indian plains by Gas Chromatography/Mass Spectrometry (GC/MS). Analyt. Chem. Lett. 2021, 11, 73–82. [Google Scholar] [CrossRef]
- Chatterjee, S.; Gupta, S.; Variyar, P.S. Comparison of essential oils obtained from different extraction techniques as an aid in identifying aroma significant compounds of nutmeg (Myristica fragrans). Nat. Prod. Comm. 2015, 10, 1443–1446. [Google Scholar] [CrossRef]
- Sharma, S.; Mulrey, L.; Byrne, M.; Jaiswal, A.K.; Jaiswal, S. Encapsulation of essential oils in nanocarriers for active food packaging. Foods 2022, 11, 2337. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Singh, R.; Kumari, T.; Verma, P.; Singh, B.P.; Raghubanshi, A.S. Compatible package-based agriculture systems: An urgent need for agro-ecological balance and climate change adaptation. Soil Ecol. Lett. 2022, 4, 187–212. [Google Scholar] [CrossRef]
- Scopus. 2022. Available online: https://www.scopus.com/results/results.uri? (accessed on 21 December 2022).
- Web of Science. 2022. Available online: https://www.webofscience.com/wos/woscc/summary/ddb0f776-fae7-4795-9de6-d459acde649a-988bbc30/relevance/1 (accessed on 21 December 2022).
- Mossa, A.T.H. Green pesticides: Essential oils as biopesticides in insect-pest management. J. Environ. Sci. Technol. 2016, 9, 354. [Google Scholar] [CrossRef] [Green Version]
- Rana, A.; Matiyani, M.; Tewari, C.; Negi, P.B.; Arya, M.C.; Das, V.; Pal, M.; Sahoo, N.G. Functionalized graphene oxide based nanocarrier for enhanced cytotoxicity of Juniperus squamata root essential oil against breast cancer cells. J. Drug Deliv. Sci. Technol. 2022, 72, 103370. [Google Scholar] [CrossRef]
- Khan, M.; Khan, S.T.; Khan, M.; Mousa, A.A.; Mahmood, A.; Alkhathlan, H.Z. Chemical diversity in leaf and stem essential oils of Origanum vulgare L. and their effects on microbicidal activities. AMB Express 2019, 9, 176. [Google Scholar] [CrossRef]
- Weluwanarak, T.; Changbunjong, T.; Leesombun, A.; Boonmasawai, S.; Sungpradit, S. Effects of Piper nigrum L. fruit essential oil toxicity against stable fly (Diptera: Muscidae). Plants 2023, 12, 1043. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; De Francesco, C.; Cornara, L.; Barreca, D.; Bellocco, E.; Ginestra, G.; Mandalari, G.; Trombetta, D. Feijoa fruit peel: Micro-morphological features, evaluation of phytochemical profile, and biological properties of its essential oil. Antioxidants 2020, 8, 320. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, W.W.; Kumar, K.J.S.; Lee, H.J.; Tsao, N.W.; Wang, S.Y. Anti-melanogenic activity of Calocedrus formosana wood essential oil and its chemical composition analysis. Plants 2021, 1, 62. [Google Scholar] [CrossRef]
- Zapata, N.; Smagghe, G. Repellency and toxicity of essential oils from the leaves and bark of Laurelia sempervirens and Drimys winteri against Tribolium castaneum. Ind. Crops Prod. 2010, 32, 405–410. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Saleh, I.; El-Razek, M.H.A.; Elkarim, A.S.A.; El-Amier, Y.A.; Mohamed, T.A.; El Gendy, A.E.G.; Afifi, S.M.; Esatbeyoglu, T.; Elshamy, A.I. Chemical profiling of significant antioxidant and phytotoxic microwave-extracted essential oil from Araucaria heterophylla resin. Separations 2023, 10, 141. [Google Scholar] [CrossRef]
- El-Hefny, M.; Abo Elgat, W.; Al-Huqail, A.; Ali, H. Essential and recovery oils from Matricaria chamomilla flowers as environmentally friendly fungicides against four fungi isolated from cultural heritage objects. Processes 2019, 7, 809. [Google Scholar] [CrossRef] [Green Version]
- El Asbahani, A.; Miladi, K.; Badri, W.; Sala, M.; Addi, E.A.; Casabianca, H.; El Mousadik, A.; Hartmann, D.; Jilale, A.; Renaud, F.N.R.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F.; Zappalà, L.; Palmeri, V. Bioactivity of essential oil-based nano-biopesticides toward Rhyzopertha dominica (Coleoptera: Bostrichidae). Ind. Crops Prod. 2021, 162, 113257. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxic effects of commercial Eucalyptus citriodora, Lavandula angustifolia, and Pinus sylvestris essential oils on weeds, crops, and invasive species. Molecules 2019, 24, 2847. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Chemical profiling, cytotoxicity and phytotoxicity of foliar volatiles of Hyptis suaveolens. Ecotoxicol. Environ. Saf. 2019, 171, 863–870. [Google Scholar] [CrossRef]
- De Mastro, G.; El Mahdi, J.; Ruta, C. Bioherbicidal potential of the essential oils from Mediterranean Lamiaceae for weed control in organic farming. Plants 2021, 10, 818. [Google Scholar] [CrossRef]
- Giannini, V.; Harris, J.R.; Todde, P.; McElroy, J.S. Concurrent weed growth suppression with essential oils and species-specific response to fractionated coconut oil. Ind. Crops Prod. 2022, 182, 114850. [Google Scholar] [CrossRef]
- Gruľová, D.; Baranová, B.; Sedlák, V.; De Martino, L.; Zheljazkov, V.D.; Konečná, M.; Poráčová, J.; Caputo, L.; De Feo, V. Juniperus horizontalis Moench: Chemical composition, herbicidal and insecticidal activities of its essential oil and of its main component, sabinene. Molecules 2022, 27, 8408. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic interactions and allelochemicals: New possibilities for sustainable weed management. Crit. Rev. Plant. Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Benvenuti, S.; Cioni, P.; Flamini, G.; Pardossi, A. Weeds for weed control: Asteraceae essential oils as natural herbicides. Weed Res. 2017, 57, 342–353. [Google Scholar] [CrossRef]
