
Effects of Drought Stress on the Morphological Structure and Flower Organ Physiological Characteristics of Camellia oleifera Flower Buds
Abstract
:1. Introduction
2. Results
2.1. Effects of Drought Stress on C. oleifera Flower Bud Growth and Development
2.2. Effects of Drought Stress on the Anatomical Structure of C. oleifera Flower Buds
2.3. Effect of Drought Stress on Pollen Germination of C. oleifera
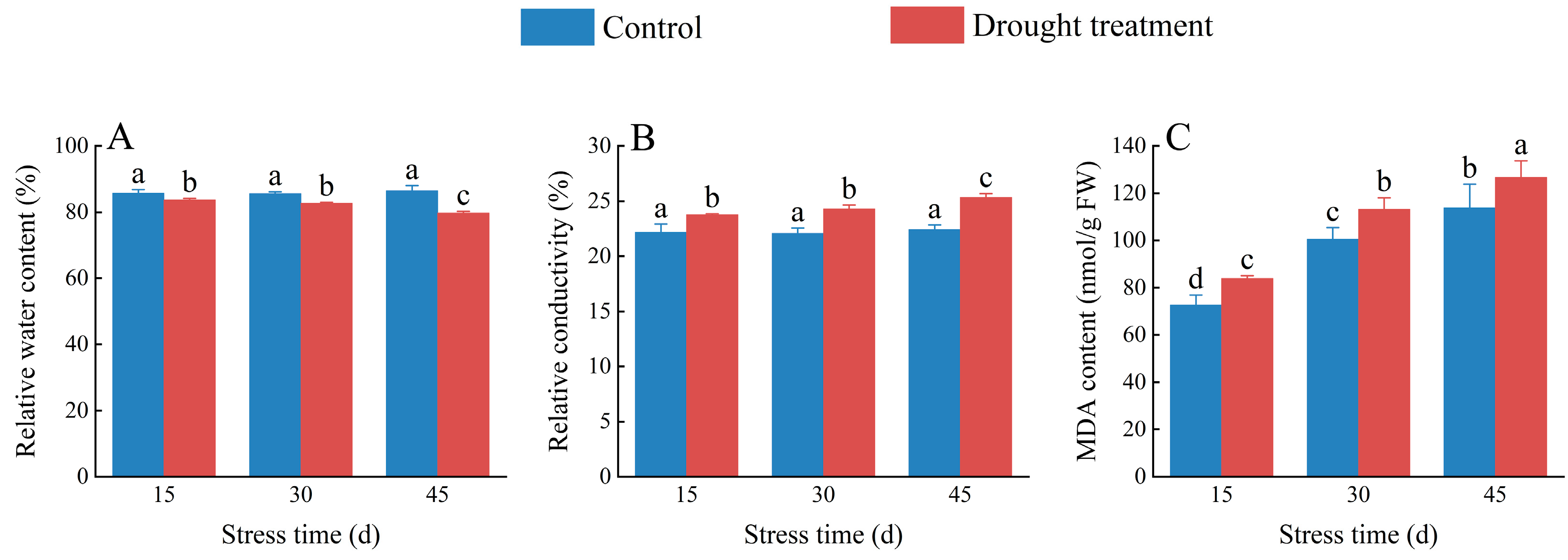
2.4. Effects of Drought Stress on the Relative Water Content, Relative Electrical Conductivity, and MDA Content in C. oleifera Petals
2.5. Effects of Drought Stress on Antioxidant Enzyme Activity and Osmoregulation Substance Content in C. oleifera Pistils and Stamens
2.6. Effects of Drought Stress on Endogenous Hormone Content in C. oleifera
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Treatments, and Sample Collection
4.2. Observation of Flower Bud Anatomical Structure
4.3. Determination of Pollen Germination
4.4. Determination of Physiological Indicators
4.4.1. Relative Water Content and Relative Conductivity
4.4.2. Antioxidant Enzymes
4.4.3. Osmoregulation Substances
4.4.4. Endogenous Hormone Content
4.5. Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rehman, B.; Javed, J.; Rauf, M.; Khan, S.A.; Arif, M.; Hamayun, M.; Gul, H.; Khilji, S.A.; Sajid, Z.A.; Kim, W.C.; et al. ACC deaminase-producing endophytic fungal consortia promotes drought stress tolerance in M. oleifera by mitigating ethylene and H2O2. Front. Plant Sci. 2022, 13, 967672. [Google Scholar] [CrossRef]
- Deng, Y.; Yan, J.; Yang, H.; Xu, L. Research progress on drought resistance of herbaceous garden plants. Seed 2017, 36, 51–57. [Google Scholar]
- La, B.; Hu, J.; Zhang, X. Advances in research on the effects of drought stress on plant physiology and the response of molecular mechanisms. Qinghai Pratacult. 2022, 31, 5. [Google Scholar]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Yu, A.; Cheng, K.; Li, H.; Tian, G.; Wang, Y.; Chen, X.; Zhang, P.; Liu, H. Changes in physiological responses of different millet cultivar to drought stress during germination and screening of drought resistance indexes. J. Agric. Sci. Technol. 2020, 22, 39–49. [Google Scholar]
- Lai, L. Physiological Response of Three Camellia Species to Drought Stress. Master’s Thesis, South China Agricultural University, Guangdong, China, 2018. [Google Scholar]
- Chen, Y. Excellent Germplasm Resources of Camellia oleifera; China Forestry Press: Beijing, China, 2008. [Google Scholar]
- Chang, Y.; Gong, W.; Xu, J.; Gong, H.; Song, Q.; Xiao, S.; Yuan, D. Integration of semi-in vivo assays and multi-omics data reveals the effect of galloylated catechins on self-pollen tube inhibition in Camellia oleifera. Hortic. Res. 2023, 10, uhac248. [Google Scholar] [CrossRef]
- Tan, X. Analysis on the cultivation and management techniques of Camellia oleifera and the development prospect of Camellia oleifera industry. Pract. Rural Technol. 2022, 243, 78–79. [Google Scholar]
