
Spatial Distribution of Pinus koraiensis Trees and Community-Level Spatial Associations in Broad-Leaved Korean Pine Mixed Forests in Northeastern China

Abstract
:1. Introduction
2. Results
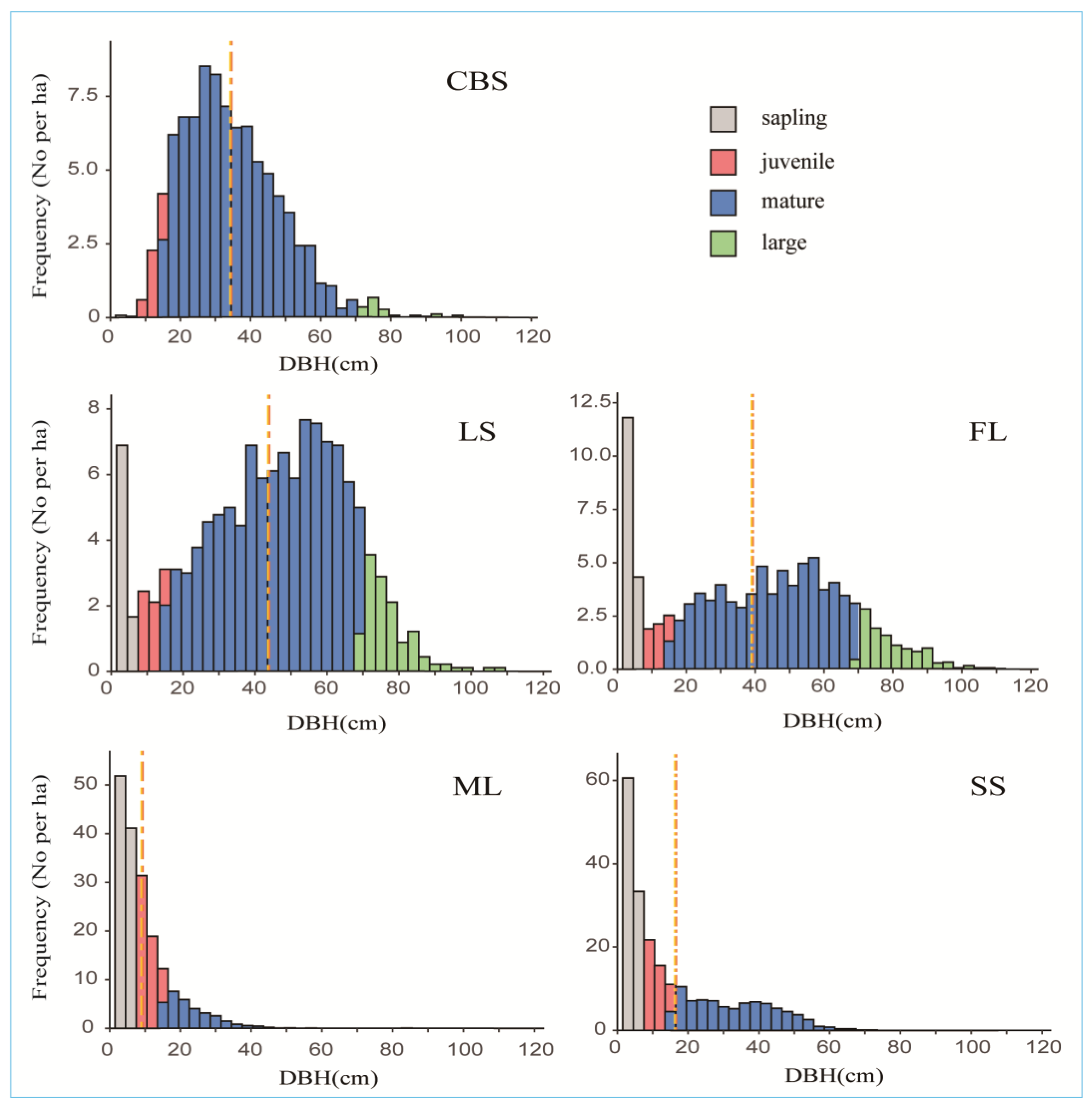
2.1. Population Structure of Korean Pine Trees
2.2. Spatial Distribution of and Spatial Association among Life Stages of KP Trees
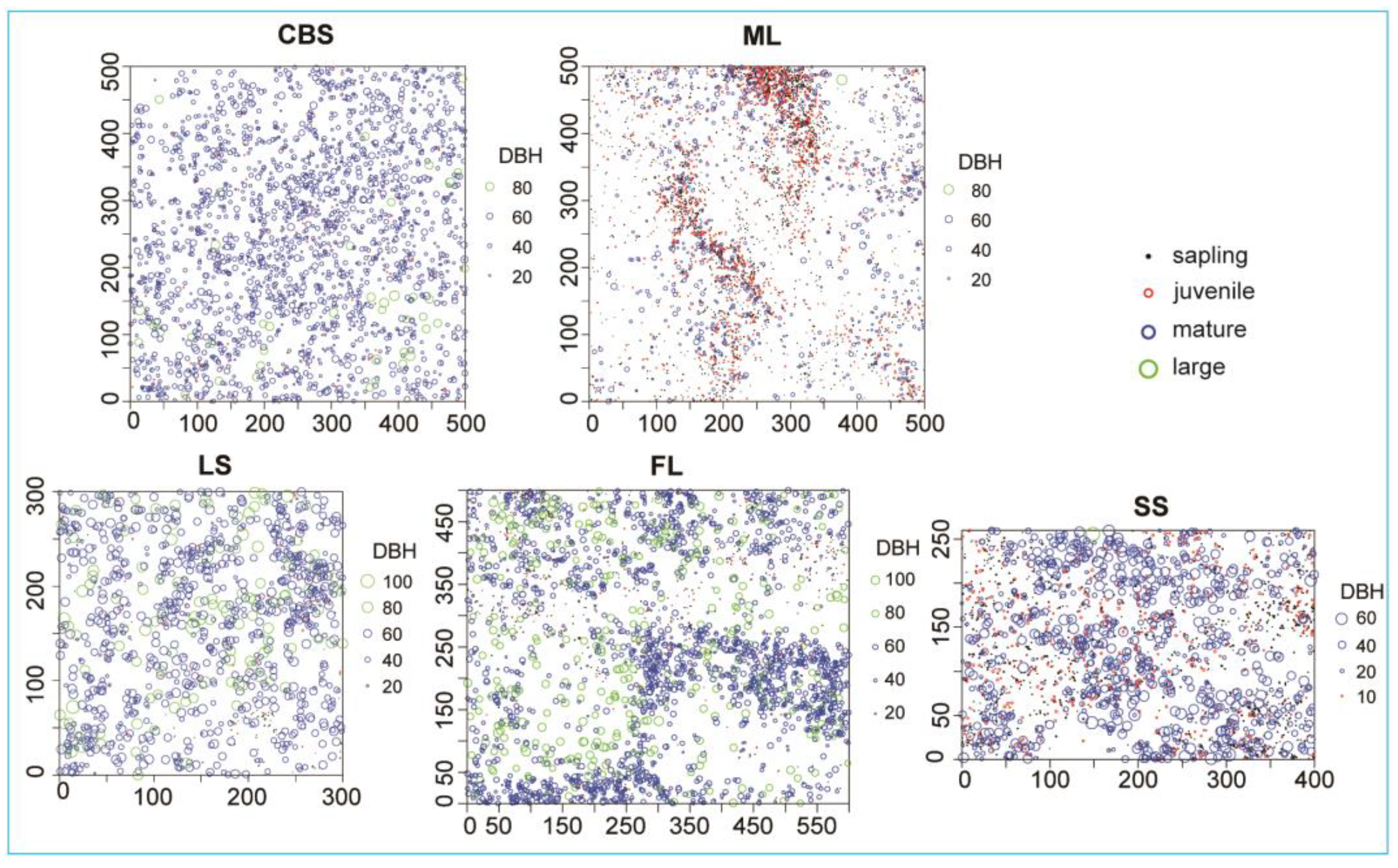
2.2.1. Point Pattern Analysis of KP Trees
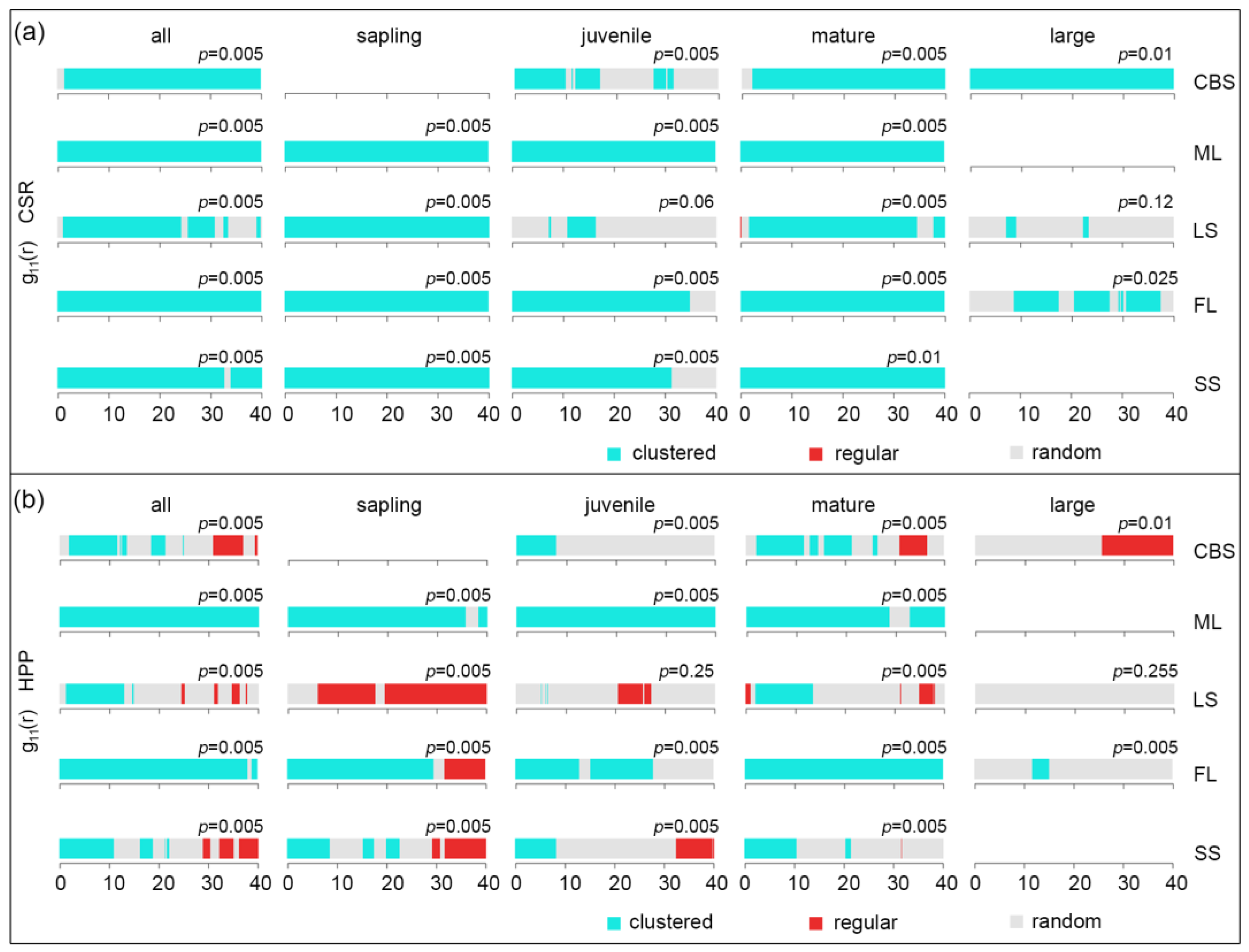
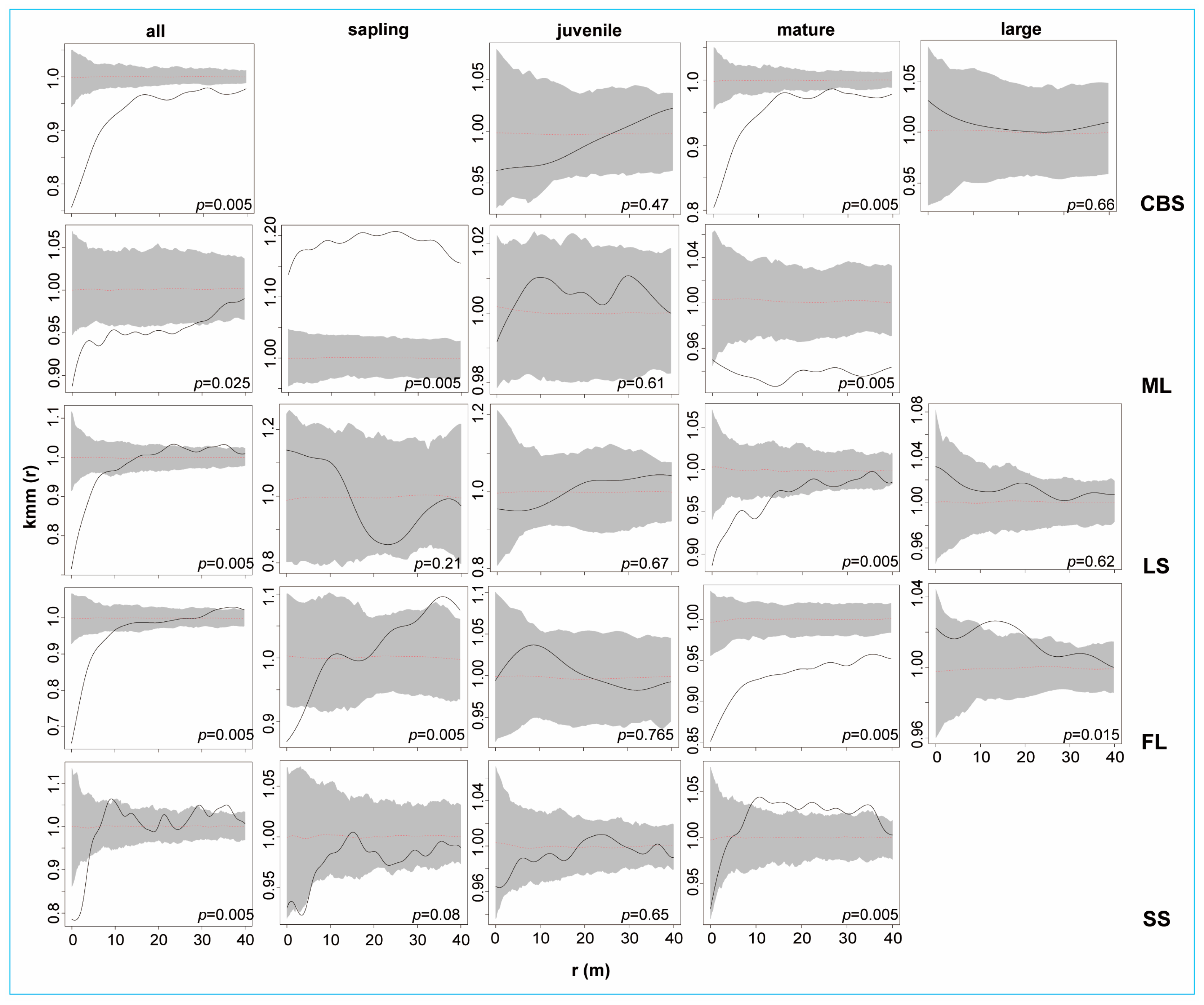
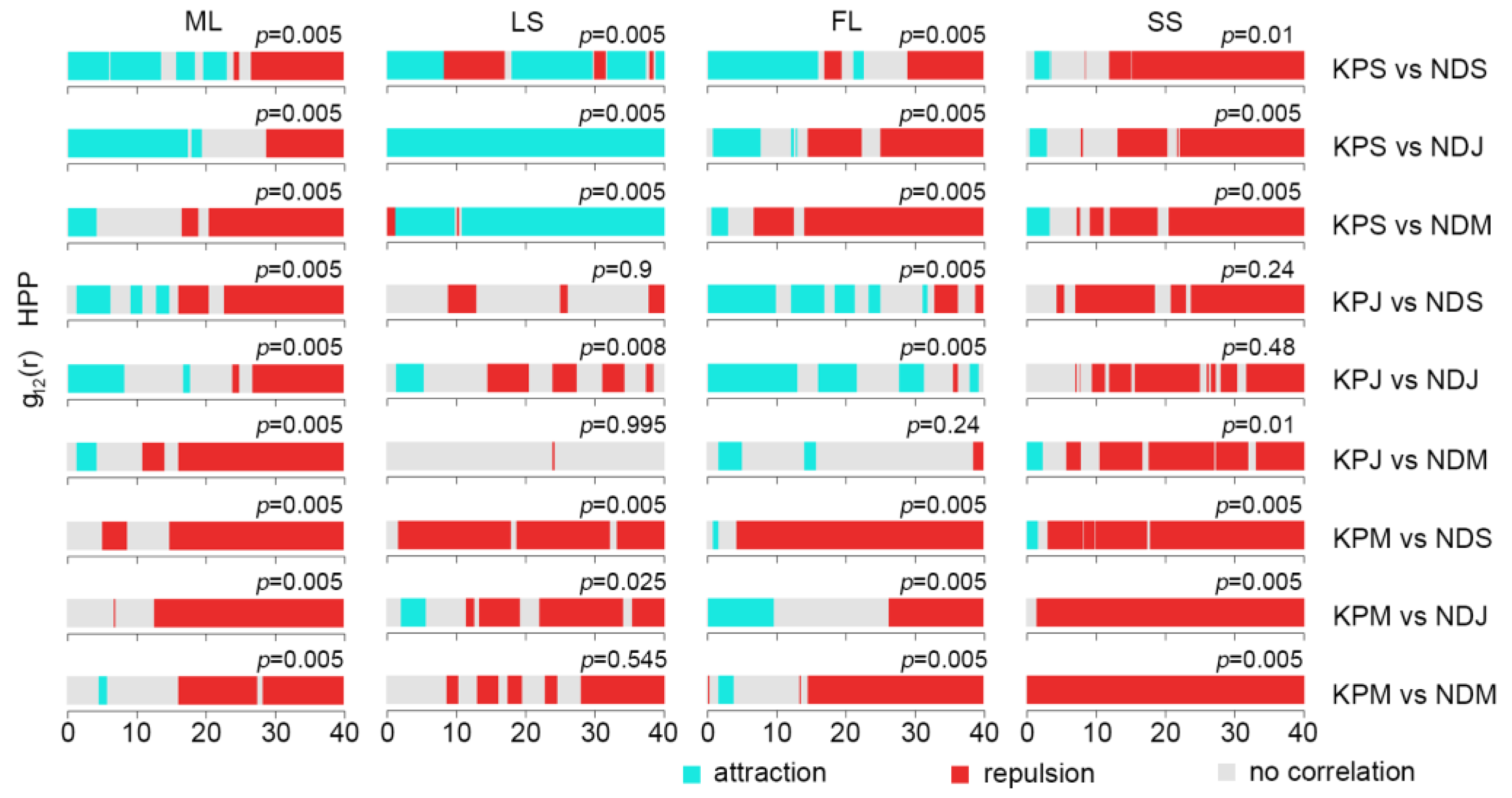
2.2.2. Intraspecific Association of KP Trees
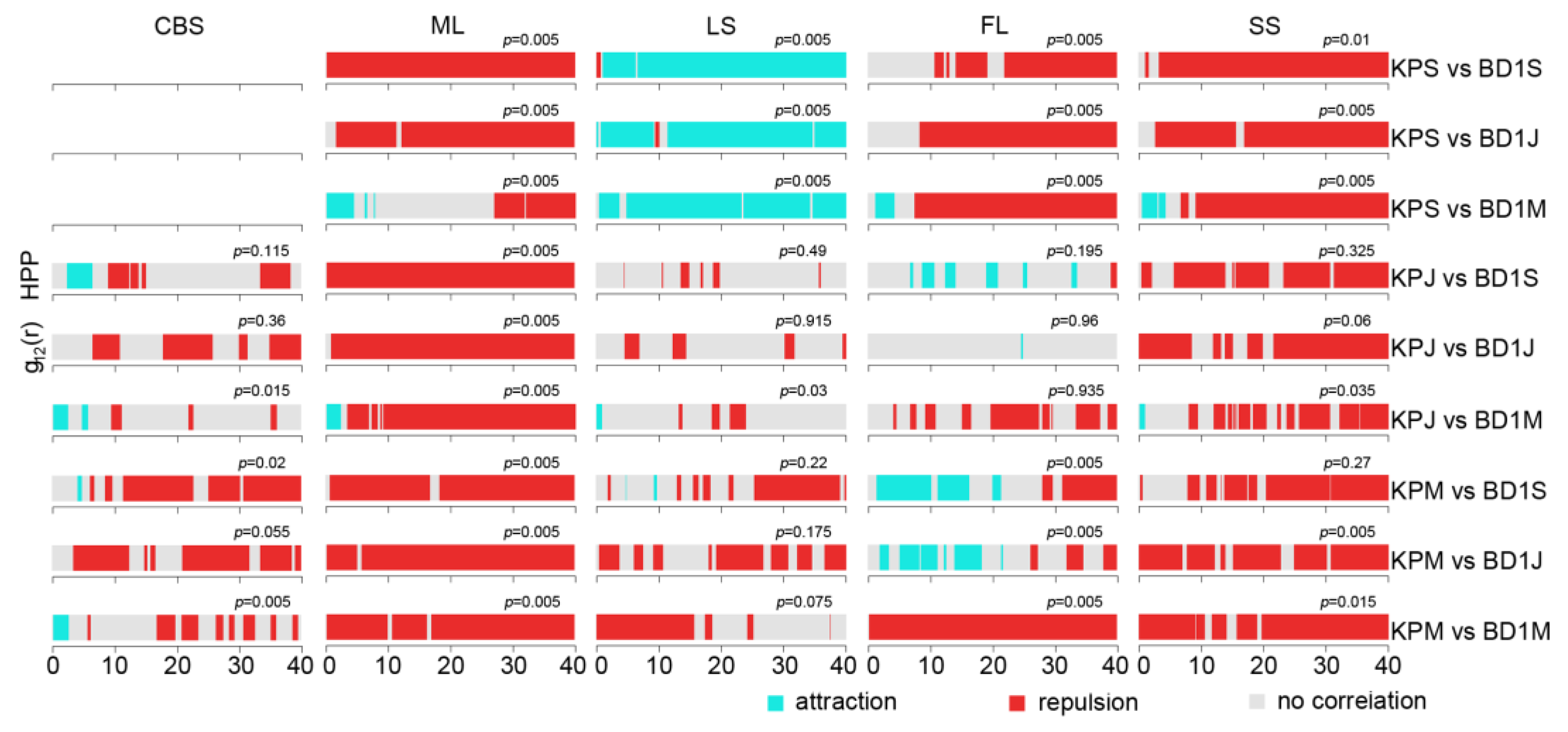
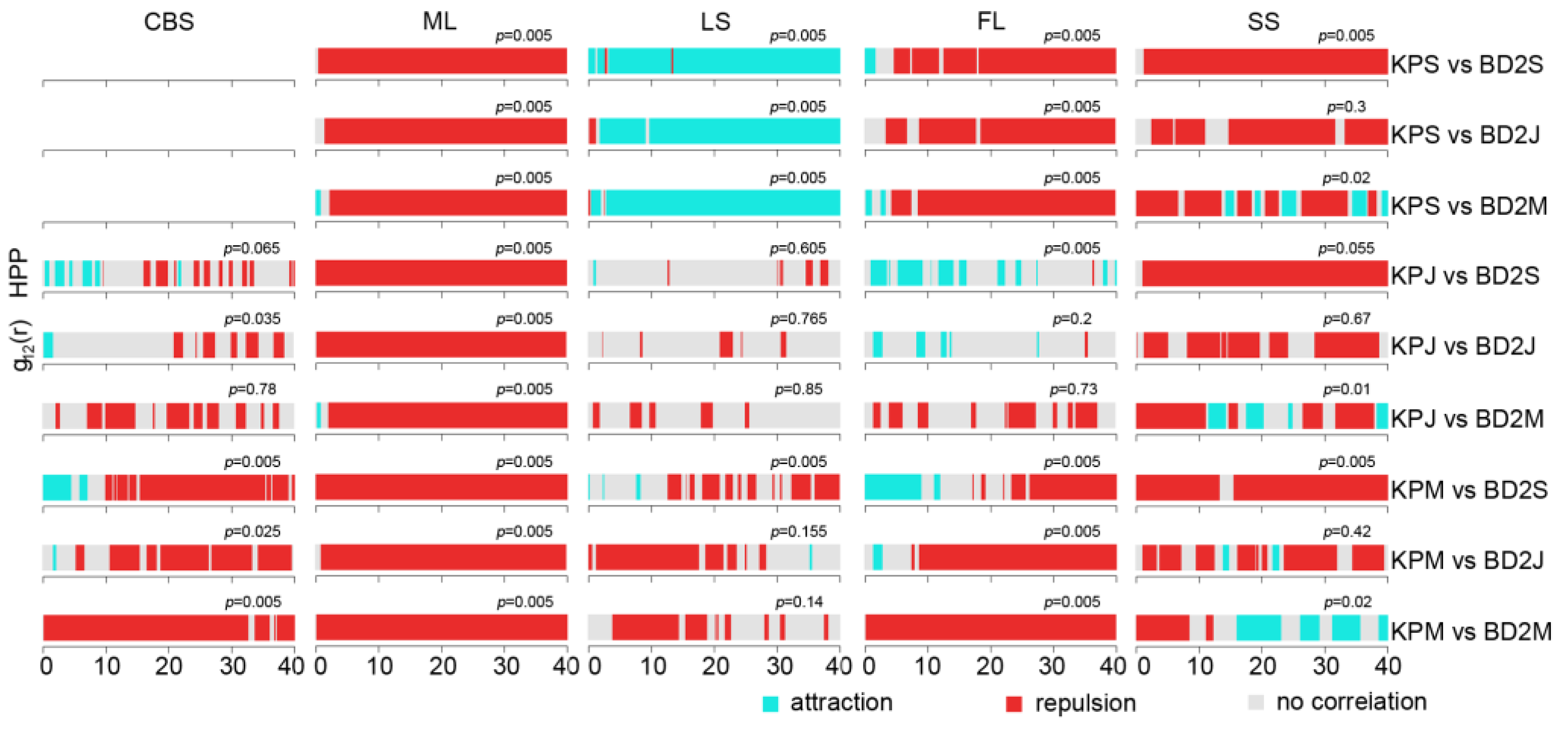
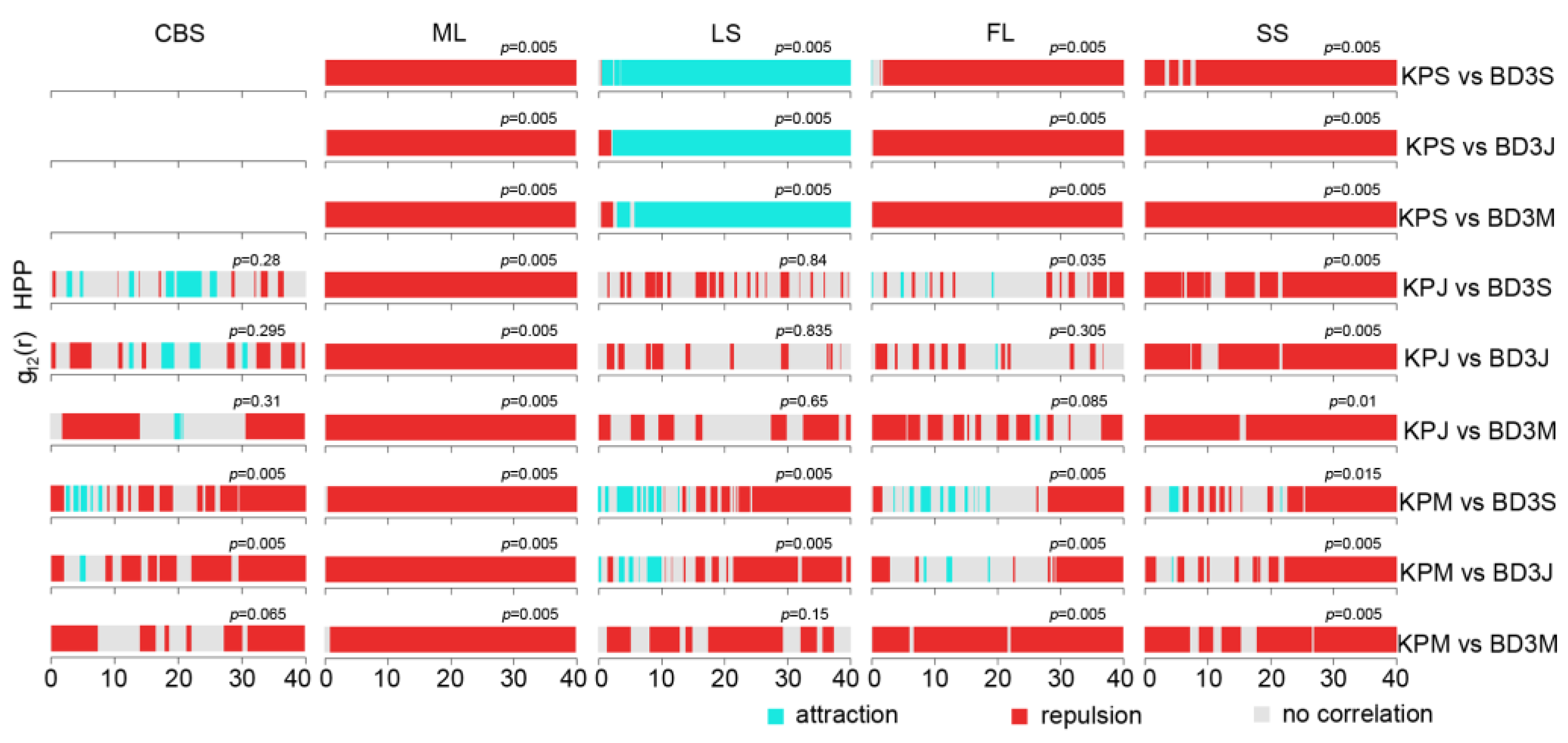
2.3. Interspecific Association between Korean Pine and Other Tree Species
3. Discussion
3.1. Population Dynamics and Spatial Patterns of KP Trees in Northeastern China
3.2. The Intraspecific Association of KP Trees and Interspecific Association with Other Tree Species at the Community Level
4. Materials and Methods
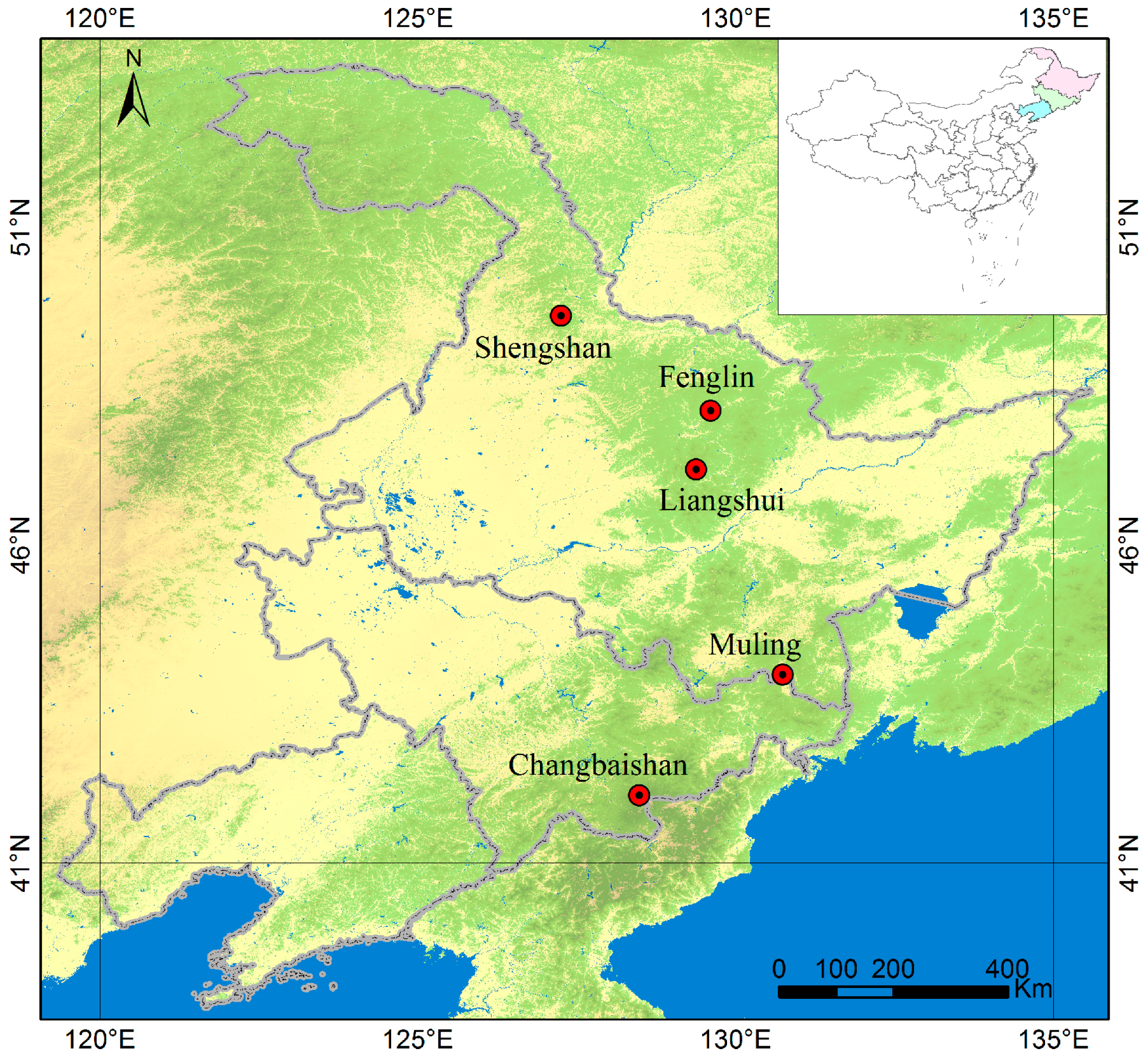
4.1. Study Area
4.2. Field Data Collection
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Omelko, A.; Ukhvatkina, O.; Zhmerenetsky, A. Disturbance history and natural regeneration of an old-growth Korean pine-broadleaved forest in the Sikhote-Alin mountain range, Southeastern Russia. For. Ecol. Manag. 2016, 360, 221–234. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.T.; Wei, J.T.; Li, Y.; Tigabu, M.; Zhao, X.Y. Genetic Improvement of Pinus koraiensis in China: Current Situation and Future Prospects. Forests 2020, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Stone, R. Ecology. A threatened nature reserve breaks down Asian borders. Science 2006, 313, 1379–1380. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yan, H. A study of the impacts of climate change on the geographic distribution of Pinus koraiensis in China. Environ. Int. 2001, 27, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Blanco, J.A.; Wei, X.; Kang, X.; Wang, W.; Guo, Y. Determining suitable selection cutting intensities based on long-term observations on aboveground forest carbon, growth, and stand structure in Changbai Mountain, Northeast China. Scand. J. For. Res. 2014, 29, 436–454. [Google Scholar] [CrossRef]
- Wu, C.; Venevsky, S.; Sitch, S.; Yang, Y.; Wang, M.; Wang, L.; Gao, Y. Present-day and future contribution of climate and fires to vegetation composition in the boreal forest of China. Ecosphere 2017, 8, e1917. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.; Han, S.; Wang, H. Genetic diversity and genetic differentiation of natural Pinus koraiensis population. J. For. Res. 2006, 17, 21–24. [Google Scholar] [CrossRef]
