
The Effect of Ripening Stages on the Accumulation of Polyphenols and Antioxidant Activity of the Fruit Extracts of Solanum Species
 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
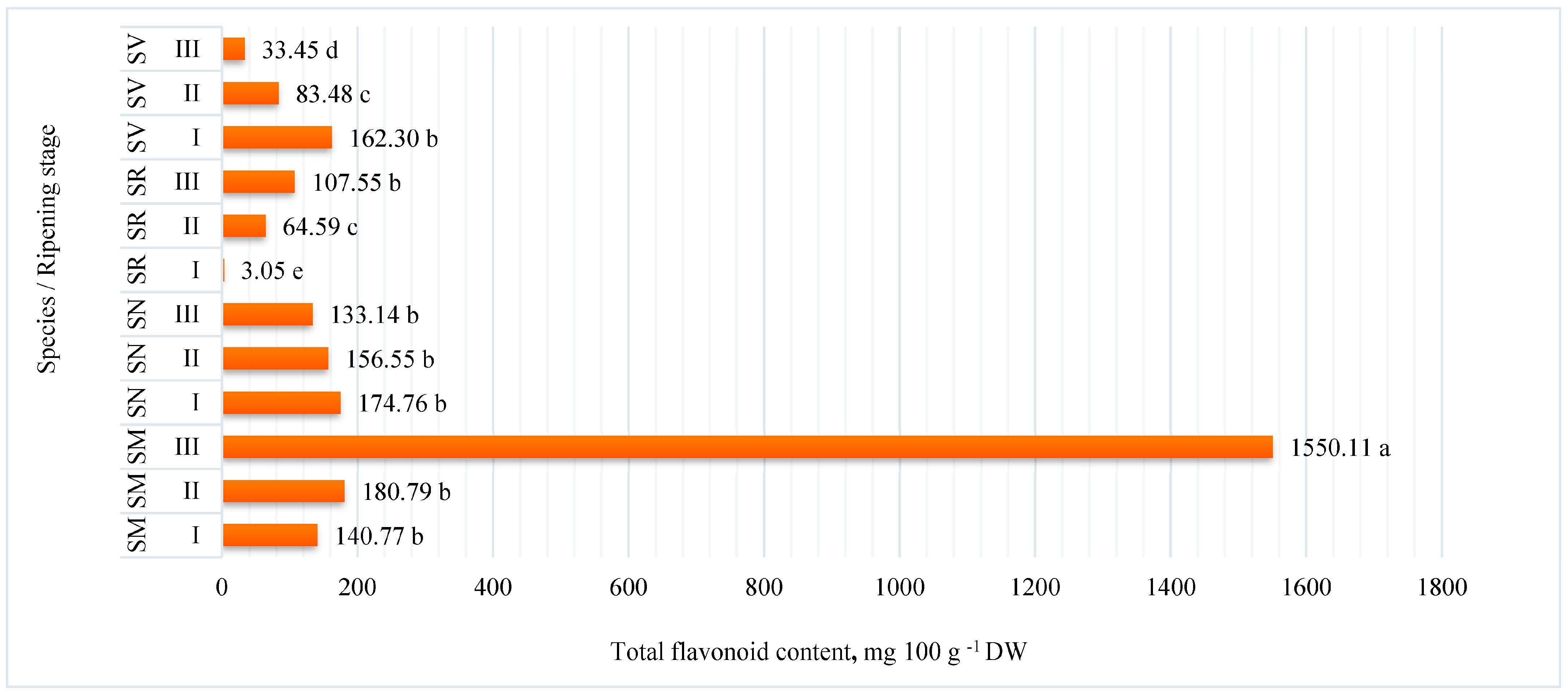
2.1. Flavonoid Content
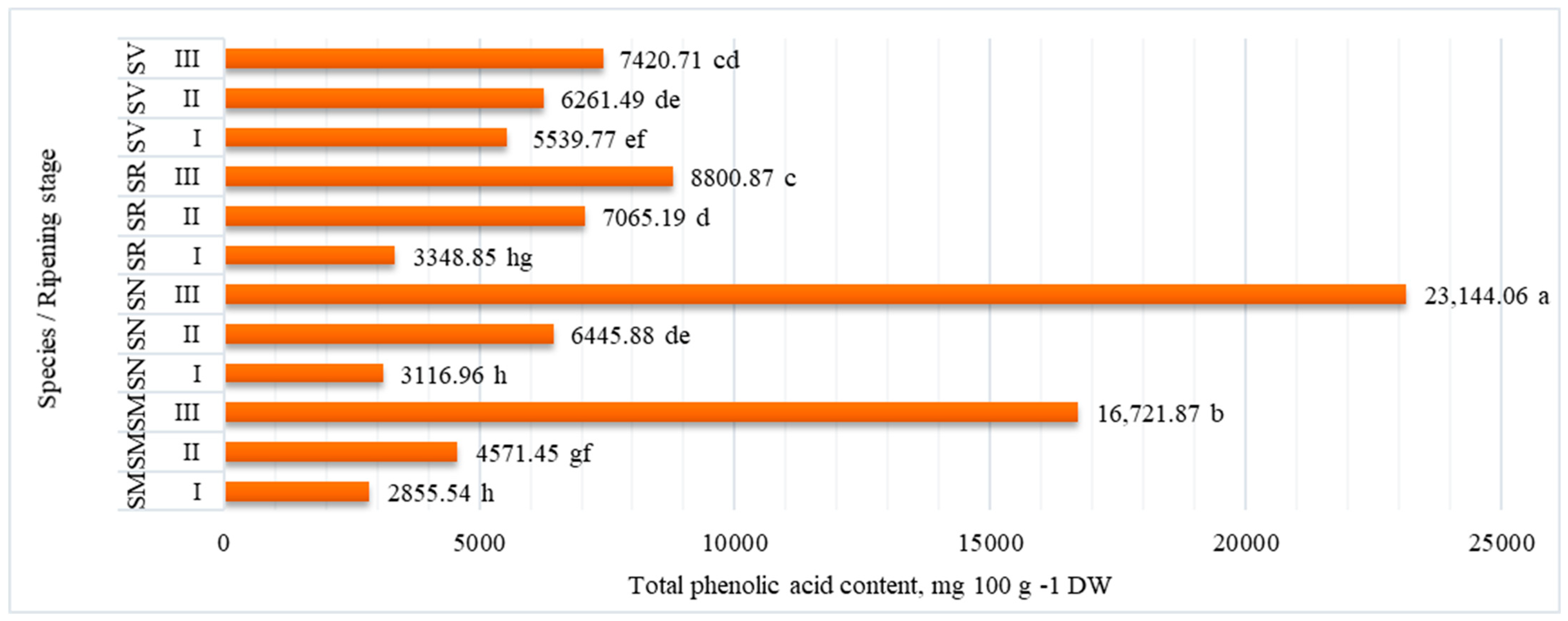
2.2. Phenolic Acid Content
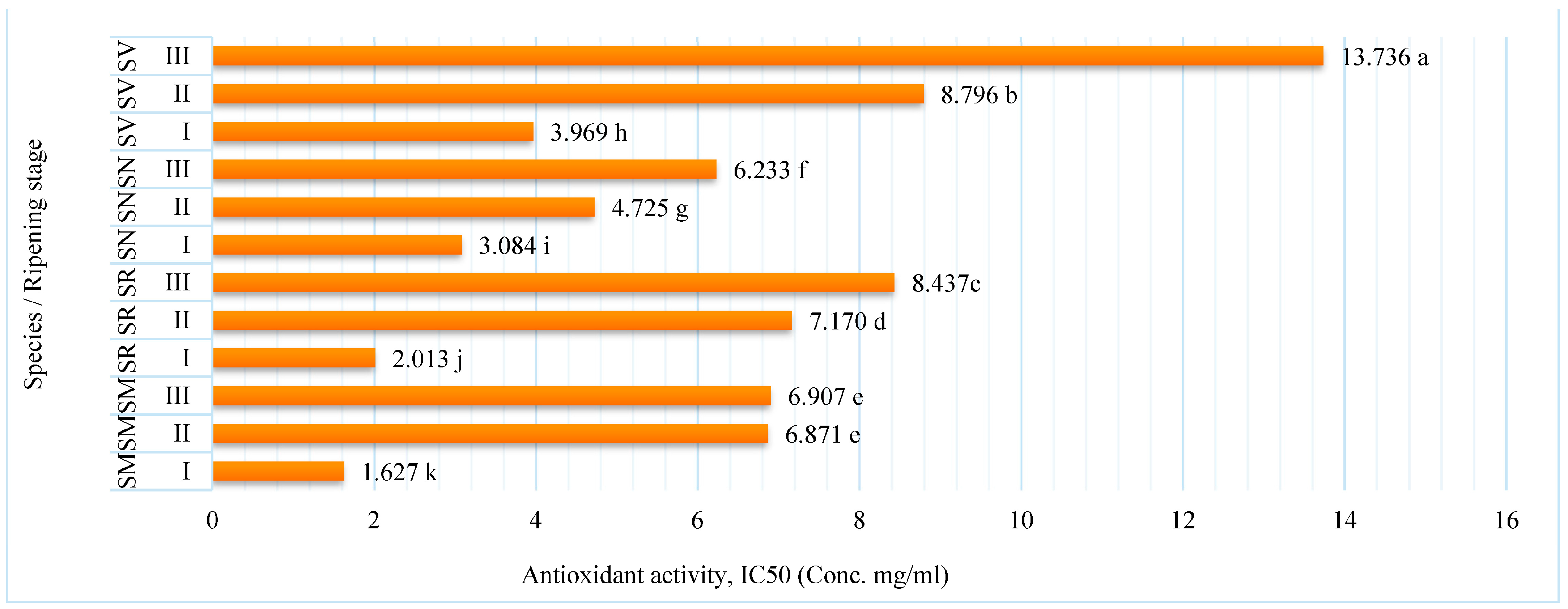
2.3. Antioxidant Activity of Fruit Extracts
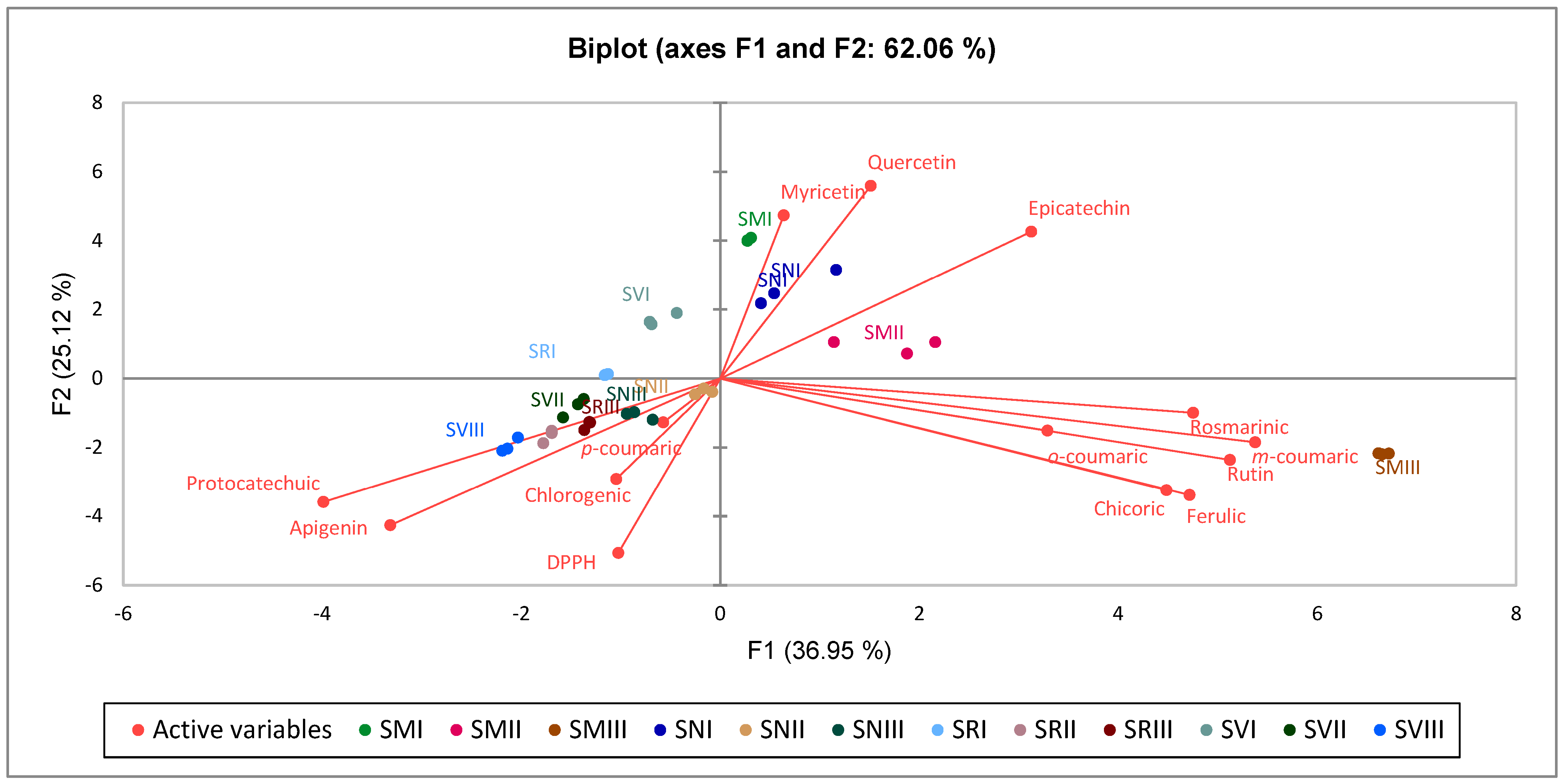
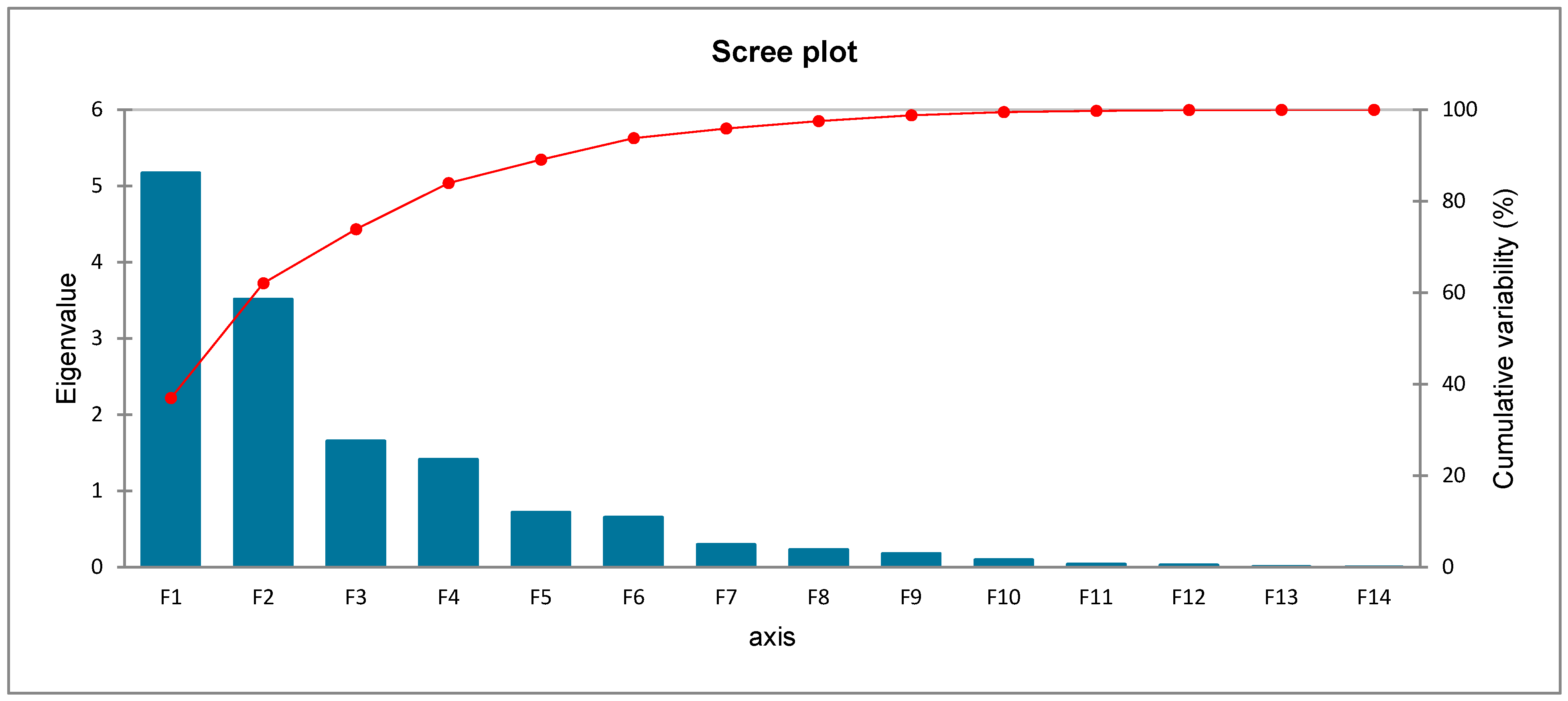
2.4. Principal Component Analysis (PCA)
3. Materials and Methods
3.1. Field Experiment
3.2. Sample Preparation
3.3. Soil Agrochemical Analyses
3.4. Meteorological Conditions
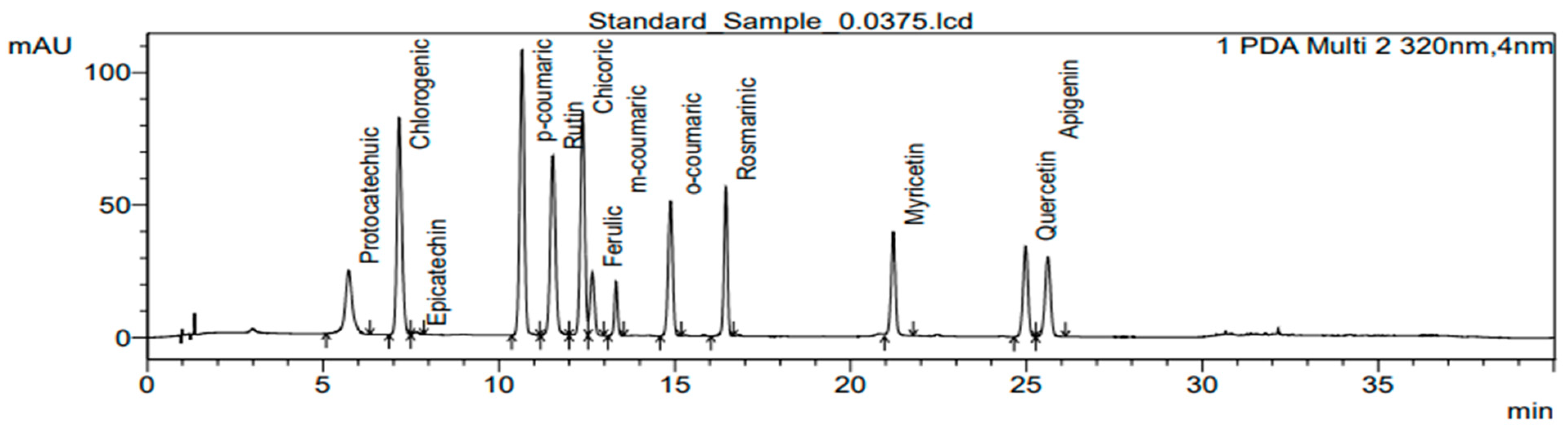
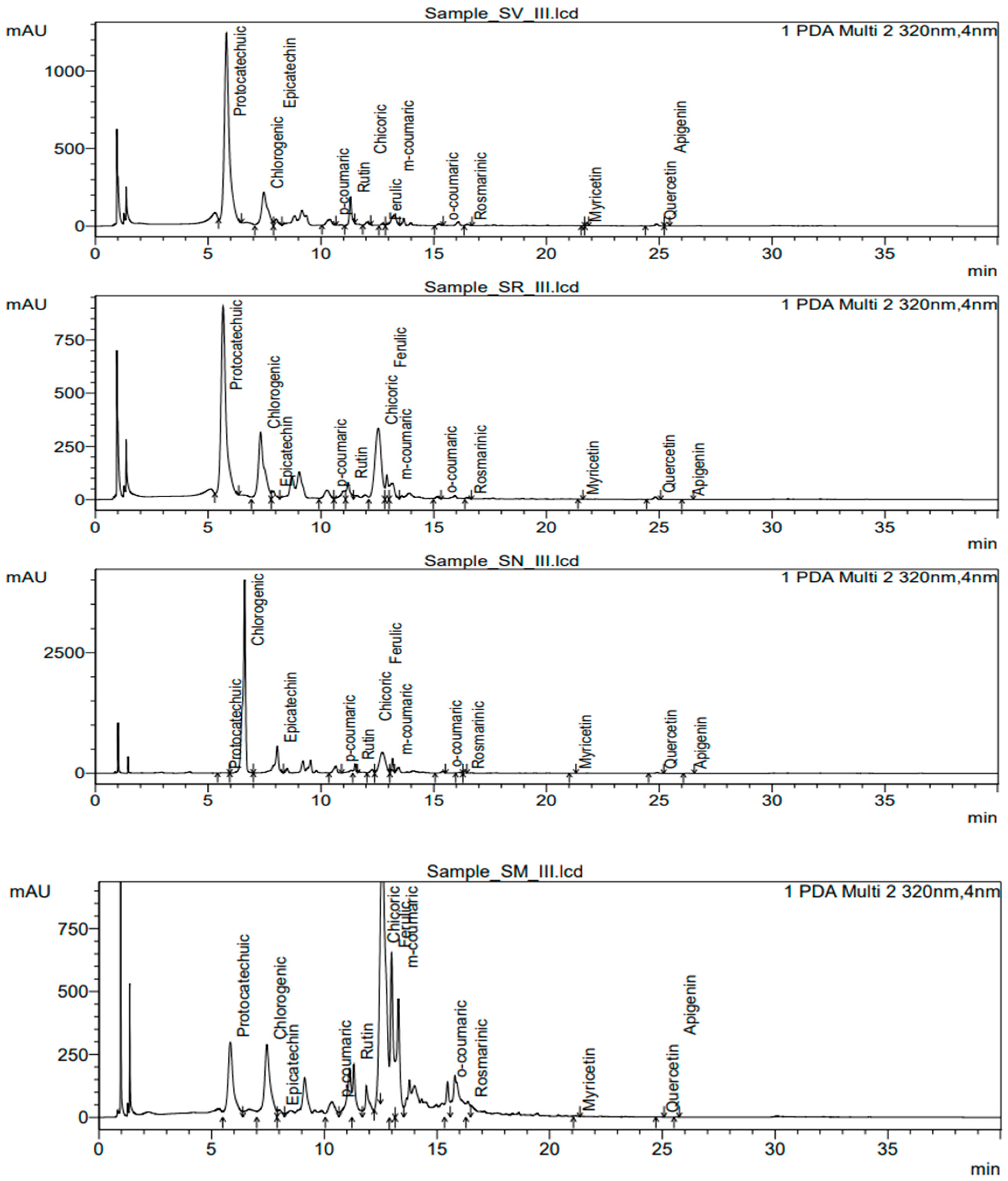
3.5. Determination of Polyphenolic Compounds
3.6. Determination of DPPH Radical Scavenging Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Knapp, S.; Bohs, L.; Nee, M.; Spooner, D.M. Solanaceae—A model for linking genomics with biodiversity. Comp. Funct. Genom. 2004, 5, 285–291. [Google Scholar]
- Kumar, P.; Kumar, J.; Kumar, R.; Dubey, R.C. Studies on phytochemical constituents and antimicrobial activities of leaves, fruits and stems of Solanum nigrum L. Asian J. Plant Sci. Res. 2016, 6, 57–68. [Google Scholar]
- Veerapagu, M.; Jeya, K.R.; Sankaranarayanan, A.; Rathika, A. In vitro antioxidant properties of methanolic extract of Solanum nigrum L. fruit. Pharma Innov. J. 2018, 7, 371–374. [Google Scholar]
- Zubair, M.; Rizwan, K.; Rasool, N.; Afshan, N.; Shahid, M.; Ahmed, V.U. Antimicrobial potential of various extract and fractions of leaves of Solanum nigrum. Int. J. Phytomed. 2011, 3, 63. [Google Scholar]
- Aly, Y.; Shallan, M.A. Antioxidant properties of wild Solanum nigrum ripe fruit. Planta Med. 2011, 77, PJ20. [Google Scholar]
