
Molecular Mechanisms Underlying Potential Pathogen Resistance in Cannabis sativa


Abstract
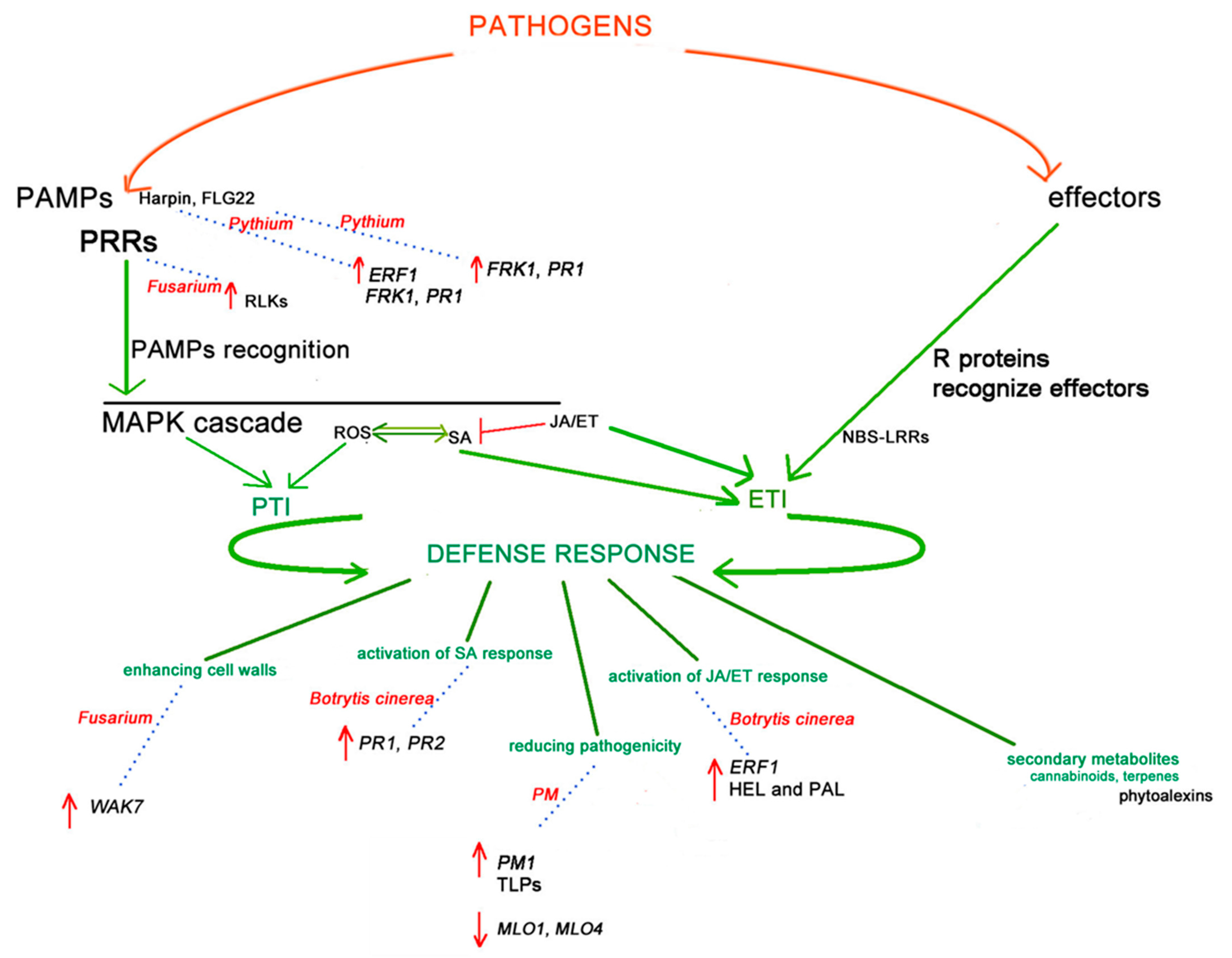
:1. Introduction
2. Overview of Cannabis Resistance Genes to Pathogens
3. Powdery Mildew Pathogens—Golovinomyces spp.
4. Fusarium spp.
5. Botrytis cinerea
6. Pythium
7. Genome Editing to Generate Disease-Resistant Cannabis Varieties
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clarke, R.C.; Merlin, M.D. Cannabis domestication, breeding history, present day genetic diversity, and future prospects. Crit. Rev. Plant Sci. 2016, 35, 293–327. [Google Scholar] [CrossRef]
- Stuyt, E. The problem with the current high potency THC marijuana from the perspective of an addiction psychiatrist. Mo. Med. 2018, 115, 482–486. [Google Scholar] [PubMed]
- Schwabe, A.L.; Johnson, V.; Harrelson, J.; McGlaughlin, M.E. Uncomfortably high: Testing reveals inflated THC potency on retail Cannabis labels. PLoS ONE 2023, 18, e0282396. [Google Scholar] [CrossRef]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular targets of the phytocannabinoids: A complex picture. Prog. Chem. Org. Nat. Prod. 2017, 103, 103–131. [Google Scholar] [PubMed] [Green Version]
- Sirangelo, T.M.; Ludlow, R.A.; Spadafora, N.D. Multi-Omics approaches to study molecular mechanisms in Cannabis sativa. Plants 2022, 11, 2182. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; Elsohly, M.A. Cannabis sativa L.: Botany and horticulture. In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., Elsohly, M.A., Eds.; Springer: Cham, Switzerland, 2017; pp. 79–100. [Google Scholar]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.A.; Lewsey, M.G. Recent advances in Cannabis sativa genomics research. N. Phytol. 2020, 230, 73–89. [Google Scholar] [CrossRef]
- Pacula, R.L.; Smart, R. Medical marijuana and marijuana legalization. Ann. Rev. Clin. Psychol. 2017, 13, 397–419. [Google Scholar] [CrossRef]
- Gao, S.; Wang, B.; Xie, S.; Xu, X.; Zhang, J.; Pei, L.; Yu, Y.; Yang, W.; Zhang, Y. A high quality reference genome of wild Cannabis sativa. Hortic. Res. 2020, 7, 73. [Google Scholar] [CrossRef]
- Petit, J.; Salentijn, E.M.J.; Paulo, M.J.; Denneboom, C.; van Loo, E.N.; Trindade, L.M. Elucidating the genetic architecture of fiber quality in hemp (Cannabis sativa L.) using a Genome-Wide Association study. Front. Genet. 2020, 11, 566314. [Google Scholar] [CrossRef]
- Sirangelo, T.M.; Ludlow, R.A.; Chenet, T.; Pasti, L.; Spadafora, N.D. Multi-Omics and genome editing studies on plant cell walls to improve biomass quality. Agriculture 2023, 13, 752. [Google Scholar] [CrossRef]
- Punja, Z.K. Emerging diseases of Cannabis sativa and sustainable management. Pest Manag. Sci. 2021, 77, 3857–3870. [Google Scholar] [CrossRef]
- Punja, Z.K.; Collyer, D.; Scott, C.; Lung, S.; Holmes, J.; Sutton, D. Pathogens and molds affecting production and quality of Cannabis sativa L. Front. Plant Sci. 2019, 10, 1120. [Google Scholar] [CrossRef] [Green Version]