- Shi, B.; Jin, G. Variability of soil respiration at different spatial scales in temperate forests. Biol. Fert. Soils 2016, 52, 561–571. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, J.; Li, M.; Zhang, G.; Yan, Q. Different light acclimation strategies of two coexisting tree species seedlings in a temperate secondary forest along five natural light levels. For. Ecol. Manag. 2013, 306, 234–242. [Google Scholar] [CrossRef]
- Wang, J.; Geoff Wang, G.; Li, R.; Sun, Y.; Yu, L.; Zhu, J.; Yan, Q. Cascading effect of source limitation on the granivore-mediated seed dispersal of Korean pine (Pinus koraiensis) in secondary forest ecosystems. Ecol. Process. 2022, 11, 1. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Li, J.M.; Chang, S.L.; Li, X.; Lu, J. Spatial distribution pattern of Picea schrenkiana population in the Middle Tianshan Mountains and the relationship with topographic attributes. J. Arid. Land 2012, 4, 457–468. [Google Scholar] [CrossRef]
- Gadow, K.V.; Zhang, C.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. Forest Structure and Diversity. In Continuous Cover Forestry; Springer: Dordrecht, The Netherlands, 2012; pp. 28–83. [Google Scholar]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species associations in a heterogeneous Sri lankan dipterocarp forest. Am. Nat. 2007, 170, E77–E95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Wang, W.; Bai, Z.; Guo, Z.; Ren, W.; Huang, J.; Xu, Y.; Yao, J.; Ding, Y.; Zang, R. Competition and facilitation co-regulate the spatial patterns of boreal tree species in Kanas of Xinjiang, northwest China. For. Ecol. Manag. 2020, 467, 118167. [Google Scholar] [CrossRef]
- Punchi-Manage, R.; Wiegand, K.; Wiegand, T.; Getzin, S.; Huth, A.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Neighborhood diversity of large trees shows independent species patterns in a mixed dipterocarp forest in Sri Lanka. Ecology 2015, 96, 1823–1834. [Google Scholar] [CrossRef]
- Shen, G.; Wiegand, T.; Mi, X.; He, F. Quantifying spatial phylogenetic structures of fully stem-mapped plant communities. Methods Ecol. Evol. 2013, 4, 1132–1141. [Google Scholar] [CrossRef]
- Omelko, A.; Ukhvatkina, O.; Zhmerenetsky, A.; Sibirina, L.; Petrenko, T.; Bobrovsky, M. From young to adult trees: How spatial patterns of plants with different life strategies change during age development in an old-growth Korean pine-broadleaved forest. For. Ecol. Manag. 2018, 411, 46–66. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial Patterns in the Distribution of Tropical Tree Species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Feng, Y.; Guan, W.; Cao, X.; Li, Z.; Ding, J. Using unmanned aerial vehicles to quantify spatial patterns of dominant vegetation along an elevation gradient in the typical Gobi region in Xinjiang, Northwest China. Glob. Ecol. Conserv. 2021, 27, e1571. [Google Scholar] [CrossRef]