- Yang, R.Y.; Ojiewo, C. African nightshades and African eggplants: Taxonomy, crop management, utilization, and phytonutrients. Discov. Chall. Chem. Health Nutr. 2013, 2, 137–165. [Google Scholar]
- Sivaraj, C.; Yamini, S.; Yahavi, A.; Praveen, K.R.; Arumugam, P.; Manimaaran, A. Antioxidant, antimicrobial activities and GCMS analysis of fruit extract of Solanum nigrum L. J. Pharmacogn. Phytochem. 2020, 9, 1114–1121. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar]
- Rathod, N.B.; Kulawik, P.; Ozogul, F.; Regenstein, J.M.; Ozogul, Y. Biological activity of plant-based carvacrol and thymol and their impact on human health and food quality. Trends Food Sci. Technol. 2021, 116, 733–748. [Google Scholar]
- Rosa-Martínez, E.; Bovy, A.; Plazas, M.; Tikunov, Y.; Prohens, J.; Pereira-Dias, L. Genetics and breeding of phenolic content in tomato, eggplant and pepper fruits. Front. Plant Sci. 2023, 14, 989. [Google Scholar]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [PubMed]
- Palma, J.M.; Corpas, F.J.; Del Río, L.A. Proteomics as an approach to the understanding of the molecular physiology of fruit development and ripening. J. Proteom. 2011, 74, 1230–1243. [Google Scholar]
- Anton, D.; Bender, I.; Kaart, T.; Roasto, M.; Heinonen, M.; Luik, A.; Püssa, T. Changes in polyphenols contents and antioxidant capacities of organically and conventionally cultivated tomato (Solanum lycopersicum L.) fruits during ripening. Int. J. Anal. Chem. 2017, 2017, 2367453. [Google Scholar] [PubMed] [Green Version]
- Kamau, E.H.; Mathara, J.M.; Kenji, G.M. Sugar Content and Physical Characterization of Four Selected African Nightshade (Solanum nigrum) Edible Berries. Eur. J. Agric. Food Sci. 2020, 2, 1–7. [Google Scholar]