- McCain, A.H.; Noviello, C. Biological control of Cannabis sativa. In Proceedings of the VI International Symposium on Biological Control of Weeds, Vancouver, WA, Canada, 19–25 August 1984; Delfosse, E.S., Ed.; Agricultural Canada: Ottawa, ON, Canada, 1985; pp. 635–642. [Google Scholar]
- Punja, Z.K. Epidemiology of Fusarium oxysporum causing root and crown rot of Cannabis (Cannabis sativa L., marijuana) plants in commercial greenhouse production. Can. J. Plant Pathol. 2020, 43, 216–235. [Google Scholar] [CrossRef]
- Cheng, Y.; Tang, X.; Gao, C.; Li, Z.; Chen, J.; Guo, L.; Wang, T.; Xu, J. Molecular diagnostics and pathogenesis of fungal pathogens on bast fiber crops. Pathogens 2020, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Punja, Z.K.; Rodriguez, G. Fusarium and Pythium species infecting roots of hydroponically grown marijuana (Cannabis sativa L.) plants. Can. J. Plant Pathol. 2018, 40, 498–513. [Google Scholar] [CrossRef] [Green Version]
- Warren, J.G.; Mercado, J.; Grace, D. Occurrence of hop latent viroid causing disease in Cannabis sativa in California. Plant Dis. 2019, 103, 2699. [Google Scholar] [CrossRef]
- Wolfenbarger, S.N.; Massie, S.T.; Ocamb, C.; Eck, E.B.; Grove, G.G.; Nelson, M.E.; Probst, C.; Twomey, M.C.; Gent, D.H. Distribution and characterization of Podosphaera macularis virulent on hop cultivars possessing R6-Based resistance to Powdery Mildew. Plant Dis. 2016, 100, 1212–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; APS Press: St. Paul, MN, USA, 2006. [Google Scholar]
- Vujanovic, V.; Korber, D.R.; Vujanovic, S.; Vujanovic, J.; Jabaji, S. Scientific prospects for cannabis-microbiome research to ensure quality and safety of products. Microorganisms 2020, 8, 290. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and perspectives in the use of biocontrol agents against fungal plant diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Massart, S.; Martinez-Medina, M.; Jijakli, M.H. Biological control in the microbiome era: Challenges and opportunities. Biol. Control 2015, 89, 98–108. [Google Scholar] [CrossRef]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [Green Version]
- Pépin, N.; Punja, Z.K.; Joly, D.L. Occurrence of powdery mildew caused by Golovinomyces chicoracearum sensu lato on Cannabis sativa in Canada. Plant Dis. 2018, 102, 2644. [Google Scholar] [CrossRef]
- Garfinkel, A.R. Three Botrytis species found causing gray mold on industrial hemp (Cannabis sativa) in Oregon. Plant Dis. 2020, 104, 2026. [Google Scholar] [CrossRef] [Green Version]
- Aliferis, K.A.; Bernard-Perron, D. Cannabinomics: Application of metabolomics in Cannabis (Cannabis sativa L.). Research and development. Front. Plant. Sci. 2020, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Rodriguez, G.; Chen, S. Assessing genetic diversity in Cannabis sativa using molecular approaches. In Cannabis sativa L.-Botany and Biotechnology; Chandra, S., Lata, L., ElSohly, M.A., Eds.; Springer: Berlin, Germany, 2017; pp. 395–418. [Google Scholar]