- Ellison, A.; Buckley, H.; Case, B.; Cardenas, D.; Duque, J.; Lutz, J.A.; Myers, J.A.; Orwig, D.A.; Zimmerman, J.K. Species Diversity Associated with Foundation Species in Temperate and Tropical Forests. Forests 2019, 10, 128. [Google Scholar] [CrossRef] [Green Version]
- Carrer, M.; Castagneri, D.; Popa, I.; Pividori, M.; Lingua, E. Tree spatial patterns and stand attributes in temperate forests: The importance of plot size, sampling design, and null model. For. Ecol. Manag. 2018, 407, 125–134. [Google Scholar] [CrossRef]
- Velázquez, E.; Martínez, I.; Getzin, S.; Moloney, K.A.; Wiegand, T. An evaluation of the state of spatial point pattern analysis in ecology. Ecography 2016, 39, 1042–1055. [Google Scholar] [CrossRef]
- Petritan, I.C.; Marzano, R.; Petritan, A.M.; Lingua, E. Overstory succession in a mixed Quercus petraea–Fagus sylvatica old growth forest revealed through the spatial pattern of competition and mortality. For. Ecol. Manag. 2014, 326, 9–17. [Google Scholar] [CrossRef]
- Sterner, R.W.; Ribic, C.A.; Schatz, G.E. Testing for life historical changes in spatial patterns of four tropical tree species. J. Ecol. 1986, 74, 621–633. [Google Scholar] [CrossRef]
- Gray, L.; He, F. Spatial point-pattern analysis for detecting density-dependent competition in a boreal chronosequence of Alberta. For. Ecol. Manag. 2009, 259, 98–106. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Ben-Said, M. Spatial point-pattern analysis as a powerful tool in identifying pattern-process relationships in plant ecology: An updated review. Ecol. Process. 2021, 10, 56. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology, 1st ed.; Chapman and Hall/CRC: Portsmouth, UK, 2014. [Google Scholar]
- Zhao, C.; Zhao, W.; Jin, M.; Zhou, J.; Ta, F.; Wang, L.; Mou, W.; Lei, L.; Liu, J.; Du, J.; et al. Spatial patterns of Picea crassifolia driven by environmental heterogeneity and intraspecific interactions. J. For. Res. 2023, 34, 949–962. [Google Scholar] [CrossRef]
- He, C.; Jia, S.; Luo, Y.; Hao, Z.; Yin, Q. Spatial Distribution and Species Association of Dominant Tree Species in Huangguan Plot of Qinling Mountains, China. Forests 2022, 13, 866. [Google Scholar] [CrossRef]
- Shen, G.; Yu, M.; Hu, X.S.; Mi, X.; Ren, H.; Sun, I.-F.; Ma, K. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology 2009, 90, 3033–3041. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Bi, L.; Song, G.; Wang, Q.; Jin, G. Species-habitat associations in an old-growth temperate forest in northeastern China. BMC Ecol. 2018, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.; Wiegand, T.; González-Taboada, F.; Obeso, J.R. Spatial associations among tree species in a temperate forest community in North-western Spain. For. Ecol. Manag. 2010, 260, 456–465. [Google Scholar] [CrossRef]
- Perea, A.J.; Wiegand, T.; Garrido, J.L.; Rey, P.J.; Alcántara, J.M. Legacy effects of seed dispersal mechanisms shape the spatial interaction network of plant species in Mediterranean forests. J. Ecol. 2021, 109, 3670–3684. [Google Scholar] [CrossRef]