- Danna, C.; Cornara, L.; Smeriglio, A.; Trombetta, D.; Amato, G.; Aicardi, P.; De Martino, L.; De Feo, V.; Caputo, L. Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ essential oils as potential natural herbicides. Molecules 2021, 26, 6749. [Google Scholar] [CrossRef] [PubMed]
- Polito, F.; Kouki, H.; Khedhri, S.; Hamrouni, L.; Mabrouk, Y.; Amri, I.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical composition and phytotoxic and antibiofilm activity of the essential oils of Eucalyptus bicostata, E. gigantea, E. intertexta, E. obliqua, E. pauciflora and E. tereticornis. Plants 2022, 11, 3017. [Google Scholar] [CrossRef]
- Malaspina, P.; Papaianni, M.; Ranesi, M.; Polito, F.; Danna, C.; Aicardi, P.; Cornara, L.; Woo, S.L.; De Feo, V. Eucalyptus cinerea and E. nicholii by-products as source of bioactive compounds for agricultural applications. Plants 2022, 11, 2777. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Lamia, H.; Mohsen, H.; Samia, G.; Bassem, J. Chemical composition, bio-herbicidal and antifungal activities of essential oils isolated from Tunisian common cypress (Cupressus sempervirens L.). J. Med. Plant Res. 2013, 7, 1070–1080. [Google Scholar]
- Ismail, A.; Habiba, K.; Yassine, M.; Mohsen, H.; Bassem, J.; Lamia, H. Essential oils of Tunisian Pinus radiata D. Don, chemical composition and study of their herbicidal activity. Vietnam. J. Chem. 2021, 59, 247–252. [Google Scholar]
- Kordali, S.; Kabaagac, G.; Sen, İ.; Yilmaz, F.; Najda, A. Phytotoxic effects of three Origanum species extracts and essential oil on seed germinations and seedling growths of four weed species. Agronomy 2022, 12, 2581. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poonpaiboonpipat, T.; Pangnakorn, U.; Suvunnamek, U.; Teerarak, M.; Charoenying, P.; Laosinwattana, C. Phytotoxic effects of essential oil from Cymbopogon citratus and its physiological mechanisms on barnyardgrass (Echinochloa crus-galli). Ind. Crops Prod. 2013, 41, 403–407. [Google Scholar] [CrossRef]
- Singh, N.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Yadav, S.S. Chemical characterization, phytotoxic, and cytotoxic activities of essential oil of Mentha longifolia. Environ. Sci. Pollut. Res. 2020, 27, 13512–13523. [Google Scholar] [CrossRef]
- Pouresmaeil, M.; Nojadeh, M.S.; Movafeghi, A.; Maggi, F. Exploring the bio-control efficacy of Artemisia fragrans essential oil on the perennial weed Convolvulus arvensis: Inhibitory effects on the photosynthetic machinery and induction of oxidative stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Watson, S.B.; Asolkar, R.N.; Boddy, L.G. Sarmentine, a natural herbicide from Piper species with multiple herbicide mechanisms of action. Front. Plant Sci. 2015, 6, 222. [Google Scholar] [CrossRef] [Green Version]
- Issa, M.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Yadav, S.S.; Kumari, A. Appraisal of phytotoxic, cytotoxic and genotoxic potential of essential oil of a medicinal plant Vitex negundo. Ind. Crops Prod. 2020, 145, 112083. [Google Scholar] [CrossRef]
- Lins, L.; Dal Maso, S.; Foncoux, B.; Kamili, A.; Laurin, Y.; Genva, M.; Jijakli, M.H.; De Clerck, C.; Fauconnier, M.L.; Deleu, M. Insights into the relationships between herbicide activities, molecular structure and membrane interaction of cinnamon and citronella essential oils components. Int. J. Mol. Sci. 2019, 20, 4007. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils from Foeniculum vulgare Miller as a safe environmental insecticide against the aphid Myzus persicae Sulzer. Environ. Sci. Pollut. Res. 2018, 25, 10904–10910. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Zorzetto, C.; Sánchez-Mateo, C.C.; Santini, G.; Canale, A.; Maggi, F. Insecticidal activity of the essential oil from Schizogyne sericea (Asteraceae) on four insect pests and two non-target species. Entomol. Gen. 2019, 39, 9–18. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Ponsankar, A.; Thanigaivel, A.; Edwin, E.S.; Selin-Rani, S.; Chellappandian, M.; Pradeepa, V.; Lija-Escaline, J.; Kalaivani, K. Comparative analysis of mosquito (Diptera: Culicidae: Aedes aegypti Liston) responses to the insecticide Temephos and plant derived essential oil derived from Piper betle L. Ecotoxicol. Environ. Saf. 2017, 139, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Sarma, R.; Khanikor, B.; Mahanta, S. Essential oil from Citrus grandis (Sapindales: Rutaceae) as insecticide against Aedes aegypti (L.) (Diptera: Culicidae). Int. J. Mosq. Res. 2017, 4, 88–92. [Google Scholar]
- Kordali, Ş.; Kesdek, M.; Akkan, Ü.; Usanmaz Bozhüyük, A. Insecticidal activities of some plant essential oils on Rhyzopertha dominica (F.) and Sitophilus granarius L. (Coleoptera: Bostrichidae and Curculionidae) adults. ACU J. For. Fac. 2022, 23, 127–134. [Google Scholar] [CrossRef]
- Üstüner, T.; Kordali, Ş.; Bozhüyük, A.U.; Kesdek, M. Investigation of pesticidal activities of essential oil of Eucalyptus camaldulensis Dehnh Rec. Nat. Prod. 2018, 12, 557–568. [Google Scholar]
- Gokturk, T.; Kordali, S.; Ak, K.; Kesdek, M.; Bozhuyuk, A.U. Insecticidal effects of some essential oils against Tribolium confusum (du Val.) and Acanthoscelides obtectus (Say), (Coleoptera: Tenebrionidae and Bruchidae) adults. Int. J. Trop. Insect Sci. 2020, 40, 637–643. [Google Scholar] [CrossRef]
