
Water and Nitrogen Coupling on the Regulation of Soil Nutrient–Microbial Biomass Balance and Its Effect on the Yield of Wolfberry (Lycium barbarum L.)
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Contents of Csoil, Nsoil, and Psoil and the Changes in the Ecological Stoichiometry
2.2. Characteristics of Cmic, Nmic, and Pmic, and the Changes in the Ecological Stoichiometry
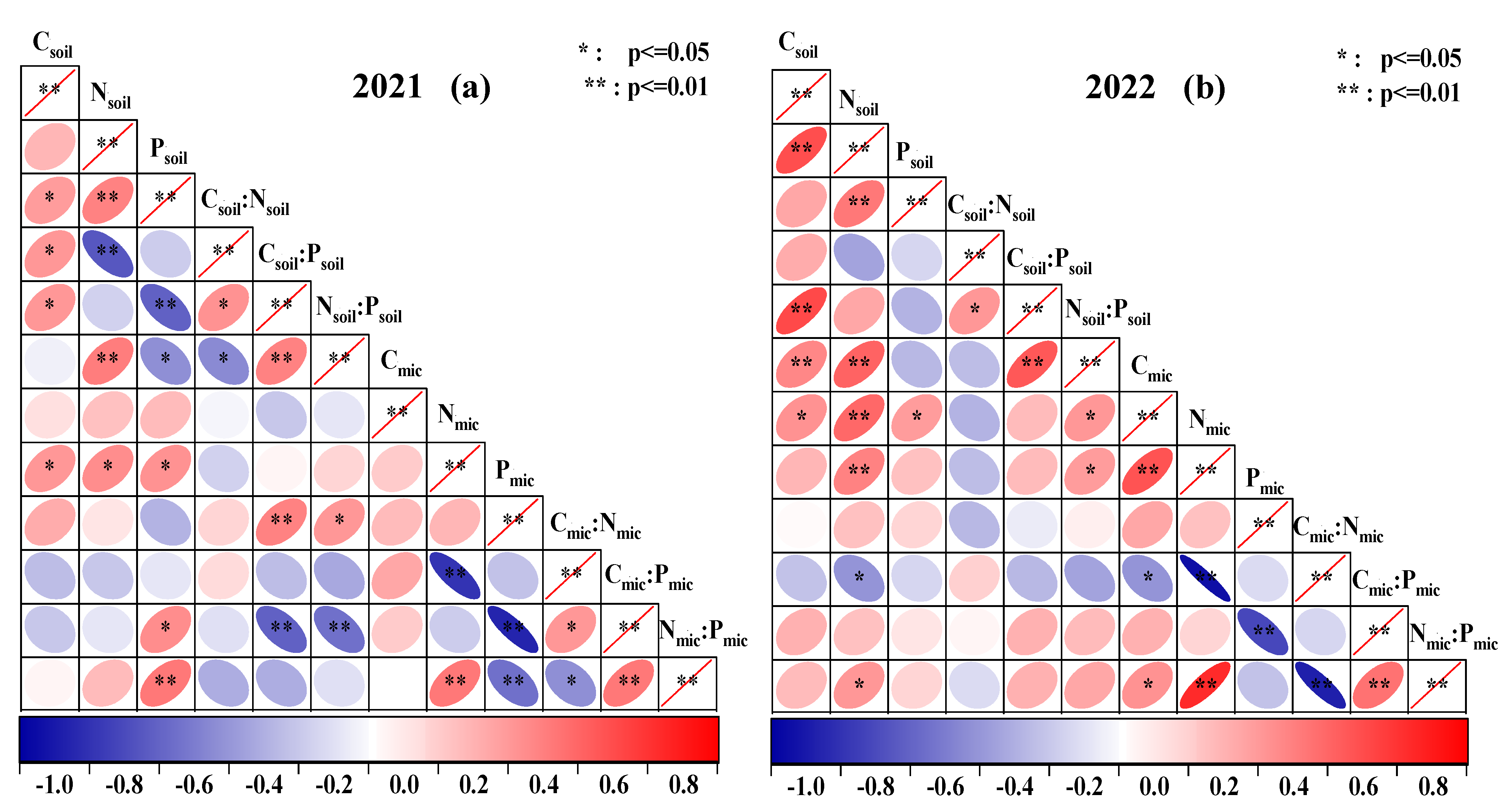
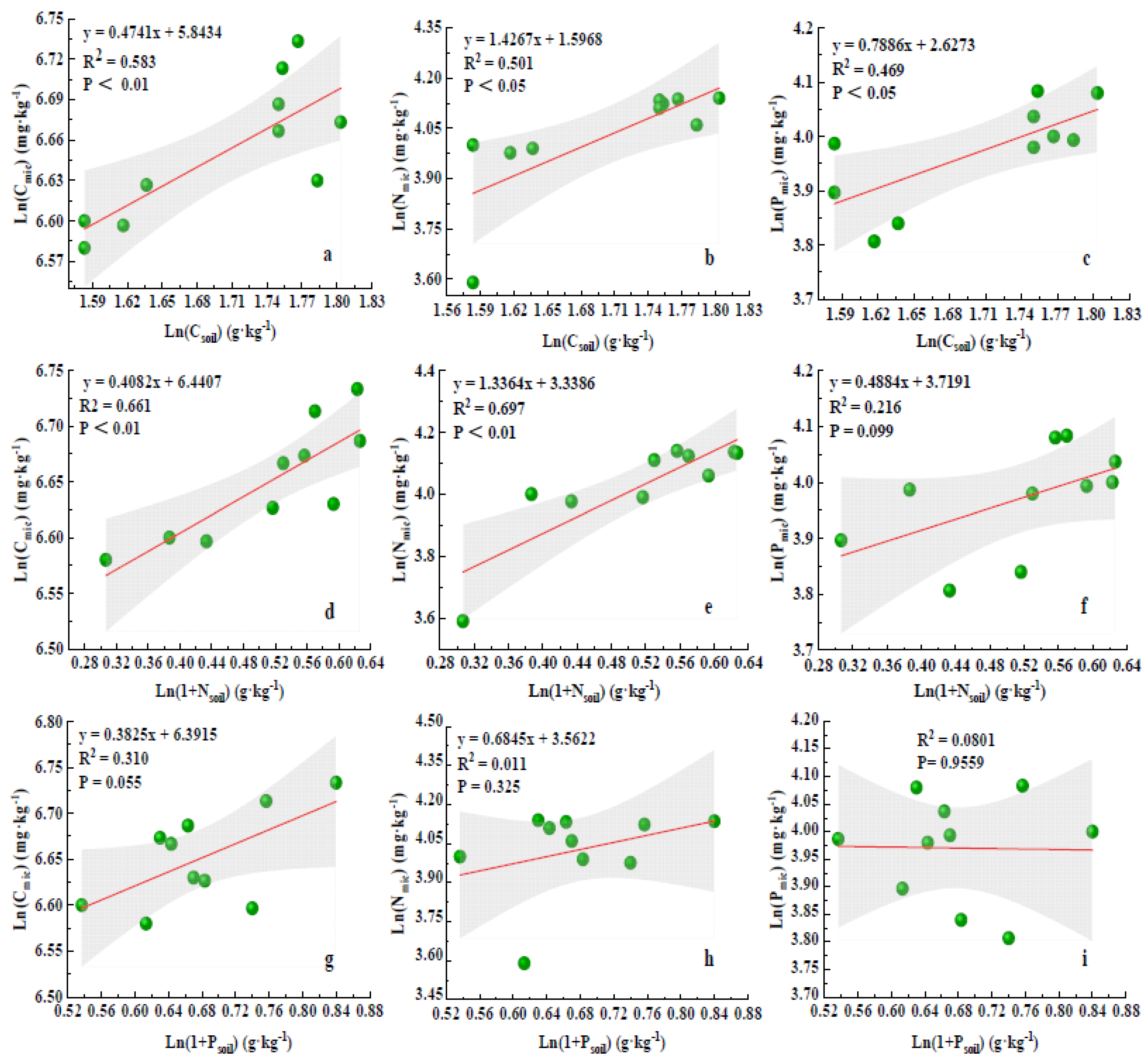
2.3. Correlation between Soil Nutrient–Microbial Biomass and the Ecological Stoichiometry Ratio
2.4. Effect of Water and Nitrogen Regulation on the Yield of Wolfberry
2.5. Integrated Evaluation of Soil Nutrient–Microbial Biomass and the Effect on the Stoichiometric Characteristics
2.5.1. Evaluation of Soil Nutrient–Microbial Biomass and the Stoichiometric Characteristics Based on PCA
2.5.2. Evaluation of Soil Nutrient–Microbial Biomass and the Stoichiometric Characteristics Based on the TOPSIS Method
2.6. Effect of Soil Quality on the Yield of Wolfberry
3. Discussion
3.1. The Response of Csoil, Nsoil, and Psoil Contents and Their Stoichiometric Imbalance to the Water and Nitrogen Coupling Regulation
3.2. The Response of Cmic, Nmic, Pmic, and Their Stoichiometric Imbalance to the Water and Nitrogen Coupling Regulation
3.3. The Response Mechanism of Wolfberry Yield to Soil Quality under Water and Nitrogen Coupling Regulation
4. Materials and Methods
4.1. Study Area
4.2. Experimental Design
4.3. Soil Sample Collection
4.4. Measurement Indices and Methods
4.4.1. Soil Index Measurements and Methods
4.4.2. Methods for Measuring the Yield of Wolfberry
4.5. Data Processing and Analysis
4.5.1. Principal Component Analysis
- (1)
- The formula for calculating each principal component score is as follows:
- (2)
- The data for the calculation of the combined score are as follows:
4.5.2. TOPSIS Method
- (1)
- A matrix was constructed based on the original evaluation parameters [49]. Assuming there were m evaluation objects and n evaluation indicators, the original data was expressed as the matrix = () m × n; where, represents the original data of the jth indicator of the ith treatment. The metrics were normalized as follows:Here, i = (1, 2, …, m) and j = (1, 2, …, n).