- Davies, K.M.; Schwinn, K.E. Molecular biology and biotechnology of flavonoid biosynthesis. In Flavonoids: Chemistry, Biochemistry and Applications; CRC Press LLC: Boca Raton, FL, USA, 2006; pp. 143–218. [Google Scholar]
- Osorio, S.; Scossa, F.; Fernie, A.R. Molecular regulation of fruit ripening. Front. Plant Sci. 2013, 4, 198. [Google Scholar]
- Kapoor, L.; Simkin, A.J.; Priya, D.C.; Siva, R. Fruit ripening: Dynamics and integrated analysis of carotenoids and anthocyanins. BMC Plant Biol. 2022, 22, 22–27. [Google Scholar]
- Kotíková, Z.; Lachman, J.; Hejtmánková, A.; Hejtmánková, K. Determination of antioxidant activity and antioxidant content in tomato varieties and evaluation of mutual interactions between antioxidants. LWT-Food Sci. Technol. 2011, 44, 1703–1710. [Google Scholar]
- Kamau, E.H.; Mathara, J.M.; Kenji, G.M. Characterization of the Chemical and Phytochemical Profiles during Fruit Development and Ripening in Selected Cultivars of African Nightshade (Solanum Nigrum Complex) Edible Berries. J. Agric. Stud. 2020, 8, 806–819. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar]
- Dūma, M.; Alsiņa, I.; Dubova, L.; Erdberga, I. Bioactive compounds in tomatoes at different stages of maturity. Proc. Latv. Acad. Sci. Sect. B. Nat. Exact Appl. Sci. 2018, 72, 85–90. [Google Scholar]
- Auger, C.; Al-Awwadi, N.; Bornet, A.; Rouanet, J.M.; Gasc, F.; Cros, G.; Teissedre, P.L. Catechins and procyanidins in Mediterranean diets. Food Res. Int. 2004, 37, 233–245. [Google Scholar] [CrossRef]
- Chang, J.J.; Chung, D.J.; Lee, Y.J.; Wen, B.H.; Jao, H.Y.; Wang, C.J. Solanum nigrum polyphenol extracts inhibit hepatic inflammation, oxidative stress, and lipogenesis in high-fat-diet-treated mice. J. Agric. Food Chem. 2017, 65, 9255–9265. [Google Scholar] [CrossRef] [PubMed]