- Kordali, Ş.; Usanmaz, A.; Bayrak, N.; Çakır, A. Fumigation of volatile monoterpenes and aromatic compounds against adults of Sitophilus granarius (L.) (Coleoptera: Curculionidae). Rec. Nat. Prod. 2017, 11, 362–373. [Google Scholar]
- Khani, A.; Asghari, J. Insecticide activity of essential oils of Mentha longifolia, Pulicaria gnaphalodes and Achillea wilhelmsii against two stored product pests, the flour beetle, Tribolium castaneum, and the cowpea weevil, Callosobruchus maculatus. J. Insect Sci. 2012, 12, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jairoce, C.F.; Teixeira, C.M.; Nunes, C.F.; Nunes, A.M.; Pereira, C.M.; Garcia, F.R. Insecticide activity of clove essential oil on bean weevil and maize weevil. Rev. Bras. De Eng. Agric. E Ambient. 2016, 20, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Santini, G.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; Canale, A.; Maggi, F. The essential oil from industrial hemp (Cannabis sativa L.) by-products as an effective tool for insect pest management in organic crops. Ind. Crops Prod. 2018, 122, 308–315. [Google Scholar] [CrossRef]
- Bhavya, M.; Chandu, A.; Devi, S.S. Ocimum tenuiflorum oil, a potential insecticide against rice weevil with anti-acetylcholinesterase activity. Ind. Crops Prod. 2018, 126, 434–439. [Google Scholar] [CrossRef]
- Cossetin, L.F.; Santi, E.M.T.; Cossetin, J.F.; Dillmann, J.B.; Baldissera, M.D.; Garlet, Q.I.; de Souza, T.P.; Loebens, L.; Heinzmann, B.M.; Machado, M.M. In vitro safety and efficacy of lavender essential oil (Lamiales: Lamiaceae) as an insecticide against houseflies (Diptera: Muscidae) and blowflies (Diptera: Calliphoridae). J. Econ. Entomol. 2018, 111, 1974–1982. [Google Scholar] [CrossRef]
- Rajkumar, V.; Gunasekaran, C.; Christy, I.K.; Dharmaraj, J.; Chinnaraj, P.; Paul, C.A. Toxicity, antifeedant and biochemical efficacy of Mentha piperita L. essential oil and their major constituents against stored grain pest. Pestic. Biochem. Phys. 2019, 156, 138–144. [Google Scholar] [CrossRef]
- Gal, T.Z.; Aussenberg, E.R.; Burdman, S.; Kapulnik, Y.; Koltai, H. Expression of a plant expansin is involved in the establishment of root knot nematode parasitism in tomato. Planta 2006, 224, 155–162. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Barbosa, P.; Vieira, P.; Vicente, C.S.L.; Figueiredo, A.C.; Mota, M. Phytochemicals as biopesticides against the pinewood nematode Bursaphelenchus xylophilus: A review on essential oils and their volatiles. Plants 2021, 10, 2614. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Lapied, B.; Jankowski, W.; Stankiewicz, M. The unusual action of essential oil component, menthol, in potentiating the effect of the carbamate insecticide, bendiocarb. Pestic. Biochem. Physiol. 2019, 158, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.D.; Temeyer, K.B.; Day, T.A.; de León, A.A.P.; Kimber, M.J.; Coats, J.R. Interaction of plant essential oil terpenoids with the southern cattle tick tyramine receptor: A potential biopesticide target. Chem. Biol. Interact. 2017, 263, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tayoub, G.; Odeh, A.; Ghanem, I. Chemical composition and fumigation toxicity of Laurus nobilis L. and Salvia officinalis L. essential oils on larvae of khapra beetle (Trogoderma granarium Everts). Herba. Pol. 2012, 58, 26–37. [Google Scholar]
- Ebadollahi, A.; Ashrafi Parchin, R.; Farjaminezhad, M. Phytochemistry, toxicity and feeding nhibitory activity of Melissa occinalis L. essential oil against a cosmopolitan insect pest; Tribolium castaneum Herbst. Toxin Rev. 2016, 35, 77–82. [Google Scholar] [CrossRef]
- Abd-Elhady, H. Insecticidal activity and chemical composition of essential oil from Artemisia judaica L. against Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. Plant Protect. Res. 2012, 52, 347–352. [Google Scholar] [CrossRef]
- Saad, M.M.G.; El-Deeb, D.A.; Abdelgaleil, S.A.M. Insecticidal potential and repellent and biochemical effects of phenylpropenes and monoterpenes on the red flour beetle, Tribolium castaneum Herbst. Environ. Sci. Pollut. Res. 2019, 26, 6801–6810. [Google Scholar] [CrossRef]
- Huang, S.H.; Xian, J.D.; Kong, S.Z.; Li, Y.C.; Xie, J.H.; Lin, J.; Chen, J.N.; Wang, H.F.; Su, Z.R. Insecticidal activity of pogostone against Spodoptera litura and Spodoptera exigua (Lepidoptera: Noctuidae). Pest Manage. Sci. 2014, 70, 510–516. [Google Scholar] [CrossRef]
- Taghizadeh-Saroukolai, A.; Nouri-Ganbalani, G.; Rafiee-Dastjerdi, H.; Hadian, J. Antifeedant activity and toxicity of some plant essential oils to Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Plant Protect. Sci. 2014, 50, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Aziza, S.; Rahman, H.A.; Abdel-Aziz, S.; Moawad, S. Natural plant oils and terpenes as protectors for the potato tubers against Phthorimaea operculella (Zeller) infestation by different application methods. Egypt. J. Biol. Pest Control 2014, 24, 265–274. [Google Scholar]