- (2)
- The normalization matrix can be expressed as = () m × n. The optimal and inferior vectors composed of the maximum and minimum values of each column were determined using Equations (4) and (5), respectively.
- (3)
- The Euclidean distances (Di+ and Di−) were determined using Equations (6) and (7).
- (4)
- The fit of the ith treatment to the optimal solution was determined using the equation.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Elser, J. Biological stoichiometry: A chemical brudge between ecology and evolutionary biology. Am. Nat. 2006, 168, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Gorokhova, E.; Fagan, W.F.; Markow, T.A. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.J.; Li, Q.K.; Fu, X.L.; Kou, L.; Dai, X.Q.; Wang, H.M. Foliar, root and rhizospheric soil C:N:P stoichiometries of overstory and understory species in subtropical plantations. Catena 2021, 198, 105020. [Google Scholar] [CrossRef]
- Bai, X.J.; Wang, B.R.; An, S.S.; Zeng, Q.C.; Zhang, H.X. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. Catena 2019, 183, 104186. [Google Scholar] [CrossRef]
- Carrillo, Y.; Bell, C.; Koyama, A.; Canarini, A.; Boot, C.M.; Wallenstein, M. Plant traits, stoichiometry and microbes as drivers of decomposition in the rhizosphere in a temperate grassland. J. Ecol. 2017, 105, 1750–1765. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Zhou, P.; Yang, L.H.; Gao, M. Effects of land reclamation on the physical, chemical, and microbial quantity and enzyme activity properties of degraded agricultural soils. J. Soils Sediments 2020, 20, 973–981. [Google Scholar] [CrossRef]
- Kwabiah, A.B.; Palm, C.A.; Stoskopf, N.C.; Voroney, R.P. Response of soil microbial biomass dynamics to quality of plant materials with emphasis on P availability. Soil Biol. Biochem. 2003, 35, 207–216. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, H.; Pumpanen, J.; Sietio, O.M.; Heinonsalo, J.; Koster, K. The impact of wildfire on microbial C:N:P stoichiometry and the fungal-to-bacterial ratio in permafrost soil. Biogeochemistry 2019, 142, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.C.; Liu, Y.; Fang, Y.; Ma, R.T.; Lal, R.T.; An, S.S. Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yun wu Mountain Reserve of China. Ecol. Eng. 2017, 109, 92–100. [Google Scholar] [CrossRef]
- Cui, H.; Ou, Y.; Lv, D.; Wang, L.X.; Liang, A.Z.; Yan, B.X. Aggregate-related microbial communities and nutrient stoichiometry under different croplands. Ecol. Process. 2020, 9, 1435–1443. [Google Scholar] [CrossRef]
- Xue, Q.; Newman, R.K.; Newman, C.W. Effects of heat treatment of barley starches on in vitro digestibility and glucose responses in rats. Cereal Chem. 1996, 73, 588–592. [Google Scholar]
- Yang, D.X.; Song, L.; Jin, G.Z. The soil C:N:P stoichiometry is more sensitive than the leaf C:N:P stoichiometry to nitrogen addition: A four-year nitrogen addition experiment in a Pinus koraiensis plantation. Plant Soil 2019, 442, 183–198. [Google Scholar] [CrossRef]
- Roldan, A.; Caravaca, F.; Hernandez, M.T.; Garcia, C.; Sanchez-Brito, C.; Velasquez, M. No-tillage, crop residue additions, and legume cover cropping effects on soil quality characteristics under maize in Patzcuaro watershed (Mexico). Soil Tillage Res. 2003, 72, 65–73. [Google Scholar] [CrossRef]
- Luo, Y.; Gong, Y.; Li, Y.M. Stoichiometry Characteristics of root, Stem and Leaf of Maize in Different Growth Stages in the Upper Reaches of the Tarim River. Res. Soil Water Conserv. 2018, 2502, 112–119. [Google Scholar]