- Elmastaş, M.; Demir, A.; Genç, N.; Dölek, Ü.; Güneş, M. Changes in flavonoid and phenolic acid contents in some Rosa species during ripening. Food Chem. 2017, 235, 154–159. [Google Scholar]
- Zambrano-Moreno, E.L.; Chávez-Jáuregui, R.N.; Plaza, M.D.L.; Wessel-Beaver, L. Phenolic content and antioxidant capacity in organically and conventionally grown eggplant (Solanum melongena) fruits following thermal processing. Food Sci. Technol. 2015, 35, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Guofang, X.; Xiaoyan, X.; Xiaoli, Z.; Yongling, L.; Zhibing, Z. Changes in phenolic profiles and antioxidant activity in Rabbiteye blueberries during ripening. Int. J. Food Prop. 2019, 22, 320–329. [Google Scholar] [CrossRef] [Green Version]
- N’Dri, D.; Calani, L.; Mazzeo, T.; Scazzina, F.; Rinaldi, M.; Del Rio, D.; Pellegrini, N.; Brighenti, F. Effects of Different Maturity Stages on Antioxidant Content of Ivorian Gnagnan (Solanum indicum L.) Berries. Molecules 2010, 15, 7125–7138. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, M.; Bouamar, S.; Di Lorenzo, A.; Temporini, C.; Daglia, M.; Riazi, A. The Influence of Ripeness on the Phenolic Content, Antioxidant and Antimicrobial Activities of Pumpkins (Cucurbita moschata Duchesne). Molecules 2021, 26, 3623. [Google Scholar] [CrossRef]
- Meléndez, M.; Antonio, J.; Fraser-Paul, D.; Bramley-Peter, M. Accumulation of health promoting phytochemicals in wild relatives of tomato and their contribution to in vitro antioxidant activity. Phytochemistry 2010, 71, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Cano, A.; Acosta, M.; Arnao, M.B. Hydrophilic and lipophilic antioxidant activity changes during on-vine ripening of tomatoes (Lycopersicon esculentum Mill). Postharvest Biol. Technol. 2003, 1, 59–65. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Lee, J.G. Ripening-dependent changes in antioxidants, color attributes, and antioxidant activity of seven tomato (Solanum lycopersicum L.) cultivars. J. Anal. Methods Chem. 2016, 2016, 5498618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesús-Periago, M.; García-Alonso, J.; Jacob, K.; Belén-Olivares, A.; José-Bernal, M.; Dolores-Iniesta, M.; Ros, G. Bioactive compounds, folates and antioxidant properties of tomatoes (Lycopersicum esculentum) during vine ripening. Int. J. Food Sci. Nutr. 2009, 60, 694–708. [Google Scholar] [CrossRef]
- Fattahi, J.; Hamidoghli, Y.; Fotouhi, R.; Ghasemnejad, M.; Bakhshi, D. Assessment of fruit quality and antioxidant activity of three citrus species during ripening. South-West. J. Hortic. Biol. Environ. 2011, 2, 113–128. [Google Scholar]
- Park, M.H.; Sangwanangkul, P.; Baek, D.R. Changes in carotenoid and chlorophyll content of black tomatoes (Lycopersicone sculentum L.) during storage at various temperatures. Saudi J. Biol. Sci. 2018, 25, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyago-Cruz, E.; Corell, M.; Moriana, A.; Hernanz, D.; Stinco, C.M.; Mapelli-Brahm, P.; Meléndez-Martínez, A.J. Effect of regulated deficit irrigation on commercial quality parameters, carotenoids, phenolics and sugars of the black cherry tomato (Solanum lycopersicum L.) ‘Sunchocola’. J. Food Compos. Anal. 2022, 105, 104220. [Google Scholar]
- LST ISO 10390:2005; Soil Quality—Determination of pH. Lithuanian Organization for Standardization: Vilnius, Lithuania, 2005.
- Oreshkin, N. Extraction of mobile forms of phosphorus and potassium by the Egner–Riehm–Domingo method. Agrokhimiia 1980, 8, 135–138. [Google Scholar]
- Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Rasiukevičiūtė, N.; Viršilė, A.; Miliauskienė, J.; Laužikė, K.; Valiuškaitė, A.; Dėnė, L.; Chrapačienė, S.; et al. Phenolic Compounds Content Evaluation of Lettuce Grown under Short-Term Preharvest Daytime or Nighttime Supplemental LEDs. Plants 2022, 11, 1123. [Google Scholar] [CrossRef]
- Leong, L.P.; Shui, G. An investigation of antioxidant capacity of fruits in Singapore markets. Food Chem. 2022, 76, 69. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Ripening Stage | Epicatechin | Rutin | Myricetin | Quercetin | Apigenin |
|---|---|---|---|---|---|---|
| SM | I | 99.54 bc | 14.86 d | 9.54 a | 14.38 a | 2.46 f |
| SM | II | 117.05 b | 52.22 b | 4.02 b | 6.18 c | 1.32 fg |
| SM | III | 92.03 c | 1448.30 a | 3.08 bcd | 5.83 c | 0.88 g |
| SN | I | 146.35 a | 8.87 d | 3.48 bc | 15.15 a | 0.91 g |
| SN | II | 91.94 c | 47.49 b | 1.80 cd | 2.34 de | 13.00 d |
| SN | III | 69.32 d | 48.48 b | 3.850 b | 0.44 f | 11.32 e |
| SR | I | 0.85 f | 0.48 e | 1.26 d | 0.22 f | 0.23 g |
| SR | II | 7.13 f | 26.24 c | 2.97 bcd | 3.73 cde | 24.52 b |
| SR | III | 51.63 de | 30.30 c | 2.60 bcd | 1.29 de | 21.73 c |
| SV | I | 94.65 c | 52.95 b | 4.19 b | 9.98 b | 0.53 g |
| SV | II | 42.32 e | 6.88 d | 3.17 bc | 4.50 cd | 26.61 a |
| SV | III | 2.26 f | 0.71 e | 2.92 bcd | 3.66 cde | 23.90 b |
| P > F (Model) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Significant | Yes | Yes | Yes | Yes | Yes | |
| Species | Ripening Stage | P-Cumaric Acid | CHICORIC ACID | Ferulic Acid | M-Coumaric Acid | O-Coumaric Acid | Rosmarinic Acid | Protocatechuic Acid | Chlorogenic Acid |
|---|---|---|---|---|---|---|---|---|---|
| SM | I | 11.026 d | 390.05 e | 321.69 de | 296.60 c | 6.72 e | 2.77 g | 1647.94 c | 178.75 g |
| SM | II | 22.952 d | 632.62 d | 1103.89 b | 1018.20 b | 47.43 c | 39.21 bc | 1375.98 c | 331.17 f |
| SM | III | 243.64 b | 9260.75 a | 2472.87 a | 2771.45 a | 66.37 b | 79.49 a | 1394.13 c | 433.17 c |