- Scott, C.; Punja, Z.K. Evaluation of disease management approaches for powdery mildew on Cannabis sativa L. (marijuana) plants. Can. J. Plant Pathol. 2021, 34, 394–412. [Google Scholar] [CrossRef]
- Stack, G.M.; Toth, J.A.; Carlson, C.H.; Cala, A.R.; Marrero-González, M.I.; Wilk, R.L.; Gentner, D.R.; Crawford, J.L.; Philippe, G.; Rose, J.K.C. Season-long characterization of high-cannabinoid hemp (Cannabis sativa L.) reveals variation in cannabinoid accumulation, flowering time, and disease resistance. GCB Bioenergy 2021, 13, 546–561. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, C.C.; Hulst, R.V.D.; Meijer-Dekens, F.; Bonnema, G.; Lindhout, P. QTLs for tomato powdery mildew resistance (Oidium lycopersici) in Lycopersicon parviflorum G1.1601 localize with two qualitative powdery mildew resistance genes. Mol. Plant Microbe Interact. 2003, 16, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Tassone, M.R.; Bagnaresi, P.; Desiderio, F.; Bassolino, L.; Barchi, L.; Florio, F.E.; Sunseri, F.; Sirangelo, T.M.; Rotino, G.L.; Toppino, L. A Genomic BSAseq Approach for the characterization of QTLs underlying resistance to Fusarium oxysporum in eggplant. Cells 2022, 11, 2548. [Google Scholar] [CrossRef]
- Nachappa, P.; Fulladolsa, A.C.; Stenglein, M. Wild wild west: Emerging viruses and viroids of hemp. Outlooks Pest Manag. 2020, 31, 175–179. [Google Scholar] [CrossRef]
- Oh, S.; Choi, D. Receptor-mediated nonhost resistance in plants. Essays Biochem. 2022, 66, 435–445. [Google Scholar]
- Lee, H.A.; Lee, H.Y.; Seo, E.; Lee, J.; Kim, S.B.; Oh, S.; Choi, E.; Choi, E.; Lee, S.E.; Choi, D. Current understandings of plant nonhost resistance. Mol. Plant-Microbe Interact. 2017, 30, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Sands, L.B.; Cheek, T.; Reynolds, J.; Ma, Y.; Berkowitz, G.A. Effects of Harpin and Flg22 on growth enhancement and pathogen defense in Cannabis sativa seedlings. Plants 2022, 11, 1178. [Google Scholar] [CrossRef]
- Benjamin, G.; Pandharikar, G.; Frendo, P. Salicylic acid in plant symbioses: Beyond plant pathogen interactions. Biology 2022, 11, 861. [Google Scholar] [CrossRef]
- Ding, L.N.; Li, Y.T.; Wu, Y.Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.-L. Plant disease resistance-related signaling pathways: Recent progress and future prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef] [PubMed]
- Boba, A.; Kostyn, K.; Kozak, B.; Zalewski, I.; Szopa, J.; Kulma, A. Transcriptomic profiling of susceptible and resistant flax seedlings after Fusarium oxysporum lini infection. PLoS ONE 2021, 16, e0246052. [Google Scholar] [CrossRef] [PubMed]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R.W. Plant NBS-LRR proteins: Adaptable guards. Genome Biol. 2006, 7, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfo, G.; Dohm, J.C.; Himmelbauer, H. Prediction of NB-LRR resistance genes based on full-length sequence homology. Plant J. 2022, 110, 1592–1602. [Google Scholar] [CrossRef]