- Duan, W.W.; Zhang, Z.J.; Zhu, J.J.; Zhang, D.; Qian, D.; Teng, F. Comparative Analysis of the Phenolic Profile of Lycium barbarum L. Fruits from Different Regions in China. Molecules 2022, 27, 5842. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Chang, S.C.; Inbaraj, B.S.; Chen, B.H. Isolation of carotenoids, flavonoids and polysaccharides from Lycium barbarum L. and evaluation of antioxidant activity. Food Chem. 2010, 120, 184–192. [Google Scholar] [CrossRef]
- Mao, F.; Xiao, B.; Jiang, Z.; Zhao, J.; Huang, X.; Guo, J. Anticancer effect of Lycium barbarum polysaccharides on coloncancer cells involves G0/G1 phase arrest. Med. Oncol. 2011, 28, 121–126. [Google Scholar] [CrossRef]
- Deng, Z.; Yin, J.; Wu, J.B.; Zhang, H.J. Comprehensive evaluation of water and fertilizer application for Lycium barbarum L. based on AHP and entropy weight method. J. Drain. Irrig. Mach. Eng. 2021, 39, 712–719. [Google Scholar]
- Huang, L.; Yang, W.P.; Hu, X.Q.; Tao, Y.; Yao, S.M.; Ou, X.Q. Effects of Irrigation and Nitrogen Interaction on Water Consumption Characteristics and Nitrogen Utilization of Winter Wheat. J. Soil Water Conserv. 2016, 30, 168–174. [Google Scholar]
- Ma, Z.H.; Yin, J.; Yang, Y.P.; Sun, F.B.; Yang, Z. Effect of water and nitrogen coupling regulation on the growth, physiology, yield, and quality attributes and comprehensive evaluation of wolfberry (Lycium barbarum L.). Front. Plant Sci. 2023, 14, 1130109. [Google Scholar] [CrossRef]
- Dai, Z.G.; Fei, L.J.; Huang, D.L.; Zeng, J.; Chen, L.; Cai, Y.H. Coupling effects of irrigation and nitrogen levels on yield, water and nitrogen use efficiency of surge-root irrigated jujube in a semiarid region. Agric. Water Manag. 2019, 213, 146–154. [Google Scholar] [CrossRef]
- Mahmoodi-Eshkaftaki, M.; Rafiee, M.R. Optimization of irrigation management: A multi-objective approach based on crop yield, growth, evapotranspiration, water use efficiency and soil salinity. J. Clean. Prod. 2020, 252, 119901. [Google Scholar] [CrossRef]
- Liu, S.X.; AN, H.; Zhang, X.W.; Xing, B.B.; Wen, Z.L.; Wang, B. Effects of Nitrogen and Phosphorus Addition on Soil Nutrient Content and Stoi-chiometry in Desert Grassland. Environ. Sci. 2023, 44, 2724–2734. [Google Scholar]
- Spieles, D.J.; Mitsch, W.J. Macroinvertebrate community structure in high-and low-nutrient constructed wetlands. Wetlands 2000, 20, 716–729. [Google Scholar] [CrossRef]
- Schleuss, P.M.; Widdig, M.; Heintz-Buschart, A.; Guhr, A.; Martin, S.; Kirkman, K. Stoichiometric controls of soil carbon and nitrogen cycling after long-term nitrogen and phosphorus addition in a mesic grassland in South Africa. Soil Biol. Biochem. 2019, 135, 294–303. [Google Scholar] [CrossRef]
- Zhan, S.X.; Wang, Y.; Zhu, Z.C.; Li, W.H.; Bai, Y.F. Nitrogen enrichment alters plant N:P stoichiometry and intensifies phosphorus limitation in a steppe ecosystem. Environ. Exp. Bot. 2017, 134, 21–32. [Google Scholar] [CrossRef]
- Wang, F.; Li, Q.H.; Lin, C.; Zhong, S.J.; He, C.M.; Liu, Y.J. Effect of different fertilization modes on topsoil organic carbon sequestration and productivity in yellow paddy field of southern China. Plant Nutr. Fert. Sci. 2015, 21, 1447–1454. [Google Scholar]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; Huang, J.H. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Gao, Y.; He, N.P.; Yu, G.R.; Chen, W.L.; Wang, Q.F. Long-term effects of different land use types on C, N and P stoichiometry and storage in subtropical ecosystems: A case study in China. Ecol. Eng. 2014, 67, 171–181. [Google Scholar] [CrossRef]
