
The Chemical Profile, and Antidermatophytic, Anti-Inflammatory, Antioxidant and Antitumor Activities of Withania chevalieri A.E. Gonç. Ethanolic Extract

 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
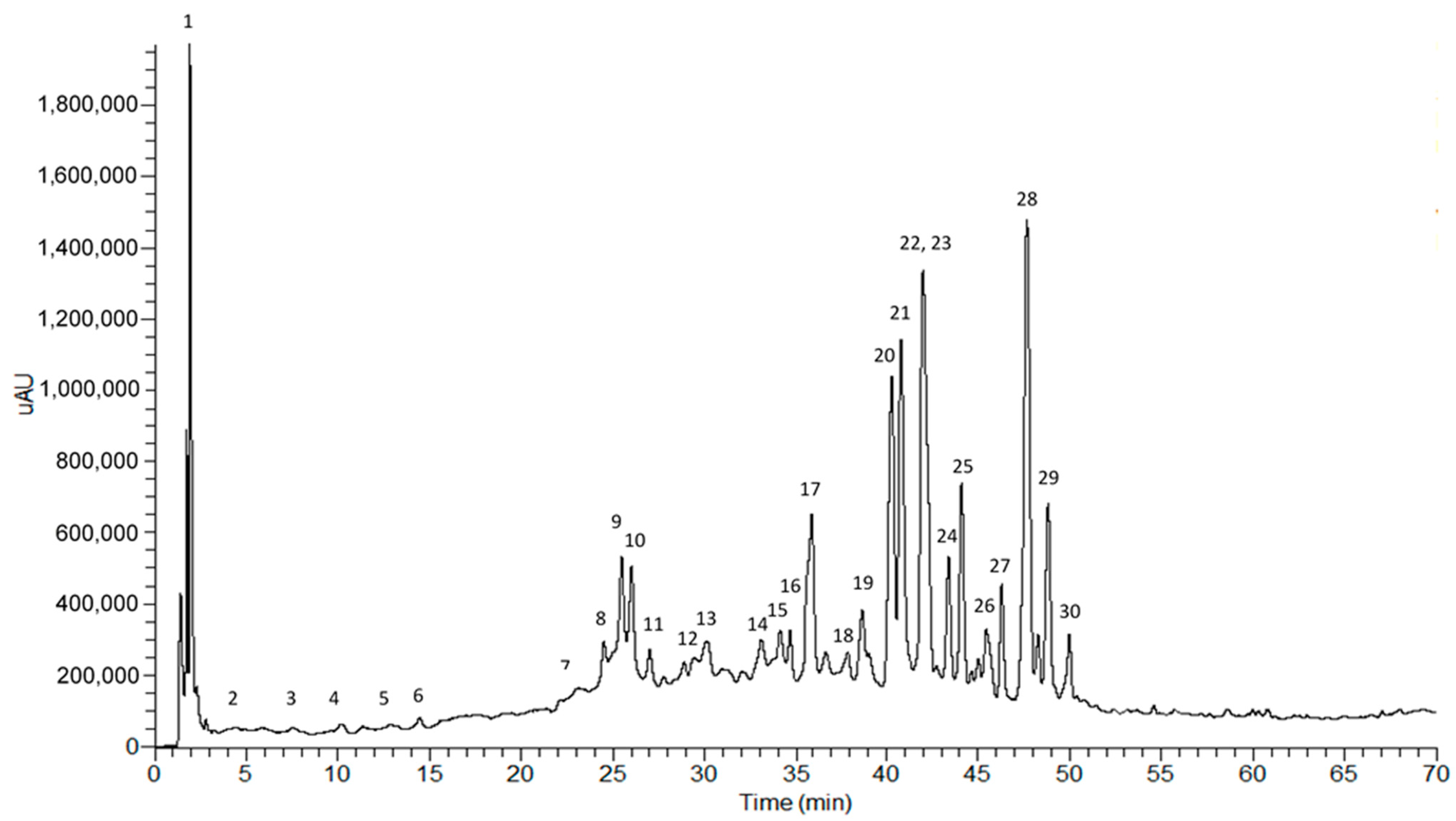
2.1. Phytochemical Profile of the Crude Ethanol Extract
2.1.1. Hydroxycinnamic Acids
2.1.2. Hydroxycinnamic Amides
2.1.3. O-Glycosylflavonoids
2.1.4. Withanolides
2.1.5. Other Compounds
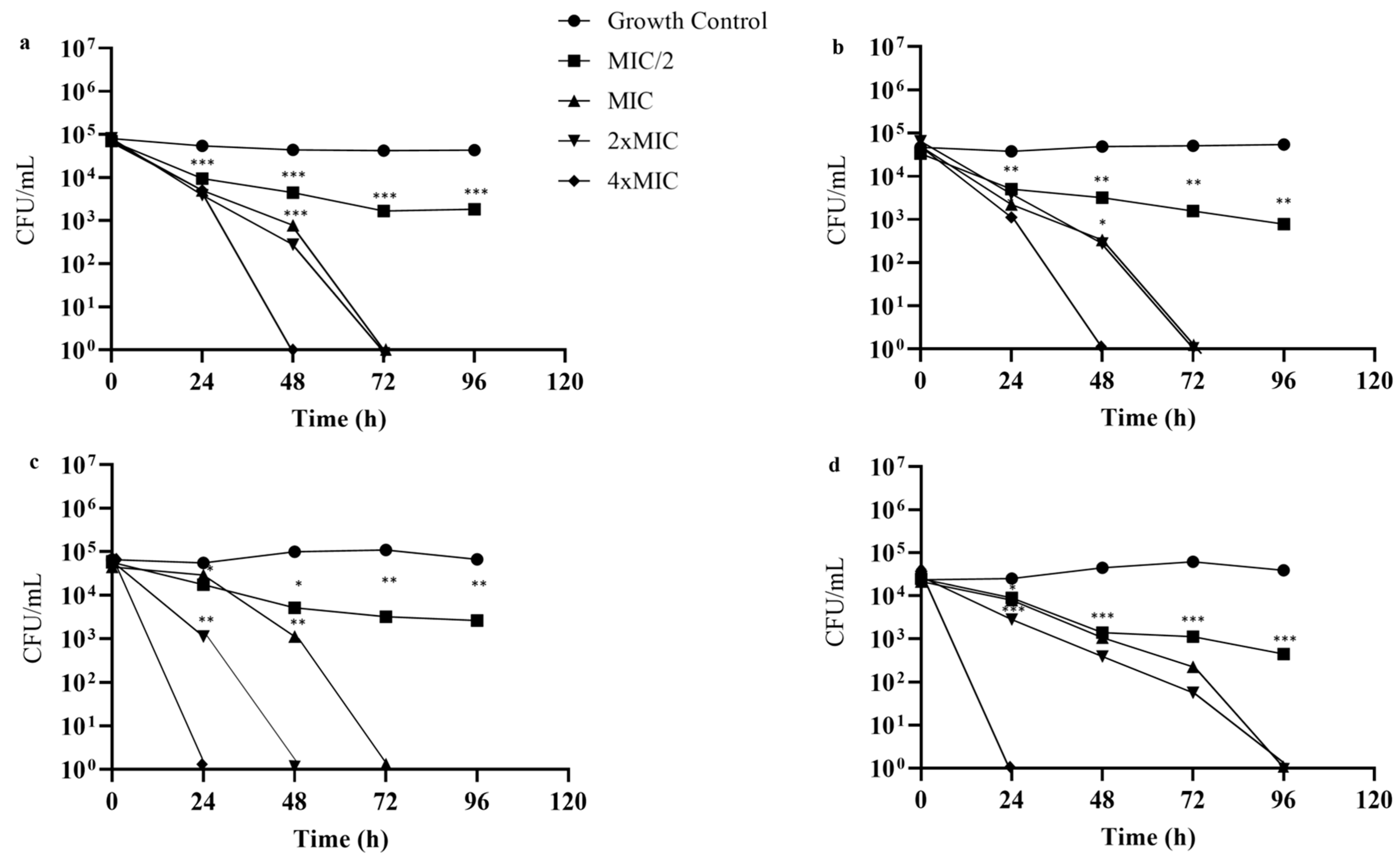
2.2. Antifungal Activity
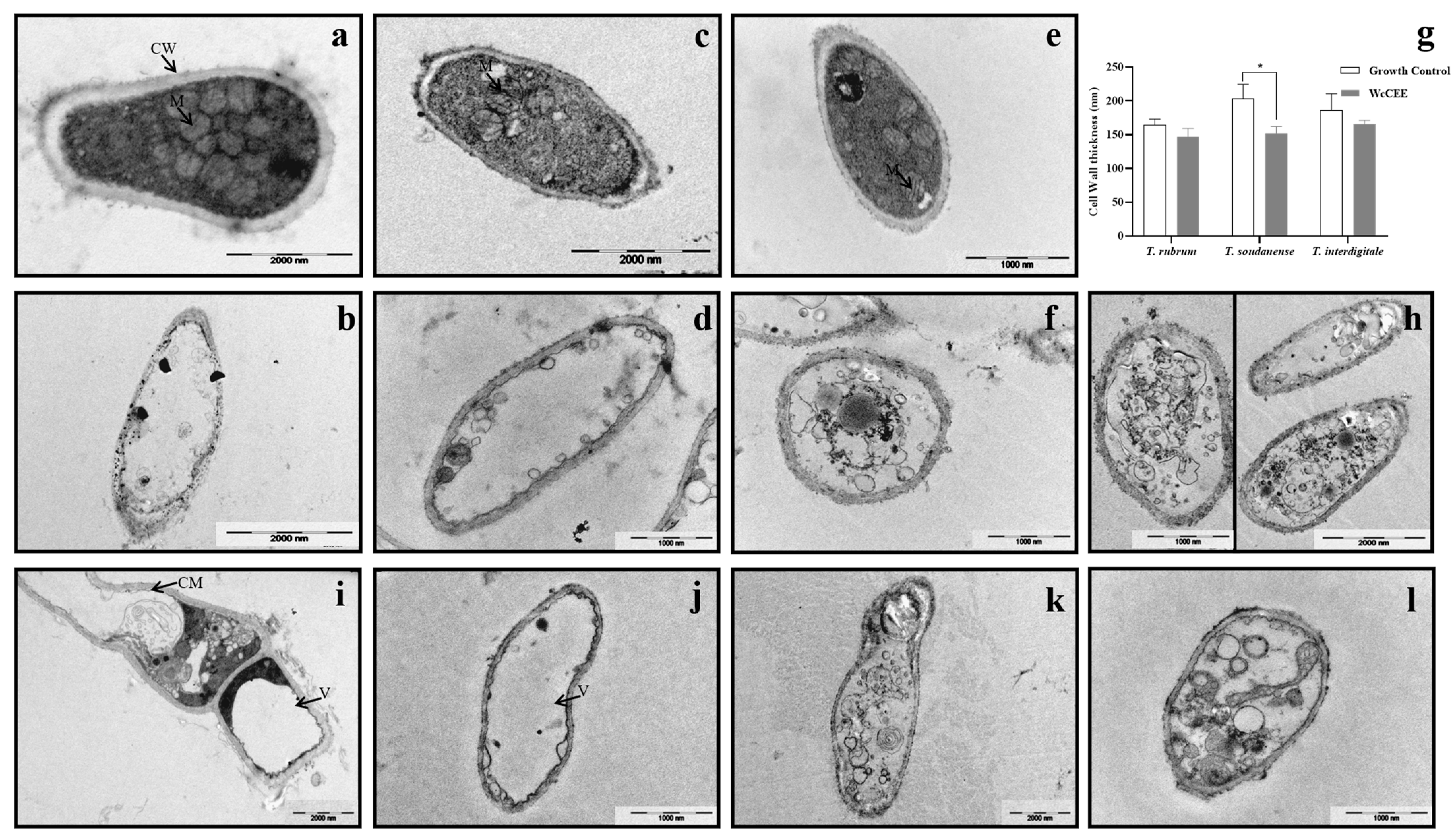
2.3. Effect of WcCEE on the Ultrastructure of Dermatophytes
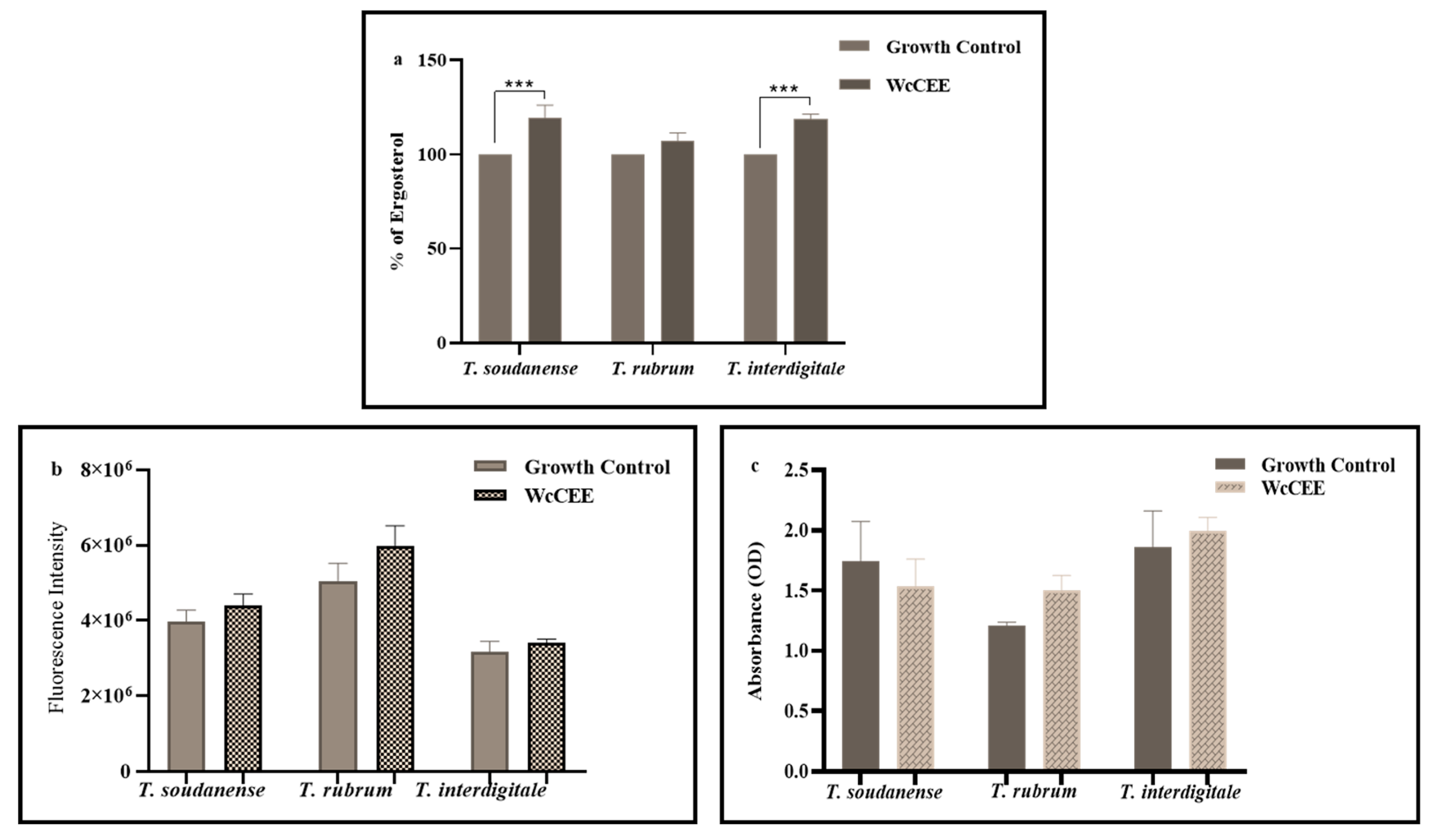
2.4. Antioxidant Activity
2.5. Anti-Inflammatory Activity
2.6. Cytotoxicity in Tumor and Non-Tumor Cells
3. Discussion
4. Materials and Methods
4.1. Extract Preparation
4.2. Dermatophyte Isolates and Fungal Culture Conditions
4.3. Chemical Characterization of the Crude Ethanolic Extract
4.4. Antifungal Susceptibility Assays
4.4.1. Preparation of Inocula and Antifungal Solutions
4.4.2. MIC and MFC Determination
4.4.3. Time-Kill Assay
4.4.4. Checkerboard Assay
4.4.5. Development of Resistance
4.5. Effect of WcCEE in Ergosterol and Cell Wall Components, and in Fungal Ultrastructure
4.6. Antioxidant Activity
4.7. Anti-Inflammatory Activity
4.8. Cytotoxicity Activity