- Bagchi, R.; Swinfield, T.; Gallery, R.E.; Lewis, O.T.; Gripenberg, S.; Narayan, L.; Freckleton, R.P. Testing the Janzen-Connell mechanism: Pathogens cause overcompensating density dependence in a tropical tree. Ecol. Lett. 2010, 13, 1262–1269. [Google Scholar] [CrossRef]
- Fedriani, J.M.; Wiegand, T.; Calvo, G.; Suárez-Esteban, A.; Jácome, M.; Żywiec, M.; Delibes, M. Unravelling conflicting density- and distance-dependent effects on plant reproduction using a spatially explicit approach. J. Ecol. 2015, 103, 1344–1353. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.J.; Wiegand, T.; Comita, L.S. Distance-dependent seedling mortality and long-term spacing dynamics in a neotropical forest community. Ecol. Lett. 2017, 20, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Piao, T.; Comita, L.S.; Jin, G.; Kim, J.H. Density dependence across multiple life stages in a temperate old-growth forest of northeast China. Oecologia 2013, 172, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Pérez, J.; Imbert, B.; Peralta, J. Environment and density-dependency explain the fine-scale aggregation of tree recruits before and after thinning in a mixed forest of Southern Europe. PeerJ 2022, 10, e13892. [Google Scholar] [CrossRef]
- Johnson, D.J.; Bourg, N.A.; Howe, R.; McShea, W.J.; Wolf, A.; Clay, K. Conspecific negative density-dependent mortality and the structure of temperate forests. Ecology 2014, 95, 2493–2503. [Google Scholar] [CrossRef]
- Orwig, D.A.; Aylward, J.A.; Buckley, H.L.; Case, B.S.; Ellison, A.M. Land-use history impacts spatial patterns and composition of woody plant species across a 35-hectare temperate forest plot. PeerJ 2022, 10, e12693. [Google Scholar] [CrossRef]
- Hao, Z.; Zhang, J.; Song, B.; Ye, J.; Li, B. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Zhang, J.; Song, B.; Li, B.; Ye, J.; Wang, X.-G.; Hao, Z.-Q. Spatial patterns and associations of six congeneric species in an old-growth temperate forest. Acta Oecol. 2010, 36, 29–38. [Google Scholar] [CrossRef]
- Wang, X.; Wiegand, T.; Hao, Z.; Li, B.; Ye, J.; Lin, F. Species associations in an old-growth temperate forest in north-eastern China. J. Ecol. 2010, 98, 674–686. [Google Scholar] [CrossRef]
- Gong, Z.; Kang, X.; Gu, L. Quantitative division of succession and spatial patterns among different stand developmental stages in Changbai Mountains. J. Mt. Sci.-Engl. 2019, 16, 2063–2078. [Google Scholar] [CrossRef]
- Liu, Y.; Li, F.; Jin, G. Spatial patterns and associations of four species in an old-growth temperate forest. J. Plant Interact. 2014, 9, 745–753. [Google Scholar] [CrossRef]
- Li, J.; Li, J. Regeneration and restoration of broad-leaved Korean pine forests in Lesser Xing′an Mountains of Northeast China. Acta Ecol. Sin. 2003, 23, 1268–1277. [Google Scholar] [CrossRef]