| SN | I | 113.60 c | 143.23 fg | 172.10 f | 48.71 ef | 7.04 e | 54.74 b | 2037.70 c | 539.85 b |
| SN | II | 128.14 c | 820.32 c | 405.63 d | 136.08 d | 81.05 a | 23.38 de | 4410.50 b | 440.78 c |
| SN | III | 12,813.79 a | 3244.17 b | 566.60 c | 58.90 def | 50.30 c | 8.31 efg | 6040.11 a | 361.88 def |
| SR | I | 5.51 d | 4.28 g | 8.13 g | 38.82 ef | 28.84 d | 12.65 defg | 2735.02 c | 515.60 b |
| SR | II | 27.38 d | 766.10 cd | 620.36 c | 84.30 def | 2.01 e | 26.13 cd | 4730.28 ab | 808.64 a |
| SR | III | 35.16 d | 3198.90 b | 659.12 c | 26.75 f | 0.37 e | 6.16 fg | 4358.81 b | 515.63 b |
| SV | I | 0.51 d | 257.01 ef | 180.47 ef | 114.48 de | 3.58 e | 17.05 defg | 4550.88 b | 415.79 cd |
| SV | II | 144.91 c | 53.71 g | 659.71 c | 230.66 c | 2.54 e | 21.20 def | 4807.72 ab | 341.04 ef |
| SV | III | 150.21 c | 44.30 g | 619.35 c | 229.30 c | 0.30 e | 15.92 defg | 5968.41 a | 392.92 cde |
| P > F (Model) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Significant | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | |
| Factors | F1 | F2 |
|---|---|---|
| Eigenvalue | 5.172 | 3.516 |
| Variability (%) | 36.946 | 25.117 |
| Cumulative % | 36.946 | 62.062 |
| Factor loadings | ||
| Epicatechin | 0.541 | 0.609 |
| p-coumaric | −0.099 | −0.181 |
| Rutin | 0.887 | −0.337 |
| Chicoric acid | 0.776 | −0.462 |
| Ferulic acid | 0.816 | −0.483 |
| m-coumaric | 0.930 | −0.263 |
| o-coumaric | 0.569 | −0.215 |
| Rosmarinic acid | 0.822 | −0.142 |
| Myricetin | 0.111 | 0.676 |
| Quercetin | 0.262 | 0.797 |
| Apigenin | −0.574 | −0.608 |
| Protocatechuic acid | −0.690 | −0.511 |
| Chlorogenic acid | −0.181 | −0.416 |
| DPPH radical scavenging activity | −0.177 | −0.722 |
| Factor scores | ||
| SRI | −1.148 | 0.110 |