- Osman, S.E.I.; Swidan, M.H.; Kheirallah, D.A.; Nour, F.E. Histological effects of essential oils, their monoterpenoids and insect growth regulators on midgut, integument of larvae and ovaries of khapra beetle, Trogoderma granarium everts. J. Biol. Sci. 2016, 16, 93–101. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Cardenas-Ortega, N.C.; González-Chávez, M.M.; Figueroa-Brito, R.; Flores-Macías, A.; Romo-Asunción, D.; Martínez-González, D.E.; Pérez-Moreno, V.; Ramos-López, M.A. Composition of the essential oil of Salvia ballotiflora (Lamiaceae) and its insecticidal activity. Molecules 2015, 20, 8048–8059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeom, H.J.; Jung, C.S.; Kang, J.S.; Kim, J.; Lee, J.H.; Kim, D.S.; Kim, H.S.; Park, P.S.; Kang, K.S.; Park, I.K. Insecticidal and acetylcholine esterase inhibition activity of Asteraceae plant essential oils and their constituents against adults of the German cockroach (Blattella germanica). J. Agric. Food Chem. 2015, 63, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Gaire, S.; Scharf, M.E.; Gondhalekar, A.D. Toxicity and neurophysiological impacts of plant essential oil components on bed bugs (Cimicidae: Hemiptera). Sci. Rep. 2019, 9, 3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blenau, W.; Rademacher, E.; Baumann, A. Plant essential oils and formamidines as insecticides/acaricides: What are the molecular targets? Apidologie 2012, 43, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Orhan, I.E.; Senol, F.S.; Gülpinar, A.R.; Kartal, M.; Sekeroglu, N.; Deveci, M.; Kan, Y.; Sener, B. Acetylcholinesterase inhibitory and antioxidant properties of Cyclotrichium niveum, Thymus praecox subsp. caucasicus var. caucasicus, Echinacea purpurea and E. pallida. Food Chem. Toxicol. 2009, 47, 1304–1310. [Google Scholar]
- Orhan, I.E.; Senol, F.S.; Ozturk, N.; Celik, S.A.; Pulur, A.; Kan, Y. Phytochemical contents and enzyme inhibitory and antioxidant properties of Anethum graveolens L. (dill) samples cultivated under organic and conventional agricultural conditions. Food Chem. Toxicol. 2013, 59, 96–103. [Google Scholar] [CrossRef]
- Seo, S.M.; Kim, J.; Kang, J.S.; Koh, S.H.; Ahn, Y.J.; Kang, K.S.; Park, I.K. Fumigant toxicity and acetylcholinesterase inhibitory activity of 4 Asteraceae plant essential oils and their constituents against Japanese termite (Reticulitermes speratus Kolbe). Pestic. Biochem. Physiol. 2014, 113, 55–61. [Google Scholar] [CrossRef]
- Re, L.; Barocci, S.; Sonnino, S.; Mencarelli, A.; Vivani, C.; Paolucci, G.; Scarpantonio, A.; Rinaldi, L.; Mosca, E. Linalool modifies the nicotinic receptor-ion channel kinetics at the mouse neuromuscular function. Pharmacol. Res. 2000, 42, 177–181. [Google Scholar] [CrossRef]
- López, M.; Pascual-Villalobos, M. Mode of inhibition of acetylcholinesterase by monoterpenoids and implications for pest control. Ind. Crops Prod. 2010, 3, 284–288. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 052–063. [Google Scholar]
- Enan, E.E. Molecular response of Drosophila melanogaster tyramine receptor cascade to plant essential oils. Insect Biochem. Mol. Bio. 2005, 35, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Ozoe, Y. Molecular signalling, pharmacology, and physiology of octopamine and tyramine receptors as potential insect pest control targets. In Advances in Insect Physiology; Jurenka, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 73–166. [Google Scholar]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system–a review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [Green Version]
- Al-Harbi, N.A.; Al Attar, N.M.; Hikal, D.M.; Mohamed, S.E.; Arafat Abdel Hamed, A.L.; Ibrahim, A.A.; Abdein, M.A. Evaluation of insecticidal effects of plants essential oils extracted from basil, black seeds and lavender against Sitophilus oryzae. Plants 2021, 10, 829. [Google Scholar] [CrossRef]
- Zaker, M. Natural plant products as eco-friendly fungicides for plant diseases control–a review. Agriculturists 2016, 14, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Han, X.B.; Zhao, J.; Cao, J.M.; Zhang, C.S. Essential oil of Chrysanthemum indicum L.: Potential biocontrol agent against plant pathogen Phytophthora nicotianae. Environ. Sci. Pollut. Res. 2019, 26, 7013–7023. [Google Scholar] [CrossRef]
- Regnier, T.; Combrinck, S.; Veldman, W.; Du Plooy, W. Application of essential oils as multi-target fungicides for the control of Geotrichum citri-aurantii and other postharvest pathogens of citrus. Ind. Crops Prod. 2014, 61, 151–159. [Google Scholar] [CrossRef]
- Boubaker, H.; Karim, H.; El Hamdaoui, A.; Msanda, F.; Leach, D.; Bombarda, I.; Vanloot, P.; Abbad, A.; Boudyach, E.; Aoumar, A.A.B. Chemical characterization and antifungal activities of four Thymus species essential oils against postharvest fungal pathogens of citrus. Ind. Crops Prod. 2016, 86, 95–101. [Google Scholar] [CrossRef]
- Hamd, M.; Shazia, I.; Iftikhar, A.; Fateh, F.; Kazmi, M. Identification and characterization of post harvest fungal pathogens of mango from domestic markets of Punjab. Intl. J. Agron. Plant Prod. 2013, 4, 650–658. [Google Scholar]
- Perumal, A.B.; Sellamuthu, P.S.; Nambiar, R.B.; Sadiku, E.R. Antifungal activity of five different essential oils in vapour phase for the control of Colletotrichum gloeosporioides and Lasiodiplodia theobromae in vitro and on mango. Int. J. Food Sci. Technol. 2016, 51, 411–418. [Google Scholar] [CrossRef]
- Cindi, M.D.; Soundy, P.; Romanazzi, G.; Sivakumar, D. Different defense responses and brown rot control in two Prunus persica cultivars to essential oil vapours after storage. Postharvest Biol. Technol. 2016, 119, 9–17. [Google Scholar] [CrossRef]