- Zhou, Q.; Shi, H.; Shu, X.; Xie, F.; Zhang, K.; Zhang, Q.; Dang, H. Spatial distribution and interspecific associations in a deciduous broad-leaved forest in north-central China. J. Veg. Sci. 2019, 30, 1153–1163. [Google Scholar] [CrossRef]
- Gu, L.; O’Hara, K.L.; Li, W.Z.; Gong, Z. Spatial patterns and interspecific associations among trees at different stand development stages in the natural secondary forests on the Loess Plateau, China. Ecol. Evol. 2019, 9, 6410–6421. [Google Scholar] [CrossRef]
- Boyden, S.; Binkley, D.; Shepperd, W. Spatial and temporal patterns in structure, regeneration, and mortality of an old-growth ponderosa pine forest in the Colorado Front Range. For. Ecol. Manag. 2005, 219, 43–55. [Google Scholar] [CrossRef]
- Dai, L.; Qi, L.; Wang, Q.; Su, D.; Yu, D.; Wang, Y.; Ye, Y.; Jiang, S.; Zhao, W. Changes in forest structure and composition on Changbai Mountain in Northeast China. Ann. For. Sci. 2011, 68, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Zhu, C.; Lu, D.; Wang, G.G.; Zheng, X.; Cao, J.; Zhang, J. Regeneration and succession: A 50-year gap dynamic in temperate secondary forests, Northeast China. For. Ecol. Manag. 2021, 484, 118943. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, J.; Sun, O.J.; Yan, Q. Photosynthetic and growth responses of Pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved Korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Yin, J.; Lin, F.; De Lombaerde, E.; Mao, Z.; Liu, S.; Ye, J.; Fang, S.; Wang, X. The effects of light, conspecific density and soil fungi on seedling growth of temperate tree species. For. Ecol. Manag. 2023, 529, 120683. [Google Scholar] [CrossRef]
- Wang, X.; Pederson, N.; Chen, Z.; Lawton, K.; Zhu, C.; Han, S. Recent rising temperatures drive younger and southern Korean pine growth decline. Sci. Total Environ. 2019, 649, 1105–1116. [Google Scholar] [CrossRef]
- Manzello, S.L.; Maranghides, A.; Shields, J.R.; Mell, W.E.; Hayashi, Y.; Nii, D. Mass and size distribution of firebrands generated from burning Korean pine (Pinus koraiensis) trees. Fire Mater. 2009, 33, 21–31. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, Q.; Xu, Z.; Du, W.; Yu, J.; Meng, S.; Zhou, H.; Qin, L.; Shah, S. How can the shade intolerant Korean pine survive under dense deciduous canopy? For. Ecol. Manag. 2020, 457, 117735. [Google Scholar] [CrossRef]
- Qi, L.; Yang, J.; Yu, D.; Dai, L.; Contrereas, M. Responses of regeneration and species coexistence to single-tree selective logging for a temperate mixed forest in eastern Eurasia. Ann. For. Sci. 2016, 73, 449–460. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, J.; Yu, L.; Yan, Q.L.; Wang, K. Photosynthetic characteristics of Pinus koraiensis seedlings under different light regimes. Chin. J. Ecol. 2009, 28, 850–857. [Google Scholar]
- Yang, H.; Wu, Y. Tree composition, age structure and regeneration strategy of the mixed broadleaf Korean pine forest in Changbaishan biosphere reserve, China. Sci. Silvae Sin. 1988, 24, 18–27. [Google Scholar]