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howes, M.-J.R.; Quave, C.L.; Collemare, J.; Tatsis, E.C.; Twilley, D.; Lulekal, E.; Farlow, A.; Li, L.; Cazar, M.-E.; Leaman, D.J.; et al. Molecules from nature: Reconciling biodiversity conservation and global healthcare imperatives for sustainable use of medicinal plants and fungi. Plants People Planet 2020, 2, 463–481. [Google Scholar] [CrossRef]
- Tabassum, N.; Hamdani, M. Plants used to treat skin diseases. Pharmacogn. Rev. 2014, 8, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.I.; Tavaria, F.K.; Pintado, M.E. Conventional and natural compounds for the treatment of dermatophytosis. Med. Mycol. 2020, 58, 707–720. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, F.; Abdullah; Ubaid, Z.; Bao, Y.; Poczai, P.; Mirza, B. Comparative Plastomics of Ashwagandha (Withania, Solanaceae) and Identification of Mutational Hotspots for Barcoding Medicinal Plants. Plants 2020, 9, 752. [Google Scholar] [CrossRef]
- Dhanani, T.; Shah, S.; Gajbhiye, N.A.; Kumar, S. Effect of extraction methods on yield, phytochemical constituents and antioxidant activity of Withania somnifera. Arab. J. Chem. 2017, 10, S1193–S1199. [Google Scholar] [CrossRef] [Green Version]
- Uddin, Q.; Samiulla, L.; Singh, V.K.; Jamil, S.S. Phytochemical and pharmacological profile of Withania somnifera dunal: A review. J. Appl. Pharm. Sci. 2012, 2, 170–175. [Google Scholar]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Ul-Haq, I.; Youn, U.J.; Chai, X.; Park, E.