- Xie, S.S.; Duan, C.G. Epigenetic regulation of plant immunity: From chromatin codes to plant disease resistance. aBIOTECH 2023, 1–16. [Google Scholar] [CrossRef]
- He, Z.H.; He, D.; Kohorn, B.D. Requirement for the induced expression of a cell wall associated receptor kinase for survival during the pathogen response. Plant J. 1998, 14, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Diener, A.C.; Ausubel, F.M. Resistance to Fusarium oxysporum 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 2005, 171, 305–321. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Xie, M.; Wang, X.; Wang, G.; Zhang, Y.; Li, Z.; Ma, Z. Identification of cell wall-associated kinases as important regulators involved in Gossypium hirsutum resistance to Verticillium dahliae. BMC Plant Biol. 2021, 21, 220. [Google Scholar] [CrossRef]
- Li, L.; Yu, S.; Chen, J.; Cheng, C.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Releasing the full potential of Cannabis through biotechnology. Agronomy 2022, 12, 2439. [Google Scholar] [CrossRef]
- Sipahi, H.; Whyte, T.D.; Ma, G.; Berkowitz, G. Genome-Wide identification and expression analysis of Wall-Associated Kinase (WAK) gene family in Cannabis sativa L. Plants 2022, 11, 2703. [Google Scholar] [CrossRef] [PubMed]
- Maravaneh, H.; Davarpanah, S.J. Study of cannabinoids biosynthesis-related genes in hemp (Cannabis sativa L.) under drought stress by in vitro and in silico tools. J. Appl. Biotechnol. Rep. 2022, 9, 504–510. [Google Scholar]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. Int. J. Mol. Sci. 2021, 22, 5710. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D.S. Plant natural products targeting bacterial virulence factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Woods, W.; Price, N.; Matthews, P.; McKay, J.K. Genome-wide polymorphism and genic selection in feral and domesticated lineages of Cannabis sativa. G3 2023, 13, jkac209. [Google Scholar] [CrossRef] [PubMed]
- Sommano, S.R.; Chittasupho, C.; Ruksiriwanich, W.; Jantrawut, P. The Cannabis terpenes. Molecules 2020, 25, 5792. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Schwinghamer, T.; Rosenbaum, P.; McCarty, V.; Eichhorn Bilodeau, S.; Lyu, D.; Ahmed, M.B.; Robinson, G.; Lefsrud, M.; Wilkins, O.; et al. Closing the yield gap for Cannabis: A meta-analysis of factors determining Cannabis yield. Front. Plant Sci. 2019, 10, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.J. Fungal cell wall chitinases and glucanases. Microbiology 2004, 150, 2029–2035. [Google Scholar] [CrossRef]
- Kappagantu, M.; Bullock, J.M.; Nelson, M.E.; Eastwell, K.C. Hop stunt viroid: Effect on host (Humulus lupulus) Transcriptome and its interactions with hop Powdery Mildew (Podospheara macularis). Mol. Plant-Microbe Interact. 2017, 30, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Viterbo, A.; Haran, S.; Friesem, D.; Ramot, O.; Chet, I. Antifungal activity of a novel endochitinase gene (chit36) from Trichoderma harzianum Rifai TM. FEMS Microbiol. Lett. 2001, 200, 169–174. [Google Scholar] [CrossRef]