- Beyns, R.; Bauman, D.; Drouet, T. Fine-scale tree spatial patterns are shaped by dispersal limitation which correlates with functional traits in a natural temperate forest. J. Veg. Sci. 2021, 32, e13070. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Dohn, J.; Augustine, D.J.; Hanan, N.P.; Ratnam, J.; Sankaran, M. Spatial vegetation patterns and neighborhood competition among woody plants in an East African savanna. Ecology 2017, 98, 478–488. [Google Scholar] [CrossRef] [Green Version]
- Salas, C.; LeMay, V.; Núñez, P.; Pacheco, P.; Espinosa, A. Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile. For. Ecol. Manag. 2006, 231, 38–46. [Google Scholar] [CrossRef]
- Stoyan, D.; Penttinen, A. Recent applications of point process methods in forestry statistics. Stat. Sci. 2000, 15, 61–78. [Google Scholar]
- Wang, X.; Ye, J.; Li, B.; Zhang, J.; Lin, F.; Hao, Z. Spatial distributions of species in an old-growth temperate forest, northeastern China. Can. J. For. Res. 2010, 40, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hao, Z.; Song, B.; Li, B.; Wang, X.; Ye, J. Fine-scale species co-occurrence patterns in an old-growth temperate forest. For. Ecol. Manag. 2009, 257, 2115–2120. [Google Scholar] [CrossRef]
- Cheng, X.; Han, H.; Kang, F.; Song, Y.; Liu, K. Point pattern analysis of different life stages of Quercus liaotungensis in Lingkong Mountain, Shanxi Province, China. J. Plant Interact. 2014, 9, 233–240. [Google Scholar] [CrossRef]
- Ding, S. Spatial distribution patterns and spatial associations of Pinus koraiensis in broad-leaved Korean pine mixed forest. J. Shandong For. Sci. Technol. 2019, 49, 25–28. [Google Scholar] [CrossRef]
- Chen, B.; Wang, K.; Ni, R.; Cheng, Y. Composition and spatial pattern of tree seedlings in a coniferous and broadleaved mixed forest in Changbai Mountain of northeastern China. J. Beijing For. Univ. 2018, 40, 68–75. [Google Scholar]
- Martínez, I.; González Taboada, F.; Wiegand, T.; Obeso, J.R. Spatial patterns of seedling-adult associations in a temperate forest community. For. Ecol. Manag. 2013, 296, 74–80. [Google Scholar] [CrossRef]
- Dong, X.; Du, X.; Sun, Z.; Gu, H.Y.; Chen, X.W. Spatial pattern and intraspecific association of natural Korean pine population under the influence of habitat gradient. Acta Ecol. Sin. 2020, 40, 5239–5246. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, X.; Liu, X.; Gadow, K.V. Spatial distribution and spatial association of dominant tree species in Korean pine broadleaved old-growth forests in Changbai mountain. Balt. For. 2010, 16, 66–75. [Google Scholar]
- Zhao, H.; Kang, X.; Guo, Z.; Yang, H.; Xu, M. Species Interactions in Spruce-Fir Mixed Stands and Implications for Enrichment Planting in the Changbai Mountains, China. Mt. Res. Dev. 2012, 32, 187–196. [Google Scholar] [CrossRef]