-J.; Kondratyuk, T.P.; Simmons, C.J.; Borris, R.P.; Mirza, B.; Pezzuto, J.M.; Chang, L.C. Biologically active withanolides from Withania coagulans. J. Nat. Prod. 2013, 76, 22–28. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Singh, S.V.; Kensler, T.W. Withania somnifera: From prevention to treatment of cancer. Mol. Nutr. Food Res. 2016, 60, 1342–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, V.K.; Peasah-Darkwah, G.; Dhasmana, A.; Jaggi, M.; Yallapu, M.M.; Chauhan, S.C. Withania somnifera: Progress towards a Pharmaceutical Agent for Immunomodulation and Cancer Therapeutics. Pharmaceutics 2022, 14, 611. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Chakraborty, S.; Anand, U.; Dey, S.; Nandy, S.; Ghorai, M.; Saha, S.C.; Patil, M.T.; Kandimalla, R.; Proćków, J.; et al. Withania somnifera (L.) Dunal (Ashwagandha): A comprehensive review on ethnopharmacology, pharmacotherapeutics, biomedicinal and toxicological aspects. Biomed. Pharmacother. 2021, 143, 112175. [Google Scholar] [CrossRef] [PubMed]
- Mikulska, P.; Malinowska, M.; Ignacyk, M.; Szustowski, P.; Nowak, J.; Pesta, K.; Szeląg, M.; Szklanny, D.; Judasz, E.; Kaczmarek, G.; et al. Ashwagandha (Withania somnifera)—Current Research on the Health-Promoting Activities: A Narrative Review. Pharmaceutics 2023, 15, 1057. [Google Scholar] [CrossRef]