- Acevedo-Garcia, J.; Kusch, S.; Panstruga, R. Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol. 2014, 204, 273–281. [Google Scholar] [CrossRef]
- Pépin, N.; Hebert, F.O.; Joly, D.L. Genome-Wide characterization of the MLO gene family in Cannabis sativa reveal two genes as strong candidates for Powdery Mildew susceptibility. Front. Plant Sci. 2021, 12, 729261. [Google Scholar] [CrossRef]
- Mihalyov, P.D.; Garfinkel, A.R. Discovery and genetic mapping of PM1, a powdery mildew resistance gene in Cannabis sativa L. Front. Agron. 2021, 3, 720215. [Google Scholar] [CrossRef]
- Goyal, N.; Bhatia, G.; Sharma, S.; Garewal, N.; Upadhyay, A.; Upadhyay, S.K.; Singh, K. Genome-wide characterization revealed role of NBS-LRR genes during powdery mildew infection in Vitis vinifera. Genomics 2020, 112, 312–322. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, encoding a typical CC-NBS-LRR protein, confers broad spectrum resistance to wheat powdery mildew disease. Mol. Plant. 2018, 11, 879–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padgitt-Cobb, L.K.; Kingan, S.B.; Henning, J.A. Genomic analysis of powdery mildew resistance in a hop (Humulus lupulus L.) bi-parental population segregating for “R6-locus”. Euphytica 2019, 216, 10. [Google Scholar]
- Gwinn, K.D.; Hansen, Z.; Kelly, H.; Ownley, B.H. Diseases of Cannabis sativa caused by diverse Fusarium Species. Front. Agron. 2022, 3, 796062. [Google Scholar] [CrossRef]
- Yulfo-Soto, G.E.; Smith, H.; Szarka, D.; Dixon, E.; Vaillancourt, L.J.; Gauthier, N. First report of Fusarium graminearum causing flower blight on hemp (Cannabis sativa) in Kentucky. Plant Dis. 2022, 106, 334. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. (Ed.) Plant Pathology, 5th ed.; Elsevier Academic Press: Burlington, MA, USA, 2005. [Google Scholar]
- Husaini, A.M.; Sakina, A.; Cambay, S.R. Host-Pathogen Interaction in Fusarium oxysporum infections: Where do we stand? Mol. Plant-Microbe Interact. 2018, 31, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Y.; Deng, G.M.; Yang, J.; Viljoen, A.; Jin, Y.; Kuang, R.B.; Zuo, C.W.; Lv, Z.C.; Yang, Q.S.; Sheng, O.; et al. Transcriptome profiling of resistant and susceptible Cavendish banana roots following inoculati on with Fusarium oxysporum f. sp. cubense tropical race 4. BMC Genom. 2012, 13, 374. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Jiang, H.; Ye, S.; Chen, W.P.; Liang, W.; Xu, Y.; Sun, B.; Sun, J.; Wang, Q.; Cohen, J.D.; et al. The Arabidopsis P450 protein CYP82C2 modulates jasmonate-induced root growth inhibition, defense gene expression and indole glucosinolate biosynthesis. Celi. Res. 2010, 20, 539–552. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Stephen, S.; Kazan, K.J.; In, G.; Fan, L.; Taylor, J.; Dennis, E.S.; Helliwell, C.A.; Wang, M.B. Characterization of the defense transcriptome responsive to Fusarium oxysporum-infection in Arabidopsis using RNA-seq. Gene 2013, 512, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.; BenRejeb, I.; Punja, Z.K.; Buirs, L.; Jabaji, S. Review: Understanding bud rot development, caused by Botrytis cinerea, on cannabis (Cannabis sativa L.) plants grown under greenhouse conditions. Botany 2023, 101, 200–231. [Google Scholar] [CrossRef]