| SRII | −1.721 | −1.657 |
| SRIII | −1.329 | −1.342 |
| SMI | 0.287 | 4.033 |
| SMII | 1.727 | 0.943 |
| SMIII | 6.663 | −2.179 |
| SVI | −0.612 | 1.704 |
| SVII | −1.461 | −0.824 |
| SVIII | −2.121 | −1.943 |
| SNI | 0.706 | 2.599 |
| SNII | −0.166 | −0.378 |
| SNIII | −0.827 | −1.065 |
| Months | |||||||
|---|---|---|---|---|---|---|---|
| Years | May | June | July | August | September | October | |
| Air Temperature °C | Average | ||||||
| 2021 | 11.41 | 19.47 | 22.51 | 16.40 | 11.64 | 8.13 | 14.93 |
| 2022 | 11.01 | 17.58 | 17.99 | 20.86 | 11.11 | 10.07 | 14.77 |
| SCN | 13.00 | 16.30 | 18.90 | 17.80 | 12.90 | 7.20 | 14.35 |
| Rainfall, mm | Sum | ||||||
| 2021 | 121.70 | 40.30 | 48.40 | 122.20 | 29.10 | 29.80 | 391.50 |
| 2022 | 84.00 | 77.60 | 100.50 | 38.70 | 26.00 | 17.70 | 344.50 |
| SCN | 53 | 65 | 88 | 77 | 51 | 61 | 395.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staveckienė, J.; Kulaitienė, J.; Levickienė, D.; Vaitkevičienė, N.; Vaštakaitė-Kairienė, V. The Effect of Ripening Stages on the Accumulation of Polyphenols and Antioxidant Activity of the Fruit Extracts of Solanum Species. Plants 2023, 12, 2672. https://doi.org/10.3390/plants12142672
Staveckienė J, Kulaitienė J, Levickienė D, Vaitkevičienė N, Vaštakaitė-Kairienė V. The Effect of Ripening Stages on the Accumulation of Polyphenols and Antioxidant Activity of the Fruit Extracts of Solanum Species. Plants. 2023; 12(14):2672. https://doi.org/10.3390/plants12142672
Chicago/Turabian StyleStaveckienė, Jūratė, Jurgita Kulaitienė, Dovilė Levickienė, Nijolė Vaitkevičienė, and Viktorija Vaštakaitė-Kairienė. 2023. "The Effect of Ripening Stages on the Accumulation of Polyphenols and Antioxidant Activity of the Fruit Extracts of Solanum Species" Plants 12, no. 14: 2672. https://doi.org/10.3390/plants12142672