- Brown, A.J.; Payne, C.J.; White, P.S.; Peet, R.K. Shade tolerance and mycorrhizal type may influence sapling susceptibility to conspecific negative density dependence. J. Ecol. 2020, 108, 325–336. [Google Scholar] [CrossRef]
- Kenkel, N.C. Pattern of Self-Thinning in Jack Pine: Testing the Random Mortality Hypothesis. Ecology 1988, 69, 1017–1024. [Google Scholar] [CrossRef]
- Zhang, M.; Kang, X.; Meng, J.; Zhang, L. Distribution patterns and associations of dominant tree species in a mixed coniferous-broadleaf forest in the Changbai Mountains. J. Mt. Sci.-Engl. 2015, 12, 659–670. [Google Scholar] [CrossRef]
- Zhang, M.T. Spatial association and optimum adjacent distribution of trees in a mixed coniferous-broadleaf forest in northeastern China. Appl. Ecol. Environ. Res. 2017, 15, 1551–1564. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, Z.; Sun, I.F.; Song, B.; Ye, J.; Li, B.; Wang, X. Density dependence on tree survival in an old-growth temperate forest in northeastern China. Ann. For. Sci. 2009, 66, 204. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, J.; Kang, X. Spatial distribution pattern of dominant tree species in different disturbance plots in the Changbai Mountain. Sci. Rep. 2022, 12, 14161. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Liu, Q.; Liu, Z. Spatial Patterns and Interspecific Associations During Natural Regeneration in Three Types of Secondary Forest in the Central Part of the Greater Khingan Mountains, Heilongjiang Province, China. Forests 2020, 11, 152. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Jin, G. Species composition and community structure of a typical mixed broad-leaved-Korean pine (Pinus koraiensis) forest plot in Liangshui Nature Reserve, Northeast China. Biodivers. Sci. 2012, 20, 470–481. [Google Scholar] [CrossRef]
- Sasaki, T.; Konno, M.; Hasegawa, Y.; Imaji, A.; Terabaru, M.; Nakamura, R.; Ohira, N.; Matsukura, K.; Seiwa, K. Role of mycorrhizal associations in tree spatial distribution patterns based on size class in an old-growth forest. Oecologia 2019, 189, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Condit, R.S. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer: Berlin, Germany, 1998. [Google Scholar]
- Wiegand, T.; He, F.; Hubbell, S.P. A systematic comparison of summary characteristics for quantifying point patterns in ecology. Ecography 2013, 36, 92–103. [Google Scholar] [CrossRef]
- Illian, J.B.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; Wiley: Chichester, UK, 2008. [Google Scholar]
- Stoyan, D.; Stoyan, H. Fractals, Random Shapes, and Point Fields; Wiley: Chichester, UK, 1994. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Application with R; Chapman and Hall/CRC Press: London, UK, 2015. [Google Scholar]