- Yu, Z.; Liu, Y.; Xu, Y.; Jia, X. Research progress in reproductive biology and hybrid breeding of Camellia oleifera. Trop. Agric. Sci. 2022, 42, 44–49. [Google Scholar]
- Wei, H.; Liu, S.; Huang, M.; Li, Z. Analysis on the relationship between the yield of C. oleifera and meteorological conditions in lingyun county. Anhui Agric. Sci. 2012, 40, 16295–16296. [Google Scholar]
- Li, Y. Physiological Response and Transcriptome Analysis of C. oleifera to Drought Stress. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2021. [Google Scholar]
- Yang, Y.; Li, L.; He, H.; Ya, H.; Liu, Z. Preliminary report on the introduction and trial planting of macadamia nuts in Xiangming Township, Mengla County. Fujian Sci. Technol. Trop. Crops 2022, 47, 33–36. [Google Scholar]
- Cao, L. Physiological and Ecological Responses of Camellia oleifera to Drought Stress. Master’s Thesis, Chinese Academy of Forestry, Beijing, China, 2018. [Google Scholar]
- Feng, S. Comparative Study on Drought Resistance of Seedlings of 9 Camellia oleifera Cultivar. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2014. [Google Scholar]
- Xue, W.; Wang, L.; Li, X.; Tang, M.; Li, J.; Ding, B.; Kawabata, S.; Li, Y.; Zhang, Y. Evaluation of reference genes for quantitative pcr in eustoma grandiflorum under different experimental conditions. Horticulturae 2022, 8, 164. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Wang, X.; Gao, J.; Luo, N.; Meng, Q.; Wang, P. Dissecting the critical stage in the response of maize kernel set to individual and combined drought and heat stress around flowering. Environ. Exp Bot. 2020, 179, 104213. [Google Scholar]
- Ding, S.; Zhong, Q.; Yuan, T.; Cao, L.; Yan, C.; Yuan, Y.; Zhang, X.; Lin, J. Effects of drought stress on the growth and yield of C. oleifera flower buds. J. Nanjing For. Univ. 2017, 41, 197–202. [Google Scholar]
- Liu, L. Study on Intraspecific Variation of Flower Color and Quality Characteristics of Epimedium Coarse Hair. Master’s Thesis, Jiangxi University of Traditional Chinese Medicine, Nanchang, China, 2019. [Google Scholar]
- Ma, X.; Zeng, X.; Li, E.; Mei, H.; Luo, L.; Liu, Y. Drought tolerance of rice and its research progress. J. Shanghai Agric. 2022, 38, 36–45. [Google Scholar]
- Ascari, L.; Novara, C.; Dusio, V.; Oddi, L.; Siniscalco, C. Quantitative methods in microscopy to assess pollen viability in different plant taxa. Plant Reprod. 2020, 33, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Ma, X.; Hussain, Q.; Chen, K.; Farooq, S.; Asim, M.; Ren, X.; Shah, S.; Shi, Y. Transcriptome and anatomical studies reveal alterations in leaf thickness under long-term drought stress in tobacco. Plant Physiol. 2023, 281, 153920. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.N.A.M.; Aurora, M.I.; Diego, L.D.M.; Tomás, G.; Dolores, U.M.; Paloma, M. High temperature and water deficit cause epigenetic changes in somatic plants of Pinus radiata D. Don. Plant Cell Tissue Org. 2022, 151, 107–121. [Google Scholar]
- Huang, Y.; Deng, M.; Peng, C.; Wen, J. Study on the response of lily petal antioxidant enzyme system to drought stress. Acta Hortic. Sin. 2020, 47, 788–796. [Google Scholar]