- Rguez, S.; Djébali, N.; Slimene, I.B.; Abid, G.; Hammemi, M.; Chenenaoui, S.; Bachkouel, S.; Daami-Remadi, M.; Ksouri, R.; Hamrouni-Sellami, I. Cupressus sempervirens essential oils and their major compounds successfully control postharvest grey mould disease of tomato. Ind. Crops Prod. 2018, 123, 135–141. [Google Scholar] [CrossRef]
- Gruľová, D.; Caputo, L.; Elshafie, H.S.; Baranová, B.; De Martino, L.; Sedlák, V.; Gogaľová, Z.; Poráčová, J.; Camele, I.; De Feo, V. Thymol chemotype Origanum vulgare L. essential oil as a potential selective bio-based herbicide on monocot plant species. Molecules 2020, 25, 595. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Mancini, E.; Sakr, S.; De Martino, L.; Mattia, C.A.; De Feo, V.; Camele, I. Antifungal activity of some constituents of Origanum vulgare L. essential oil against postharvest disease of peach fruit. J. Med. Food 2015, 18, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Kordali, S.; Usanmaz Bozhuyuk, A.; Komaki, A.; Ilhan, G.; Ercisli, S. Biological control of Penicillium on lemon fruits by essential oils of Satureja species. Erwerbs-Obstbau 2022, 64, 703–715. [Google Scholar] [CrossRef]
- Amri, I.; Khammassi, M.; Gargouri, S.; Hanana, M.; Jamoussi, B.; Hamrouni, L.; Mabrouk, Y. Tunisian pine essential oils: Chemical composition, herbicidal and antifungal properties. J. Essent. Oil-Bear. Plants 2022, 25, 430–443. [Google Scholar] [CrossRef]
- Parikh, L.; Agindotan, B.O.; Burrows, M.E. Antifungal activity of plant-derived essential oils on pathogens of pulse crops. Plant Dis. 2021, 105, 1692–1701. [Google Scholar] [CrossRef]
- Burt, S.A.; Reinders, R.D. Antibacterial activity of selected plant essential oils against Escherichia coli O157:H7. Lett. Appl. Microbiol. 2003, 36, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Hyldgaard, M.; Mygind, T.; Rikke, L.M. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, J.; Jia, X.; Xin, L.; Zhai, H. Antifungal effects and potential mechanism of essential oils on Collelotrichum gloeosporioides in vitro and in vivo. Molecules 2019, 24, 3386. [Google Scholar] [CrossRef] [Green Version]
- Perina, F.J.; Amaral, D.C.; Fernandes, R.S.; Labory, C.R.; Teixeira, G.A.; Alves, E. Thymus vulgaris essential oil and thymol against Alternaria alternata (Fr.) Keissler: Effects on growth, viability, early infection and cellular mode of action. Pest Manag. Sci. 2015, 71, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Khoury, M.; El Beyrouthy, M.; Ouaini, N.; Eparvier, V.; Stien, D. Hirtellina lobelii DC. essential oil, its constituents, its combination with antimicrobial drugs and its mode of action. Fitoterapia 2019, 133, 130–136. [Google Scholar] [CrossRef]
- Samber, N.; Khan, A.; Varma, A.; Manzoor, N. Synergistic anti-candidal activity and mode of action of Mentha piperita essential oil and its major components. Pharm. Biol. 2015, 53, 1496–1504. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, L.R.; Rosalen, P.L.; Ferreira, G.L.S.; Freires, I.A.; de Carvalho, F.G.; Castellano, L.R.; de Castro, R.D. Antifungal activity, mode of action and anti-biofilm effects of Laurus nobilis Linnaeus essential oil against Candida spp. Arch. Oral Biol. 2017, 73, 179–185. [Google Scholar] [CrossRef]
- Dwivedy, A.K.; Kumar, M.; Upadhyay, N.; Prakash, B.; Dubey, N.K. Plant essential oils against food borne fungi and mycotoxins. Curr. Opin. Food Sci. 2016, 11, 16–21. [Google Scholar] [CrossRef]
- Dwivedy, A.K.; Prakash, B.; Chanotiya, C.S.; Bisht, D.; Dubey, N.K. Chemically characterized Mentha cardiaca L. essential oil as plant based preservative in view of efficacy against biodeteriorating fungi of dry fruits, aflatoxin secretion, lipid peroxidation and safety profile assessment. Food Chem. Toxicol. 2017, 106, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef]
- Rasooli, I.; Rezaei, M.B.; Allameh, A. Growth inhibition and morphological alterations of Aspergillus niger by essential oils from Thymus eriocalyx and Thymus x-porlock. Food Control 2006, 17, 359–364. [Google Scholar] [CrossRef]
- Nogueira, J.H.C.; Goncalez, E.; Galleti, S.R.; Facanali, R.; Marques, M.O.M.; Felicio, J.D. Ageratum conyzoides essential oil as aflatoxin suppressor of Aspergillus flavus. Int. J. Food Microbiol. 2010, 137, 55–60. [Google Scholar] [CrossRef]
- Manso, S.; Cacho-Nerin, F.; Becerril, R.; Nerin, C. Combined analytical and microbiological tools to study the effect on Aspergillus flavus of cinnamon essential oil contained in foodpackaging. Food Control 2013, 30, 370–378. [Google Scholar] [CrossRef]
- Tao, N.; OuYang, Q.; Jia, L. Citral inhibits mycelial growth of Penicillium italicum by a membrane damage mechanism. Food Control 2014, 41, 116–121. [Google Scholar] [CrossRef]
- Knowles, A. New Developments in Crop Protection Product Formulation; T&F Informa UK Ltd.: London, UK, 2005. [Google Scholar]
- Gašić, S.; Tanović, B. Biopesticide formulations, possibility of application and future trends. Pestic. Fitomed. 2013, 28, 97–102. [Google Scholar] [CrossRef]
- Kala, S.; Sogan, N.; Agarwal, A.; Naik, S.; Patanjali, P.; Kumar, J. Biopesticides: Formulations and delivery techniques. In Natural Remedies for Pest Disease and Weed Control; Egbuna, C., Sawicka, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 209–220. [Google Scholar]
- Campolo, O.; Giunti, G.; Russo, A.; Palmeri, V.; Zappalà, L. Essential oils in stored product insect pest control. J. Food Qual. 2018, 2018, 6906105. [Google Scholar] [CrossRef] [Green Version]