- McPartland, J.M.; Clarke, R.C.; Watson, D.P. Hemp Diseases and Pests Management and Biological Control; CABI Publishing: Trowbridge, UK, 2000. [Google Scholar]
- van Kan, J.A. Licensed to kill: The lifestyle of a necrotrophic plant pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.M.; Simon, A.; Viaud, M. Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef] [Green Version]
- AbuQamar, S.F.; Moustafa, K.; Tran, L.S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef]
- Reboledo, G.; Agorio, A.; Vignale, L.; Batista-García, R.A.; Ponce De León, I. Botrytis cinerea transcriptome during the infection process of the Bryophyte Physcomitrium patens and angiosperms. J. Fungi 2021, 7, 11. [Google Scholar] [CrossRef]
- Balthazar, C.; Cantin, G.; Novinscak, A.; Joly, D.L.; Filion, M. Expression of putative defense responses in Cannabis primed by Pseudomonas and/or Bacillus strains and infected by Botrytis cinerea. Front. Plant Sci. 2020, 11, 572112. [Google Scholar] [CrossRef] [PubMed]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Benhamou, N.; Asselin, A.; Belanger, R. Effect of chitosan on cucumber plants: Suppression of Pythium aphanidermatum and induction of defense reactions. Am. Phytophatological. Soc. 1993, 84, 313–320. [Google Scholar] [CrossRef]
- McPartland, J.M. A review of Cannabis diseases. J. Int. Hemp. Assoc. 1996, 3, 19–23. [Google Scholar]
- Beckerman, J.; Nisonson, H.; Albright, N.; Creswell, T. First report of Pythium aphanidermatum causing crown and root rot of industrial hemp in the United States. Plant Dis. 2017, 101, 1038. [Google Scholar] [CrossRef]
- Li, X.; Han, B.; Xu, M.; Han, L.; Zhao, Y.; Liu, Z.; Dong, H.; Zhang, C. Plant growth enhancement and associated physiological responses are coregulated by ethylene and gibberellin in response to harpin protein Hpa1. Planta 2014, 239, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Mersmann, S.; Bourdais, G.; Rietz, S.; Robatzek, S. Ethylene signaling regulates accumulation of the FLS2 receptor and is required for the oxidative burst contributing to plant immunity. Plant Physiol. 2010, 154, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Amer, B.; Baidoo, E.E.K. Omics-driven biotechnology for industrial applications. Front. Bioeng. Biotechnol. 2021, 9, 613307. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Y.; Huang, L.; Cui, X.; Liu, Y. From single- to multi-omics: Future research trends in medicinal plants. Brief. Bioinform. 2023, 24, bbac485. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Holmes, J.; Collyer, D.; Lung, S. Development of tissue culture methods for marijuana (Cannabis sativa L.) strains to achieve Agrobacterium-mediated transformation to enhance disease resistance. Vitro Cell. Dev. Biol. Anim. 2019, 55, 523. [Google Scholar]