- Liang, L.; Tang, W.; Lian, H.; Sun, B.; Huang, Z.; Sun, G.; Li, X.; Tu, L.; Li, H.; Tang, Y. Grafting promoted antioxidant capacity and carbon and nitrogen metabolism of bitter gourd seedlings under heat stress. Front. Plant Sci. 2022, 15, 1074889. [Google Scholar] [CrossRef]
- Xiong, S.; Wu, L.; Chen, Y.; Gao, M.; Jiang, X.; Li, Q.; Huang, S.; Wang, Y. Responses and evaluation of drought resistance of quercus quercus seedling leaves from different provenances to drought stress. J. Ecol. 2020, 39, 3924–3933. [Google Scholar]
- Liu, Y.; Mo, S.; He, L.; Li, M.; Shao, Y. Study on the response of different germplasms of andrographis paniculata to drought stress. Mol. Plant Breed. 2023, 1–19. [Google Scholar]
- Zhao, Y. Effects of Drought and Nitrogen Application on the Growth and Drought Resistance Physiology of C. oleifera. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2022. [Google Scholar]
- Manivannan, P.; Jaleel, C.A.; Kishorekumar, A.; Sankar, B.; Somasundaram, R.; Sridharan, R.; Panneerselvam, R. Changes in antioxidant metabolism of Vigna unguiculata (L.) Walp. by propiconazole under water deficit stress. Colloid Surf. B 2007, 57, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Hongli, C.; Wenxin, M.; Xinhui, T.; Wenhua, D. Study on the drought resistance of a new strain of cattail grass. Chin. J. Grassl. 2021, 43, 52–59. [Google Scholar]
- Liu, B.; Zhou, H.; Jia, J.; Zhang, Z.; Jin, Y. Advances in physiological research on drought resistance of forest trees in northern China. Liaoning For. Sci. Technol. 2023, 1, 46–49. [Google Scholar]
- Weidong, G.; Xiang, S.; Jiarui, L.; Xueqin, Z. Molecular mechanism of plant drought resistance. Acta Univ. Boreali-Occident. 1999, 27, 102–108. [Google Scholar]
- Ma, F.; Ma, Y. Effects of drought stress on plants and their response mechanisms. J. Ningxia Univ. Nat. Sci. Ed. 2022, 43, 391–399. [Google Scholar]
- Xu, L.; Zhang, Z.; Liang, X.; Lu, W.; Zhang, C.; Huang, F.; Wang, L.; Zhang, S. Advances in plant drought resistance genetic engineering. Chin. J. Grassl. 2014, 23, 293–303. [Google Scholar]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Tian, Y.; Fan, D.; Zhang, X. Hydraulic regulation of Populus tomentosa and Acer truncatum under drought stress. J. Plant Eco-UK 2022, 46, 1086–1097. [Google Scholar] [CrossRef]
- Bai, J.; Luo, C.; Li, X.; Gong, Y. Study on drought resistance adaptability of two wild rock plants. J. Sichuan Agric. Univ. 2005, 23, 5. [Google Scholar]
- Kang, J.; Yang, Q.; Fan, F. Effect of drought on soluble protein in alfalfa leaves. J. Grassl. 2005, 13, 199–202. [Google Scholar]
- Cao, M.; Zhao, Z.; Bi, H.; Si, G. Effects of drought on morphological and physiological characteristics of different wheat germplasms. Mol. Plant Breed. 2022, 44, 219–234. [Google Scholar]
- Tang, W. The Expression and Regulation Basis of Drought Resistant Physiological Molecules in Commonly Seen Trees in the North; Beijing Forestry University: Beijing, China, 2019. [Google Scholar]