- Giunti, G.; Palermo, D.; Laudani, F.; Algeri, G.M.; Campolo, O.; Palmeri, V. Repellence and acute toxicity of a nano-emulsion of sweet orange essential oil toward two major stored grain insect pests. Ind. Crops Prod. 2019, 142, 111869. [Google Scholar] [CrossRef]
- Pavoni, L.; Perinelli, D.R.; Bonacucina, G.; Cespi, M.; Palmieri, G.F. An overview of micro-and nanoemulsions as vehicles for essential oils: Formulation, preparation and stability. Nanomaterials 2020, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.A.; Ferreira-Sa, P.S.; Garcia, M.D., Jr.; Pereira, V.L.P.; Carvalho, J.C.T.; Rocha, L.; Fernandes, C.P.; Souto, R.N.P.; Araujo, R.S.; Botas, G.; et al. Nano-emulsions of the essential oil of Baccharis reticularia and its constituents as eco-friendly repellents against Tribolium castaneum. Ind. Crops Prod. 2021, 162, 113282. [Google Scholar] [CrossRef]
- Chiriac, A.P.; Rusu, A.G.; Nita, L.E.; Chiriac, V.M.; Neamtu, I.; Sandu, A. Polymeric carriers designed for encapsulation of essential oils with biological activity. Pharmaceutics 2021, 13, 631. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Bouquillon, S.; Fauconnier, M.L. Encapsulation of essential oils for the development of biosourced pesticides with controlled release: A review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.I.; Monteiro, M.; Araújo, A.R.; Rodrigues, A.R.O.; Castanheira, E.; Pereira, D.M.; Olim, P.; Fortes, A.G.; Gonçalves, M.S.T. Cytotoxic plant extracts towards insect cells: Bioactivity and nanoencapsulation studies for application as biopesticides. Molecules 2020, 25, 5855. [Google Scholar] [CrossRef]
- Uchida, D.T.; Siqueira, G.F.; Dos Reis, E.M.; Hegeto, F.L.; Medina Neto, A.; Reis, A.V.; Bruschi, M.L.; Villa Nova, M.; Machinski Júnior, M. Design of nanostructured lipid carriers containing Cymbopogon martinii (Palmarosa) essential oil against Aspergillus nomius. Molecules 2021, 26, 4825. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, H.; Saharkhiz, M.J.; Niakousari, M.; Moein, M. Natural herbicide activity of Satureja hortensis L. essential oil nanoemulsion on the seed germination and morphophysiological features of two important weed species. Ecotoxicol. Environ. Saf. 2017, 142, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Campolo, O.; Giunti, G.; Laigle, M.; Michel, T.; Palmeri, V. Essential oil-based nano-emulsions: Effect of different surfactants, sonication and plant species on physicochemical characteristics. Ind. Crops Prod. 2020, 157, 112935. [Google Scholar] [CrossRef]
- Sundari, S.; Singh, A.; Yadava, P. Review of current research advances in microbial and phyto-biopesticides. Int. J. Biotech. Biomed. Sci. 2016, 2, 73–77. [Google Scholar]
- Taban, A.; Saharkhiz, M.J.; Naderi, R. A natural post-emergence herbicide based on essential oil encapsulation by cross-linked biopolymers: Characterization and herbicidal activity. Environ. Sci. Pollut. Res. 2020, 27, 45844–45858. [Google Scholar] [CrossRef]
- Kaur, P.; Gupta, S.; Kaur, K.; Kaur, N.; Kumar, R.; Bhullar, M.S. Nanoemulsion of Foeniculum vulgare essential oil: A propitious striver against weeds of Triticum aestivum. Ind. Crops Prod. 2021, 168, 113601. [Google Scholar] [CrossRef]
- Synowiec, A.; Możdżeń, K.; Krajewska, A.; Landi, M.; Araniti, F. Carum carvi L. essential oil: A promising candidate for botanical herbicide against Echinochloa crus-galli (L.) P. Beauv. in maize cultivation. Ind. Crops Prod. 2019, 140, 111652. [Google Scholar] [CrossRef]
- Gharsan, F.N.; Kamel, W.M.; Alghamdi, T.S.; Alghamdi, A.A.; Althagafi, A.O.; Aljassim, F.J.; Al-Ghamdi, S.N. Toxicity of citronella essential oil and its nanoemulsion against the sawtoothed grain beetle Oryzaephilus surinamensis (Coleoptera: Silvanidae). Ind. Crops Prod. 2022, 184, 115024. [Google Scholar] [CrossRef]
- Giunti, G.; Laudani, F.; Lo Presti, E.; Bacchi, M.; Palmeri, V.; Campolo, O. Contact toxicity and ovideterrent activity of three essential oil-based nano-emulsions against the olive fruit fly Bactrocera oleae. Horticulturae 2022, 8, 240. [Google Scholar] [CrossRef]
- Tortorici, S.; Cimino, C.; Ricupero, M.; Musumeci, T.; Biondi, A.; Siscaro, G.; Carbone, C.; Zappalà, L. Nanostructured lipid carriers of essential oils as potential tools for the sustainable control of insect pests. Ind. Crops Prod. 2022, 181, 114766. [Google Scholar] [CrossRef]
- Sciortino, M.; Scurria, A.; Lino, C.; Pagliaro, M.; D’Agostino, F.; Tortorici, S.; Ricupero, M.; Biondi, A.; Zappala, L.; Ciriminna, R. Silica-microencapsulated orange oil for sustainable pest control. Adv. Sustain. Syst. 2021, 5, 2000280. [Google Scholar] [CrossRef]
- Palermo, D.; Giunti, G.; Laudani, F.; Palmeri, V.; Campolo, O. Essential oil-based nano-biopesticides: Formulation and bioactivity against the confused flour beetle Tribolium confusum. Sustainability 2021, 13, 9746. [Google Scholar] [CrossRef]
- Velho, M.C.; Cossetin, L.F.; Godoi, S.N.D.; Santos, R.C.V.; Gündel, A.; Monteiro, S.G.; Ourique, A.F. Nanobiopesticides: Development and inseticidal activity of nanoemulsions containing lemongrass or eucalyptus oils. Nat Prod Res 2021, 35, 6210–6215. [Google Scholar] [CrossRef] [PubMed]
- Tia, V.E.; Gueu, S.; Cissé, M.; Tuo, Y.; Gnago, A.J.; Konan, E. Bio-insecticidal effects of essential oil nano-emulsion of Lippia multiflora Mold. on major cabbage pests. J. Plant Prot. Res. 2021, 61, 103–109. [Google Scholar]