- Alzahrani, A.J. Promising Antioxidant and Antimicrobial Potencies of Chemically-Profiled Extract from Withania aristata (Aiton)Pauquy against Clinically-Pathogenic Microbial Strains. Molecules 2022, 27, 3614. [Google Scholar] [CrossRef]
- Romeiras, M.M.; Catarino, L.; Torrão, M.M.; Duarte, M.C. Diversity and origin of medicinal exotic flora in Cape Verde Islands. Plant Ecol. Evol. 2011, 142, 214–225. [Google Scholar] [CrossRef] [Green Version]
- Neto, C.; Costa, J.C.; Duarte, C.; Vitória, S.; Romeiras, M.; Rivas-Martinez, S. Geobotanical excursion guide of Santiago Island (Cape Verde). Quercetea 2017, 11, 25–100. [Google Scholar]
- Catarino, S.; Duarte, M.C.; Romeiras, M.M. Withania chevalieri . In The IUCN Red List of Threatened Species; p. e.T107428968A107468392. IUCN: Glenn, Switzerland, 2017. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical Scheme for LC-MS n Identification of Chlorogenic Acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Singh, A.P.; Wang, Y.; Olson, R.M.; Luthria, D.; Banuelos, G.S.; Pasakdee, S.; Vorsa, N.; Wilson, T. LC-MS-MS Analysis and the Antioxidant Activity of Flavonoids from Eggplant Skins Grown in Organic and Conventional Environments. Food Sci. Nutr. 2017, 8, 873–888. [Google Scholar] [CrossRef] [Green Version]
- Torras-Claveria, L.; Bastida, J.; Viladomat, F.; Tiburcio, A.F. Analysis of polyamines conjugated with hydroxycinnamoyl acids by high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Methods Mol. Biol. 2018, 1694, 95–104. [Google Scholar] [CrossRef]