- Guoliang, S. Study on Drought Resistance and Adaptability of Different Provenances of Pinus Softwood. Master’s Thesis, Beijing Forestry University, Beijing, China, 2009. [Google Scholar]
- Yamada, M.; Morishita, H.; Urano, K.; Shiozaki, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Yoshiba, Y. Effects of free proline accumulation in petunias under drought stress. J. Exp. Bot. 2005, 56, 1975–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; E, Z.; Sun, Y. Physiological effects of drought stress on spinach seedlings. Mod. Agric. Sci. Technol. 2023, 834, 49–51. [Google Scholar]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. N. Phytol. 2014, 203, 32–43. [Google Scholar]
- Man, D.; Bao, Y.-X.; Han, L.-B.; Zhang, X. Drought tolerance associated with proline and hormone metabolism in two tall fescue cultivars. HortScience 2011, 46, 1027–1032. [Google Scholar] [CrossRef] [Green Version]
- Jianchen, R.; Bear, D.; Europe, S.; Long, H.; Bear, X.; Junhe, Y.; Li, C.C. Effects of rhododendron mycorrhizal fungi on endogenous hormones in rhododendron persicae under continuous drought. J. Southwest Univ. Nat. Sci. Ed. 2018, 40, 26–33. [Google Scholar]
- Zhang, P.; Wang, W.-Q.; Zhang, G.-L.; Kaminek, M.; Dobrev, P.; Xu, J.; Gruissem, W. Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J. Inter. Plant Biol. 2010, 52, 653–669. [Google Scholar] [CrossRef]
- Li, W.; Yin, J.; Yue, J. GA3 effects of three treatments on chlorophyll content in grape leaves and fruit quality. Jiangsu Agric. Sci. 2019, 47, 194–197. [Google Scholar]
- Zhao, T.; Deng, X.; Xiao, Q.; Han, Y.; Zhu, S.; Chen, J. IAA priming improves the germination and seedling growth in cotton (Gossypium hirsutum L.) via regulating the endogenous phytohormones and enhancing the sucrose metabolism. Ind. Crops Prod. 2020, 155, 112788. [Google Scholar] [CrossRef]
- Liu, R.; Yang, J.; Gao, L. Changes of proline, soluble sugar, and endogenous hormone contents in leaves of chinese seabuckthorn and russian seabuckthorn under different soil water conditions. J. Soil Water Conserv. 2005, 19, 148–151. [Google Scholar]
- Li, D.; Li, C.; Sun, C.; Sun, H.; Liu, L.; Zhang, Y.; Xiao, K. Effects of drought on the content and balance of endogenous hormones in main stem and leaf of cotton. Cotton Sci. 2010, 22, 231–235. [Google Scholar]
- Alonso-Ramirez, A.; Rodriguez, D.; Reyes, D.; Jimenez, J.A.; Nicolas, G.; Lopez-Climent, M.; Gomez-Cadenas, A.; Nicolas, C. Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, E.; Zhu, X.; Hu, Y.; Liu, X.; Ruan, X. Research progress in salicylic acid and plant tolerance. J. Anhui Agric. Sci. 2021, 49, 22–24+28. [Google Scholar]
- Wang, H.; Rana, S.; Li, Z.; Geng, X.; Wang, Y.; Cai, Q.; Li, S.; Sun, J.; Liu, Z. Morphological and anatomical changes during floral bud development of the trioecious Idesia polycarpa Maxim. Braz. J. Bot. 2022, 45, 679–688. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, J.; Wang, X.; Zhao, Y. Studies on pollen morphology, pollen vitality and preservation methods of Gleditsia sinensis Lam. (Fabaceae). Forests 2023, 14, 243. [Google Scholar] [CrossRef]