- Feng, J.; Wang, R.; Chen, Z.; Zhang, S.; Yuan, S.; Cao, H.; Jafari, S.M.; Yang, W. Formulation optimization of D-limonene-loaded nanoemulsions as a natural and efficient biopesticide. Colloids Surf. A Physicochem. Eng. 2020, 596, 124746. [Google Scholar] [CrossRef]
- Miastkowska, M.; Michalczyk, A.; Figacz, K.; Sikora, E. Nanoformulations as a modern form of biofungicide. J. Environ. Health Sci. Eng. 2020, 18, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wu, J.; Zhou, W.; Liu, X.; Zhang, J.; Jia, S.; Meng, Z.; Liu, S.; Lin, R.; Liu, Y. Investigating the multi-target pharmacological mechanism of danhong injection acting on unstable angina by combined network pharmacology and molecular docking. BMC Compl. Med. Ther. 2020, 20, 66. [Google Scholar] [CrossRef] [Green Version]
- Loza-Mejía, M.A.; Salazar, J.R.; Sánchez-Tejeda, J.F. In silico studies on compounds derived from calceolaria: Phenylethanoid glycosides as potential multitarget inhibitors for the development of pesticides. Biomolecules 2018, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Borges, J.C.; Haddi, K.; Oliveira, E.E.; Andrade, B.S.; Nascimento, V.L.; Melo, T.S.; Didonet, J.; Carvalho, J.C.; Cangussu, A.S.; Soares, I.M. Mosquiticidal and repellent potential of formulations containing wood residue extracts of a Neotropical plant, Tabebuia heptaphylla. Ind. Crops Prod. 2019, 129, 424–433. [Google Scholar] [CrossRef]
- Toledo, P.F.; Ferreira, T.P.; Bastos, I.M.; Rezende, S.M.; Jumbo, L.O.V.; Didonet, J.; Andrade, B.S.; Melo, T.S.; Smagghe, G.; Oliveira, E.E. Essential oil from Negramina (Siparuna guianensis) plants controls aphids without impairing survival and predatory abilities of non-target ladybeetles. Environ. Pollut. 2019, 255, 113153. [Google Scholar] [CrossRef]
- Gouda, G.; Bhadra, P. In silico analysis of the peppermint as potential biopesticide. Int. J. Multidiscip. Educ. Res. 2020, 9, 69–78. [Google Scholar]
- Ocampo, A.B.; Mac Kevin, E.B.; Nellas, R.B. The interaction and mechanism of monoterpenes with tyramine receptor (SoTyrR) of rice weevil (Sitophilus oryzae). SN Appl. Sci. 2020, 2, 1592. [Google Scholar] [CrossRef]
- Sierra, I.; Latorre-Estivalis, J.M.; Traverso, L.; Gonzalez, P.V.; Aptekmann, A.; Nadra, A.D.; Masuh, H.; Ons, S. Transcriptomic analysis and molecular docking reveal genes involved in the response of Aedes aegypti larvae to an essential oil extracted from Eucalyptus. PLOS Negl. Trop. Dis. 2021, 15, e0009587. [Google Scholar] [CrossRef]
- Liao, M.; Xiao, J.J.; Zhou, L.J. Insecticidal activity of Melaleuca alternifolia essential oil and RNA-Seq analysis of Sitophilus zeamais transcriptome in response to oil fumigation. PLoS ONE 2016, 11, e0167748. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Zhang, K.; Wei, L.; Wei, G.; Xiong, W.; Lu, Y.; Zhang, Y.; Gao, A.; Li, B. Insecticidal activity of Artemisia vulgaris essential oil and transcriptome analysis of Tribolium castaneum in response to oil exposure. Front. Gen. 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Yang, Q.Q.; Xiao, J.J.; Huang, Y.; Zhou, L.J.; Hua, R.M.; Cao, H.Q. Toxicity of Melaleuca alternifolia essential oil to the mitochondrion and NAD+/NADH dehydrogenase in Tribolium confusum. Peer J. 2018, 6, e5693. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Shao, X.; Xu, J.; Wei, Y.; Xu, F.; Wang, H. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, M.; Yang, Q.; Shi, S.; Xiao, J.; Cao, H. Knockdown of NADPH-cytochrome P450 reductase and CYP6MS1 increases the susceptibility of Sitophilus zeamais to terpinen-4-ol. Pest Biochem. Physiol. 2020, 162, 15–22. [Google Scholar] [CrossRef]
- Muturi, E.J.; Hay, W.T.; Doll, K.M.; Ramirez, J.L.; Selling, G. Insecticidal activity of Commiphora erythraea essential oil and its emulsions against larvae of three mosquito species. J. Med. Entomol. 2020, 13, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Agwunobi, D.O.; Pei, T.; Yang, J.; Wang, X.; Lv, L.; Shen, R.; Yu, Z.; Liu, J. Expression profiles of glutathione S-transferases genes in semi-engorged Haemaphysalis longicornis (Acari: Ixodidae) exposed to Cymbopogon citratus essential oil. Syst. Appl. Acarol. 2020, 25, 918–930. [Google Scholar]
- Agwunobi, D.O.; Zhang, M.; Zhang, X.; Wang, T.; Yu, Z.; Liu, J. Transcriptome profile of Haemaphysalis longicornis (Acari: Ixodidae) exposed to Cymbopogon citratus essential oil and citronellal suggest a cytotoxic mode of action involving mitochondrial Ca2+ overload and depolarization. Pestic. Biochem. Physiol. 2021, 179, 104971. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liao, M.; Yang, Q.; Xiao, J.; Hu, Z.; Zhou, L.; Cao, H. Transcriptome profiling reveals differential gene expression of detoxification enzymes in Sitophilus zeamais responding to terpinen-4-ol fumigation. Pestic. Biochem. Physiol. 2018, 149, 44–53. [Google Scholar] [CrossRef]
- Hegedus, D.; O’grady, M.; Chamankhah, M.; Baldwin, D.; Gleddie, S.; Braun, L.; Erlandson, M. Changes in cysteine protease activity and localization during midgut metamorphosis in the crucifer root maggot (Delia radicum). Insect Biochem. Mol. Biol. 2002, 32, 1585–1596. [Google Scholar] [CrossRef]
- Jampílek, J.; Kráľová, K.; Campos, E.V.; Fraceto, L.F. Bio-based nanoemulsion formulations applicable in agriculture, medicine, and food industry. In Nanobiotechnology in Bioformulations; Prasad, R., Kumar, V., Kumar, M., Choudhary, D.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 33–84. [Google Scholar]