- Bolleddula, J.; Fitch, W.; Vareed, S.K.; Nair, M.G. Identification of metabolites in Withania somnifera fruits by liquid chromatography and high-resolution mass spectrometry. Rapid Commun. Mass. Spectrom. 2012, 26, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, D.; Gödecke, T.; Chen, S.N.; White, J.; Lankin, D.C.; Pauli, G.F.; Van Breemen, R.B. Mass spectrometric dereplication of nitrogen-containing constituents of black cohosh (Cimicifuga racemosa L.). Fitoterapia 2012, 83, 441–460. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Wang, H.; Zhao, C.; Huang, Y.Q.; Tang, X.; Cheung, H.Y. Identification and Characterization of Kukoamine Metabolites by Multiple Ion Monitoring Triggered Enhanced Product Ion Scan Method with a Triple-Quadruple Linear Ion Trap Mass Spectrometer. J. Agric. Food Chem. 2015, 63, 10785–10790. [Google Scholar] [CrossRef]
- Kachlicki, P.; Einhorn, J.; Muth, D.; Kerhoas, L.; Stobiecki, M. Evaluation of glycosylation and malonylation patterns in flavonoid glycosides during LC/MS/MS metabolite profiling. J. Mass. Spectrom. 2008, 43, 572–586. [Google Scholar] [CrossRef] [PubMed]
- Figueirinha, A.; Paranhos, A.; Pérez-Alonso, J.J.; Santos-Buelga, C.; Batista, M.T. Cymbopogon citratus leaves: Characterization of flavonoids by HPLC-PDA-ESI/MS/MS and an approach to their potential as a source of bioactive polyphenols. Food Chem. 2008, 110, 718–728. [Google Scholar] [CrossRef]