- Law, R.; Illian, J.; Burslem, D.F.R.P.; Gratzer, G.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Ecological information from spatial patterns of plants: Insights from point process theory. J. Ecol. 2009, 97, 616–628. [Google Scholar] [CrossRef]
- Baddeley, A.; Diggle, P.J.; Hardegen, A.; Lawrence, T.; Milne, R.K.; Nair, G. On tests of spatial pattern based on simulation envelopes. Ecol. Monogr. 2014, 84, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Loosmore, N.B.; Ford, E.D. Statistical inference using the g or K point pattern spatial statistics. Ecology 2006, 87, 1925–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddeley, A.J.; Turner, R. spatstat: An R Package for Analyzing Spatial Point Patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CBS (25 ha) | ML (25 ha) | LS (9 ha) | FL (30 ha) | SS (10.4 ha) | |
|---|---|---|---|---|---|
| Total No. of KP trees (n) | 2286 | 4818 | 1175 | 3127 | 2347 |
| Stem density (n/ha) | 91.44 | 192.72 | 130.56 | 104.23 | 225.67 |
| Saplings (n/ha) | 0.12 (0.13%) | 103.24 (53.57%) | 10.89 (8.34%) | 19.10 (18.33%) | 95.77 (42.44%) |
| Juveniles (n/ha) | 4.44 (4.86%) | 57.20 (29.68%) | 5.67 (4.34%) | 5.23 (5.02%) | 43.37 (19.22%) |
| Mature trees (n/ha) | 85.16 (93.13%) | 32.24 (16.73%) | 100.89 (77.28%) | 68.07 (65.30%) | 86.44 (38.30%) |
| Large trees (n/ha) | 1.72 (1.88%) | 0.04 (0.02%) | 13.11 (10.04%) | 11.83 (11.35%) | 0.1 (0.04%) |
| DBH mean (cm) | 34.4 | 9.1 | 43.8 | 39.1 | 16.4 |
| DBH max (cm) | 98.4 | 84.1 | 109.0 | 111.6 | 71.3 |
| DBH min (cm) | 2.3 | 1.0 | 1.0 | 1.0 | 1.3 |
| Plot Name | CBS | ML | LS | FL | SS |
|---|---|---|---|---|---|
| Established year | 2004 | 2013 | 2005 | 2009 | 2012 |
| Inventory year | 2014 | 2018 | 2015 | 2019 | 2022 |
| Plot size (m2) | 500 × 500 | 400 × 260 | 300 × 300 | 500 × 600 | 500 × 500 |
| Latitude (N) | 42.38° | 43.95° | 47.19° | 48.08° | 49°5 |
| Longitude (E) | 128.08° | 130.07° | 128.87° | 129.12° | 126°75 |
| AE (m) | 801.5 | 765 | 408 | 419 | 450 |
| MAT (°C) | 3.6 | −2 | −0.3 | −0.5 | −2 |
| MAP (mm) | 700 | 530 | 676 | 688 | 520 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pak, U.; Guo, Q.; Liu, Z.; Wang, X.; Liu, Y.; Jin, G. Spatial Distribution of Pinus koraiensis Trees and Community-Level Spatial Associations in Broad-Leaved Korean Pine Mixed Forests in Northeastern China. Plants 2023, 12, 2906. https://doi.org/10.3390/plants12162906
Pak U, Guo Q, Liu Z, Wang X, Liu Y, Jin G. Spatial Distribution of Pinus koraiensis Trees and Community-Level Spatial Associations in Broad-Leaved Korean Pine Mixed Forests in Northeastern China. Plants. 2023; 12(16):2906. https://doi.org/10.3390/plants12162906
Chicago/Turabian StylePak, Unil, Qingxi Guo, Zhili Liu, Xugao Wang, Yankun Liu, and Guangze Jin. 2023. "Spatial Distribution of Pinus koraiensis Trees and Community-Level Spatial Associations in Broad-Leaved Korean Pine Mixed Forests in Northeastern China" Plants 12, no. 16: 2906. https://doi.org/10.3390/plants12162906