- Jiang, Y.F.; Guo, X. Stoichiometric patterns of soil carbon, nitrogen, and phosphorus in farmland of the Poyang Lake region in Southern China. J. Soils Sediments 2019, 19, 3476–3488. [Google Scholar] [CrossRef]
- Truu, M.; Truu, J.; Heinsoo, K. Changes in soil microbial community under willow coppice: The effect of irrigation with secondary-treated municipal wastewater. Ecol. Eng. 2009, 35, 1011–1020. [Google Scholar] [CrossRef]
- Ren, C.J.; Zhao, F.Z.; Shi, Z.; Chen, J.; Han, X.H.; Yang, G.H. Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation. Soil Biol. Biochem. 2017, 115, 1–10. [Google Scholar] [CrossRef]
- Denef, K.; Six, J.; Paustian, K.; Merckx, R. Importance of macroaggregate dynamics in controlling soil carbon stabilization: Short-term effects of physical disturbance induced by dry-wet cycles. Soil Biol. Biochem. 2001, 33, 2145–2153. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef]
- Wang, L.J.; Chen, R.M.; Xiao, W.F.; Sun, P.F.; Shen, Y.F.; Zeng, L.X. Effects of nitrogen addition on soil microbial biomass and enzyme activities of Pinus massoniana-Quercus variabilis mixed plantations in the Three Gorges Reservoir Area. J. Appl. Sci. 2020, 33, 42–50. [Google Scholar]
- Zhu, X.M.; Liu, M.; Yin, H.J. Differential effects of N addition on the stoichiometry of microbes and extracellular enzymes in the rhizosphere and bulk soils of an alpine shrubland. Plant Soil 2020, 449, 285–301. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Zhou, J.C.; Liu, X.F.; Zheng, Y.; Ji, Y.H.; Li, X.F.; Xu, P.C. Effects of nitrogen deposition on soil microbial biomass and enzyme activities in Castanopsis carlesii natural forests in subtropical regions. Acta Ecol. Sin. 2017, 37, 127–135. [Google Scholar]
- Xu, X.F.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Sandhu, O.S.; Gupta, R.K.; Thind, H.S.; Jat, M.L.; Sidhu, H.S.; Yadvinder-Singh. Drip irrigation and nitrogen management for improving crop yields, nitrogen use efficiency and water productivity of maize-wheat system on permanent beds in north-west India. Agric. Water Manag. 2019, 219, 19–26. [Google Scholar] [CrossRef]
- Qian, Y.; Follett, R.F.; Kimble, J.M. Soil Organic Carbon Input from Urban Turfgrasses. Soil Sci. Soc. Am. J. 2010, 74, 366–371. [Google Scholar] [CrossRef]
- Gehl, R.J.; Schmidt, J.P.; Maddux, L.D.; Gordon, W.B. Corn yield response to nitrogen rate and timing in sandy irrigated soils. Agron. J. 2005, 97, 1230–1238. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.Y.; Yang, M.; Liu, M.; Lu, Y.; Yang, H. Nitrogen and phosphorus fertilizations alter nitrogen, phosphorus and potassium resorption of alfalfa in the Loess Plateau of China. J. Plant Nutr. 2019, 42, 2234–2246. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, H.W.; Yi, H.J. Impact of fertilizer on crop yield and C:N:P stoichiometry in arid and semi-arid soil. Int. J. Environ. Res. Public Health 2021, 18, 4341. [Google Scholar] [CrossRef] [PubMed]
- Ningxia Water Resources Department and Ningxia Institute of Water Resources Science. Ningxia Drip Irrigation Engineering Planning and Design Guidelines and Technical Regulations for Drip Irrigation Planting of Advantageous Specialty Crops; Ningxia People’s Publishing House: Yinchuan, China, 2017; Available online: https://item.kongfz.com/book/41091018.html (accessed on 26 June 2023).