- Ziyue, L.; Ruiyan, W.; Na, L.; Weihua, Z.; Lina, X. Evaluation of Cold Resistance of Four Bignoniaceae Species. Eucalyptus Sci. Technol. 2023, 40, 20–26. [Google Scholar]
- Wang, Y.C.; Qu, G.Z.; Li, H.Y.; Wu, Y.J.; Wang, C.; Liu, G.F.; Yang, C.P. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 2010, 37, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Omran, R.G. Peroxide levels and the activities of catalase, peroxidase, and indoleacetic acid oxidase during and after chilling cucumber seedlings. Plant Physiol. 1980, 65, 407–408. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Reprint of: Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hesheng, L. Experimental Principles and Techniques of Plant Physiology and Biochemistry; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Jie, J. Determination of soluble protein content in alfalfa by Coomassie brilliant blue G-250 staining. Agric. Eng. Technol. 2016, 36, 33–34. [Google Scholar]
- Wei, T.; Luo, C.; Li, H.; Li, X.; Wang, Y.; Wang, X. Effects of exogenous Methyl jasmonate on tomato seedling growth and cadmium resistance. J. Shaanxi Univ. Sci. Technol. 2021, 39, 15–20. [Google Scholar]
- Ying, Z.; Yuxiang, W.; Yongxia, L.; Guangqin, W.; Fei, N. Study on the method for determining endogenous hormones in blueberry leaves by high-performance liquid chromatography. China Fruit Tree 2018, 191, 88–91. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flower Bud Part | Treatment | Longitudinal Diameter (mm) | Lateral Diameter (mm) | ||||
| 15 Days | 30 Days | 45 Days | 15 Days | 30 Days | 45 Days | ||
| Bud | Control | 16.48 ± 1.31 bc | 17.83 ± 0.87 b | 23.43 ± 1.64 a | 6.44 ± 0.13 d | 7.68 ± 0.13 c | 9.53 ± 0.30 a |
| Drought | 12.59 ± 0.79 d | 15.34 ± 0.72 c | 17.70 ± 0.72 b | 5.39 ± 0.13 e | 6.54 ± 0.26 d | 8.10 ± 0.17 b | |
| Pistil/Stamen | Control | 7.68 ± 0.16 c | 8.41 ± 0.15 b | 9.13 ± 0.20 a | 5.21 ± 0.14 c | 6.05 ± 0.09 b | 6.65 ± 0.11 a |
| Drought | 6.57 ± 0.33 d | 7.47 ± 0.37 c | 8.35 ± 0.31 b | 4.12 ± 0.09 e | 4.54 ± 0.14 d | 5.02 ± 0.10 c | |
| Treatment | Pollen Germination Rate (%) | ||
| 15 Days | 30 Days | 45 Days | |
| Control | 83.08 ± 2.55 a | 83.24 ± 1.32 a | 83.79 ± 1.17 a |
| Drought | 47.30 ± 1.41 b | 36.04 ± 1.34 c | 22.72 ± 1.06 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.-R.; Wu, L.-L.; Wang, Y.; Liu, D.; Li, J.-A. Effects of Drought Stress on the Morphological Structure and Flower Organ Physiological Characteristics of Camellia oleifera Flower Buds. Plants 2023, 12, 2585. https://doi.org/10.3390/plants12132585
Guo P-R, Wu L-L, Wang Y, Liu D, Li J-A. Effects of Drought Stress on the Morphological Structure and Flower Organ Physiological Characteristics of Camellia oleifera Flower Buds. Plants. 2023; 12(13):2585. https://doi.org/10.3390/plants12132585
Chicago/Turabian StyleGuo, Pu-Rui, Ling-Li Wu, Ying Wang, Dan Liu, and Jian-An Li. 2023. "Effects of Drought Stress on the Morphological Structure and Flower Organ Physiological Characteristics of Camellia oleifera Flower Buds" Plants 12, no. 13: 2585. https://doi.org/10.3390/plants12132585