- Feeney, M.; Punja, Z.K. The role of Agrobacterium-mediated and other gene-transfer technologies in Cannabis research and product development, in Cannabis sativa L. In Botany and Biotechnology; Chandra, S., Lata, L., ElSohly, M.A., Eds.; Springer: Berlin, Germany, 2017; pp. 343–363. [Google Scholar]
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium mediated genetic transformation and CRISPR/Cas9-mediated targeted mutagenesis in Hemp (Cannabis sativa L.). Plant Biotechnol. J. 2021, 19, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.E.; Punja, Z.K. Agrobacterium-mediated transformation of THC-containing Cannabis sativa L. yields a high frequency of transgenic calli expressing bialaphos resistance and non-expressor of PR1 (NPR1) genes. Botany, 2023; in press. [Google Scholar] [CrossRef]
- Kumar, V.; Joshi, S.G.; Bell, A.A.; Rathore, K.S. Enhanced resistance against Thielaviopsis basicola in transgenic cotton plants expressing Arabidopsis NPR1 gene. Transgenic Res. 2013, 22, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Mir Zahoor, A.; Tyagi, A.; Mehari, H.; Meena, R.P.; Bhat, J.A.; Yadav, P.; Papalou, P.; Rawat, S.; Grover, A. Overexpression of NPR1 in Brassica juncea confers broad spectrum resistance to fungal pathogens. Front. Plant Sci. 2017, 8, 1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiels, D.; Prestwich, B.D.; Koo, O.; Kanchiswamy, C.N.; O’Halloran, R.; Badmi, R. Hemp genome editing—Challenges and opportunities. Front. Genome 2022, 4, 823486. [Google Scholar] [CrossRef]
- Mackintosh, C.A.; Lewis, J.; Radmer, L.E.; Shin, S.; Heinen, S.J.; Smith, L.A.; Wyckoff, M.N.; Dill-Macky, R.; Evans, C.K.; Kravchenko, S.; et al. Overexpression of defense response genes in transgenic wheat enhances resistance to Fusarium head blight. Plant Cell Rep. 2007, 26, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.J.; Kim, S.; Kim, J.S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Ahmed, S.; Gao, X.; Jahan, M.A.; Adams, M.; Wu, N.; Kovinich, N. Nanoparticle-based genetic transformation of Cannabis sativa. J. Biotechnol. 2021, 326, 48–51. [Google Scholar] [CrossRef]
- Dolgin, E. The bioengineering of Cannabis. Nature 2019, 572, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.R.; Jiang, R.; Chen, J.F.; Xiao, Y.; Lv, Z.Y.; Chen, W.S. Strategic nanoparticle-mediated plant disease resistance. Crit. Rev. Biotechnol. 2022, 43, 22–37. [Google Scholar] [CrossRef]
- Tyagi, S.; Kumar, R.; Kumar, V.; Won, S.Y.; Shukla, P. Engineering disease resistant plants through CRISPR-Cas9 technology. GM Crops Food 2021, 12, 125–144. [Google Scholar] [CrossRef]


{kind=link}
| Abbreviations | Definition |
|---|---|
| ABA | abscisic acid |