- Zhang, Z.; Tan, Y.; McClements, D.J. Investigate the adverse effects of foliarly applied antimicrobial nanoemulsion (carvacrol) on spinach. LWT 2021, 141, 110936. [Google Scholar] [CrossRef]
- Moradzadeh, S.; Moghaddam, S.S.; Rahimi, A.; Pourakbar, L.; Sayyed, R.Z. Combined bio-chemical fertilizers ameliorate agro-biochemical attributes of black cumin (Nigella sativa L.). Sci. Rep. 2021, 11, 11399. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Roscigno, G.; Pane, C.; Celano, G.; Di Matteo, M.; Mainente, M.; Vuotto, A.; Menchenrini, T.; Esposito, T.; Vitti, A.; et al. Essential oils and quality composts sourced by recycling vegetable residues from the aromatic plant supply chain. Ind. Crops Prod. 2021, 162, 113255. [Google Scholar] [CrossRef]
- Cid-Pérez, T.S.; Ávila-Sosa, R.; Ochoa-Velasco, C.E.; Rivera-Chavira, B.E.; Nevárez-Moorillón, G.V. Antioxidant and antimicrobial activity of Mexican oregano (Poliomintha longiflora) essential oil, hydrosol and extracts from waste solid residues. Plants 2019, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Wang, W.; Bao, L.; Wang, S.; Cao, X. Synthesis and evaluation of essential oil-derived β-methoxyacrylate derivatives as high potential fungicides. Molecules 2017, 22, 763. [Google Scholar] [CrossRef] [Green Version]
- Damalas, C.A.; Koutroubas, S.D. Botanical pesticides for eco-friendly pest management: Drawbacks and limitations. In Pesticides in Crop Production: Physiological and Biochemical Action; Srivastava, P.K., Singh, V.P., Singh, A., Tripathi, D.K., Singh, S., Prasad, S.M., Chauhan, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 181–193. [Google Scholar]
- Essiedu, J.A.; Adepoju, F.O.; Ivantsova, M.N. Benefits and limitations in using biopesticides: A review. AIP Conf. Proc. 2020, 2313, 080002. [Google Scholar]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Nyirenda, S.P.; Mvumi, B.; Sola, P.; Kamanula, J.F.; Sileshi, G.; Belmain, S.R. Pesticidal plants: A viable alternative insect pest management approach for resource-poor farming in Africa. In Biopesticides in Environment and Food Security: Issues and Strategies; Koul, O., Dhaliwal, G.S., Khokhar, S., Singh, R., Eds.; Scientific Publishers: Jodhpur, India, 2012; pp. 212–238. [Google Scholar]
- Brown, M.B.; Brown, C.M.B.; Nepomuceno, R.A. Regulatory requirements and registration of biopesticides in the Philippines. In Agriculturally Important Microorganisms; Singh, H., Sarma, B., Keswani, C., Eds.; Springer: Singapore, 2016; pp. 183–195. [Google Scholar]
- Isman, M.B.; Tak, J. Inhibition of acetylcholinesterase by essential oils and monoterpenoids: A relevant mode of action for insecticidal essential oils. Biopestic. Int. 2017, 13, 71–78. [Google Scholar]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Tangtrakulwanich, K.; Reddy, G.V. Development of insect resistance to plant biopesticides: An overview. In Advances in Plant Biopesticides; Singh, D., Ed.; Springer: Singapore, 2014; pp. 47–62. [Google Scholar]
- Sarkic, A.; Stappen, I. Essential oils and their single compounds in cosmetics–a critical review. Cosmetics 2018, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Cimino, C.; Maurel, O.M.; Musumeci, T.; Bonaccorso, A.; Drago, F.; Souto, E.M.B.; Pignatello, R.; Carbone, C. Essential oils: Pharmaceutical applications and encapsulation strategies into lipid-based delivery systems. Pharmaceutics 2021, 13, 327. [Google Scholar] [CrossRef] [PubMed]
- Padole, V.N.; Sarade, S.; Rathod, S.; More, S.; Mendhi, S. Multivalent role of essential oil in cosmetics: A review. Int. J. Pharm. Sci. Rev. Res. 2022, 73, 97–105. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic effects and mechanism of action of essential oils and terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as sources for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef]







| Type of Biopesticide | Country | PEO/Active Component | Name of Product |
|---|---|---|---|
| Biofungicide | Europe | Clove oil | BIOXEDA |
| Biofungicide + Bioinsecticide | Europe | Sweet orange oil | LIMOCIDE |
| Biofungicide + Bioinsecticide | Europe | Sweet orange oil | OROCIDE |
| Biofungicide + Bioinsecticide | Europe | Sweet orange oil | PREV-AM |
| Bioherbicide | USA | d-Limonene | GreenMatch |
| Bioherbicide | USA | Clove oil | Matratec |
| Bioherbicide | USA | 45% Clove oil + 45% cinnamon oil | WeedZap |
| Bioherbicide | USA | Lemongrass oil | GreenMatch EX |
| Bioherbicide | USA | d-Limonene | Avenger Weed Killer |
| Bioherbicide | USA | Eugenol | Weed Slayer |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, I.; Singh, R.; Muthusamy, S.; Sharma, M.; Grewal, K.; Singh, H.P.; Batish, D.R. Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints. Plants 2023, 12, 2916. https://doi.org/10.3390/plants12162916
Gupta I, Singh R, Muthusamy S, Sharma M, Grewal K, Singh HP, Batish DR. Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints. Plants. 2023; 12(16):2916. https://doi.org/10.3390/plants12162916
Chicago/Turabian StyleGupta, Ipsa, Rishikesh Singh, Suganthi Muthusamy, Mansi Sharma, Kamaljit Grewal, Harminder Pal Singh, and Daizy R. Batish. 2023. "Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints" Plants 12, no. 16: 2916. https://doi.org/10.3390/plants12162916