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B. Linking microbial C:N:P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar] [CrossRef]
- Pan, L.D.; Li, R.; Zhang, Y.S.; Li, Q.G.; Gao, J.Y. Effects of straw mulching on soil ecological stoichiometry characteristics and yield on sloping farmland in karst area, Southwestern China. Acta Ecol. Sin. 2022, 42, 4428–4438. [Google Scholar]
- Liu, Y.W.; Zhou, X.; Han, H.H.; Yang, Q.L.; Liu, X.G. Coupling scheme optimization of Panax notoginseng considering yield, quality and water-fertilizer use efficiency. Trans. Chin. Soc. Agric. Eng. 2021, 37, 139–146. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Analysis Items | Principal Component 1 | Principal Component 2 | Principal Component 3 | Analysis Items | Principal Component 1 | Principal Component 2 | Principal Component 3 |
|---|---|---|---|---|---|---|---|---|
| 2021 | Csoil | 0.776 | 0.190 | 0.348 | Pmic | 0.861 | −0.339 | 0.273 |
| Nsoil | 0.740 | 0.502 | 0.202 | Cmic:Nmic | −0.701 | −0.349 | 0.559 | |
| Psoil | 0.009 | 0.854 | 0.487 | Cmic:Pmic | −0.746 | 0.626 | −0.096 | |
| Csoil:Nsoil | −0.371 | −0.619 | 0.282 | Nmic:Pmic | −0.018 | 0.776 | −0.586 | |
| Csoil:Psoil | 0.669 | −0.691 | −0.157 | Characteristics rate | 5.11 | 3.66 | 1.71 | |
| Nsoil:Psoil | 0.770 | −0.442 | −0.322 | Contribution rate/% | 42.6 | 30.5 | 14.2 | |
| Cmic | 0.544 | 0.429 | 0.555 | Cumulative contribution rate/% | 42.6 | 73.1 | 87.3 | |
| Nmic | 0.834 | 0.409 | −0.233 | |||||
| 2022 | Csoil | 0.891 | 0.307 | 0.125 | Pmic | 0.493 | 0.369 | 0.772 |
| Nsoil | 0.972 | −0.093 | 0.096 | Cmic:Nmic | −0.931 | 0.139 | 0.092 | |
| Psoil | 0.385 | −0.715 | 0.525 | Cmic:Pmic | 0.408 | −0.469 | −0.682 | |
| Csoil:Nsoil | −0.618 | 0.611 | 0.059 | Nmic:Pmic | 0.922 | −0.178 | −0.269 | |
| Csoil:Psoil | 0.492 | 0.818 | −0.286 | Characteristics rate | 7.04 | 2.24 | 1.66 | |
| Nsoil:Psoil | 0.806 | 0.421 | −0.301 | Contribution rate/% | 58.6 | 18.7 | 13.8 | |
| Cmic | 0.885 | 0.022 | 0.192 | Cumulative contribution rate/% | 58.6 | 77.3 | 91.1 | |
| Nmic | 0.982 | 0.011 | 0.075 |
| Year | Treatment | Principal Component 1 | Principal Component 2 | Principal Component 3 | Comprehensive Scores | Ranking | Year | Treatment | Principal Component 1 | Principal Component 2 | Principal Component 3 | Comprehensive Scores | Ranking | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | I1 | N1 | −4.17 | −2.47 | 1.88 | −2.26 | 10 | 2021 | I1 | N1 | −6.14 | 1.35 | 0.83 | −3.23 | 10 |
| N2 | −1.50 | 1.73 | −1.54 | −0.33 | 7 | N2 | −0.64 | −1.68 | −0.75 | −0.79 | 7 | ||||
| N3 | 1.76 | 0.15 | −0.40 | 0.74 | 4 | N3 | 0.53 | 1.31 | −0.26 | 0.52 | 5 | ||||
| I2 | N1 | −3.20 | 1.72 | −1.23 | −1.01 | 9 | I2 | N1 | −1.48 | −2.68 | −0.67 | −1.46 | 9 | ||
| N2 | 1.45 | 0.20 | 1.11 | 0.84 | 3 | N2 | 1.14 | −0.99 | 3.00 | 0.90 | 4 | ||||
| N3 | 0.40 | 3.46 | 1.98 | 1.51 | 1 | N3 | 2.19 | −0.68 | 0.61 | 1.24 | 3 | ||||
| I3 | N1 | −0.31 | −2.15 | −1.39 | −0.98 | 8 | I3 | N1 | −1.68 | 0.71 | −0.85 | −0.97 | 8 | ||
| N2 | 1.94 | −1.96 | 0.48 | 0.30 | 6 | N2 | 2.51 | 2.13 | 0.35 | 1.92 | 1 | ||||
| N3 | 2.25 | −0.09 | −0.27 | 0.89 | 2 | N3 | 2.54 | 0.27 | −0.67 | 1.45 | 2 | ||||