| BR | Brassinosteroids |
| CBD | Cannabidiol |
| CRISPR/Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-Associated Protein |
| ERF | Ethylene Response Factor |
| ET | Ethylene |
| ETI | Effector-Triggered Immunity |
| IGS | Inter Generic Spacer |
| ITS | Internal Transcribed Spacer |
| JA | Jasmonic Acid |
| JAZ | JA-Zim |
| LRR | Leucine-Rich Repeat |
| MAPK | Mitogen-Activated Protein Kinase |
| NBS | Nucleotide Binding Site |
| NGS | Next Generation Sequencing |
| NHR | Non Host Resistance |
| NO | Nitric Oxide |
| PAL | Phenylalanine Ammonia-Lyase |
| PAMPs | Pathogen-Associated Molecular Patterns |
| PM | Powdery Mildew |
| PR | Pathogenesis-Related protein |
| PRRs | Pattern Recognition Receptors |
| PTI | PAMP-Triggered Immunity |
| PCR | Polymerase Chain Reaction |
| PCWDEs | Plant Cell Wall Degrading Enzymes |
| R genes | Resistance genes |
| RLK | Receptor-Like Kinases |
| ROS | Reactive Oxygen Species |
| S genes | Susceptibility genes |
| SA | Salicylic Acid |
| SNP | Single Nucleotide Polymorphism |
| THC | Tetrahydrocannabinol |
| TLP | Thaumatin-Like Protein |
| Pathogen | Crop | Resistance Genes/Gene Families and Proteins | References |
|---|---|---|---|
| PM, Fusarium, Botrytis cinerea, Pythium | Cannabis | - | [12] |
| PM-Golovinomyces spp. | Hops | Genes encoding NBS proteins | [62] |
| PM-Golovinomyces spp. | Cannabis | R gene, designated as PM1 | [59] |
| PM-Golovinomyces spp. | Cannabis | Genes encoding NBS-LRR proteins | [59] |
| F. oxysporum | Arabidopsis | Genes encoding JA and P450 proteins | [68] |
| F. oxysporum | Resistant crops | Genes encoding 4-coumarate-CoA ligase, polyphenol oxidase, cellulose synthase | [67] |
| F. oxysporum | Arabidopsis | WAK gene family, genes encoding RLKs, WRKY, ERF, MYB, and NAC TFs | [69] |
| F. oxysporum | Arabidopsis | Genes encoding dirigent-like protein, CAP family and wound-responsive family proteins, some ERF TFs | [66] |
| F. oxysporum | Cannabis | WAK7 | [47] |
| Fusarium spp. | Cannabis | - | [63] |
| Botrytis cinerea | Other crops | PRs, SA, JA, ET, ABA and BR gene family | [74,75] |
| Botrytis cinerea | Cannabis | Genes involved in JA/ET, HEL, PAL, SA, PR1 and PR2 pathways | [76] |
| Pythium | Other crops | Flg22 and PTI in plants | [81] |
| Pythium | Cannabis | Harpin and Flg22 PAMPs | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirangelo, T.M.; Ludlow, R.A.; Spadafora, N.D. Molecular Mechanisms Underlying Potential Pathogen Resistance in Cannabis sativa. Plants 2023, 12, 2764. https://doi.org/10.3390/plants12152764
Sirangelo TM, Ludlow RA, Spadafora ND. Molecular Mechanisms Underlying Potential Pathogen Resistance in Cannabis sativa. Plants. 2023; 12(15):2764. https://doi.org/10.3390/plants12152764
Chicago/Turabian StyleSirangelo, Tiziana M., Richard A. Ludlow, and Natasha D. Spadafora. 2023. "Molecular Mechanisms Underlying Potential Pathogen Resistance in Cannabis sativa" Plants 12, no. 15: 2764. https://doi.org/10.3390/plants12152764