| CK | 1.36 | −0.59 | −0.63 | 0.31 | 5 | CK | 1.02 | 0.26 | −1.57 | 0.43 | 6 | ||||
| Year | Treatment | Soil Nutrient–Microbial Biomass and Stoichiometric Characteristics Index | Di± | Di− | Ci | Ranking | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Csoil | Nsoil | Psoil | Csoil:Nsoil | Csoil:Psoil | Nsoil:Psoil | Cmic | Nmic | Pmic | Cmic:Nmic | Cmic:Pmic | Nmic:Pmic | ||||||
| 2021 | I1N1 | 0.283 | 0.265 | 0.299 | 0.354 | 0.294 | 0.279 | 0.297 | 0.210 | 0.276 | 0.451 | 0.333 | 0.233 | 0.294 | 0.209 | 0.415 | 10 |
| I1N2 | 0.299 | 0.320 | 0.318 | 0.288 | 0.291 | 0.313 | 0.302 | 0.299 | 0.254 | 0.307 | 0.371 | 0.367 | 0.246 | 0.410 | 0.625 | 5 | |
| I1N3 | 0.355 | 0.352 | 0.322 | 0.311 | 0.339 | 0.338 | 0.305 | 0.335 | 0.323 | 0.279 | 0.295 | 0.326 | 0.228 | 0.406 | 0.640 | 3 | |
| I2N1 | 0.277 | 0.272 | 0.330 | 0.332 | 0.259 | 0.256 | 0.300 | 0.308 | 0.250 | 0.297 | 0.368 | 0.381 | 0.285 | 0.417 | 0.594 | 8 | |
| I2N2 | 0.332 | 0.327 | 0.321 | 0.316 | 0.321 | 0.315 | 0.343 | 0.337 | 0.353 | 0.309 | 0.301 | 0.300 | 0.213 | 0.380 | 0.641 | 2 | |
| I2N3 | 0.335 | 0.355 | 0.405 | 0.291 | 0.256 | 0.273 | 0.350 | 0.338 | 0.319 | 0.314 | 0.338 | 0.329 | 0.221 | 0.422 | 0.656 | 1 | |
| I3N1 | 0.259 | 0.260 | 0.248 | 0.309 | 0.331 | 0.334 | 0.309 | 0.310 | 0.332 | 0.303 | 0.285 | 0.291 | 0.296 | 0.339 | 0.534 | 9 | |
| I3N2 | 0.349 | 0.315 | 0.294 | 0.348 | 0.372 | 0.336 | 0.317 | 0.330 | 0.360 | 0.293 | 0.271 | 0.285 | 0.244 | 0.381 | 0.609 | 7 | |
| I3N3 | 0.328 | 0.353 | 0.306 | 0.289 | 0.333 | 0.358 | 0.321 | 0.337 | 0.346 | 0.289 | 0.287 | 0.304 | 0.238 | 0.389 | 0.621 | 6 | |
| CK | 0.331 | 0.323 | 0.298 | 0.319 | 0.345 | 0.343 | 0.313 | 0.335 | 0.326 | 0.285 | 0.295 | 0.320 | 0.234 | 0.391 | 0.626 | 4 | |
| Z± | 0.355 | 0.355 | 0.405 | 0.354 | 0.372 | 0.358 | 0.350 | 0.338 | 0.360 | 0.451 | 0.371 | 0.381 | |||||
| Z− | 0.259 | 0.260 | 0.248 | 0.288 | 0.256 | 0.256 | 0.297 | 0.210 | 0.250 | 0.279 | 0.271 | 0.233 | |||||
| 2022 | I1N1 | 0.276 | 0.249 | 0.291 | 0.354 | 0.299 | 0.269 | 0.290 | 0.200 | 0.312 | 0.464 | 0.293 | 0.201 | 0.290 | 0.210 | 0.420 | 10 |
| I1N2 | 0.287 | 0.310 | 0.313 | 0.291 | 0.289 | 0.313 | 0.312 | 0.297 | 0.303 | 0.313 | 0.325 | 0.312 | 0.220 | 0.179 | 0.448 | 7 | |
| I1N3 | 0.332 | 0.327 | 0.306 | 0.317 | 0.341 | 0.338 | 0.313 | 0.303 | 0.317 | 0.308 | 0.311 | 0.302 | 0.194 | 0.208 | 0.517 | 6 | |
| I2N1 | 0.300 | 0.306 | 0.340 | 0.308 | 0.277 | 0.284 | 0.298 | 0.279 | 0.287 | 0.321 | 0.331 | 0.313 | 0.227 | 0.173 | 0.433 | 8 | |
| I2N2 | 0.326 | 0.332 | 0.354 | 0.308 | 0.289 | 0.296 | 0.329 | 0.346 | 0.348 | 0.284 | 0.297 | 0.314 | 0.215 | 0.232 | 0.519 | 5 | |
| I2N3 | 0.332 | 0.341 | 0.335 | 0.306 | 0.312 | 0.321 | 0.336 | 0.356 | 0.327 | 0.283 | 0.324 | 0.343 | 0.198 | 0.259 | 0.566 | 3 | |
| I3N1 | 0.299 | 0.291 | 0.294 | 0.322 | 0.321 | 0.314 | 0.291 | 0.290 | 0.302 | 0.305 | 0.303 | 0.308 | 0.226 | 0.168 | 0.427 | 9 | |
| I3N2 | 0.345 | 0.337 | 0.305 | 0.321 | 0.356 | 0.348 | 0.328 | 0.367 | 0.338 | 0.266 | 0.307 | 0.343 | 0.210 | 0.280 | 0.572 | 2 | |
| I3N3 | 0.330 | 0.347 | 0.311 | 0.300 | 0.334 | 0.351 | 0.333 | 0.353 | 0.321 | 0.282 | 0.327 | 0.347 | 0.199 | 0.267 | 0.573 | 1 | |
| CK | 0.327 | 0.311 | 0.308 | 0.330 | 0.334 | 0.319 | 0.328 | 0.336 | 0.303 | 0.292 | 0.341 | 0.351 | 0.196 | 0.244 | 0.554 | 4 | |
| Z± | 0.345 | 0.347 | 0.354 | 0.354 | 0.356 | 0.351 | 0.336 | 0.367 | 0.348 | 0.464 | 0.341 | 0.351 | |||||
| Z− | 0.276 | 0.249 | 0.291 | 0.291 | 0.277 | 0.269 | 0.290 | 0.200 | 0.287 | 0.266 | 0.293 | 0.201 | |||||
| Year | Depth | pH | EC | Total N | Total P | Total K | Organic Material | Ammonium N | Nitrate N |
|---|---|---|---|---|---|---|---|---|---|
| cm | μs cm−1 | g kg−1 | mg kg−1 | ||||||
| 2021 | 0–20 | 8.00 | 1615 | 0.46 | 0.43 | 16.19 | 7.58 | 12.67 | 14.69 |
| 20–40 | 8.20 | 1091 | 0.42 | 0.42 | 17.15 | 6.73 | 4.51 | 4.73 | |
| 2022 | 0–20 | 8.06 | 1524 | 0.45 | 0.40 | 17.00 | 7.21 | 12.24 | 14.17 |
| 20–40 | 8.12 | 963 | 0.41 | 0.41 | 17.00 | 6.28 | 4.42 | 4.82 | |
| Fertility Stages | 2021 | 2022 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Date | Irrigation Amount m2 hm−2 | Nitrogen Amount kg hm−2 | Date | Irrigation Amount m2 hm−2 | Nitrogen Amount kg hm−2 | |||||||||
| I1 | I2 | I3 | N1 | N2 | N3 | I1 | I2 | I3 | N1 | N2 | N3 | |||
| Spring tip period | 04–30 | 432 | 513.0 | 594 | 24.75 | 33.75 | 42.75 | 04–28 | 432 | 513.0 | 594 | 24.75 | 33.75 | 42.75 |
| Flowering period | 05–23 | 216 | 256.5 | 297 | 12.38 | 16.88 | 21.38 | 05–23 | 216 | 256.5 | 297 | 12.38 | 16.88 | 21.38 |
| 06–18 | 432 | 513.0 | 594 | 28.88 | 39.38 | 49.88 | 06–15 | 432 | 513.0 | 594 | 28.88 | 39.38 | 49.88 | |
| Fruit ripening period | 07–05 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | 07–02 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 |
| 07–15 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | 07–15 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | |
| 07–25 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | 07–28 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | |
| 08–05 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | 08–08 | 216 | 256.5 | 297 | 20.63 | 28.13 | 35.63 | |
| Defoliation period | 08–25 | 216 | 256.5 | 297 | 16.50 | 22.50 | 28.50 | 08–28 | 216 | 256.5 | 297 | 16.50 | 22.50 | 28.50 |
| Total | 2160 | 2565 | 2970 | 165 | 225 | 285 | 2160 | 2565 | 2970 | 165 | 225 | 285 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Ma, Z.; Yang, Y.; Du, B.; Sun, F.; Yang, Z. Water and Nitrogen Coupling on the Regulation of Soil Nutrient–Microbial Biomass Balance and Its Effect on the Yield of Wolfberry (Lycium barbarum L.). Plants 2023, 12, 2768. https://doi.org/10.3390/plants12152768
Yin J, Ma Z, Yang Y, Du B, Sun F, Yang Z. Water and Nitrogen Coupling on the Regulation of Soil Nutrient–Microbial Biomass Balance and Its Effect on the Yield of Wolfberry (Lycium barbarum L.). Plants. 2023; 12(15):2768. https://doi.org/10.3390/plants12152768
Chicago/Turabian StyleYin, Juan, Zhenghu Ma, Yingpan Yang, Bin Du, Fubin Sun, and Zhen Yang. 2023. "Water and Nitrogen Coupling on the Regulation of Soil Nutrient–Microbial Biomass Balance and Its Effect on the Yield of Wolfberry (Lycium barbarum L.)" Plants 12, no. 15: 2768. https://doi.org/10.3390/plants12152768