- Yang, B.Y.; Xia, Y.G.; Pan, J.; Liu, Y.; Wang, Q.H.; Kuang, H.X. Phytochemistry and biosynthesis of δ-lactone withanolides. Phytochem. Rev. 2016, 15, 771–797. [Google Scholar] [CrossRef]
- Yang, S.H.; Liu, Y.; Wang, Q.; Sun, Y.P.; Guan, W.; Liu, Y.; Yang, B.Y.; Kuang, H.X. UPLC-MS/MS Identification and Quantification of Withanolides from Six Parts of the Medicinal Plant Datura Metel L. Molecules 2020, 25, 1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, M.K.; Panda, P.; Sethi, K.K.; Jana, S. Metabolite Profiling in Withania somnifera Roots Hydroalcoholic Extract Using LC/MS, GC/MS and NMR Spectroscopy. Chem. Biodivers. 2017, 14, e1600280. [Google Scholar] [CrossRef]
- Girme, A.; Saste, G.; Pawar, S.; Balasubramaniam, A.K.; Musande, K.; Darji, B.; Satti, N.K.; Verma, M.K.; Anand, R.; Singh, R.; et al. Investigating 11 withanosides and withanolides by UHPLC-PDA and mass fragmentation studies from ashwagandha (Withania somnifera). ACS Omega 2020, 5, 27933–27943. [Google Scholar] [CrossRef]
- Ng, L.K.; Lafontaine, P.; Vanier, M. Characterization of cigarette tobacco by direct electrospray ionization-ion trap mass spectrometry (ESI-ITMS) analysis of the aqueous extract—A novel and simple approach. J. Agric. Food Chem. 2004, 52, 24. [Google Scholar] [CrossRef] [PubMed]
- Lana, A.J.D.; Pippi, B.; Carvalho, A.R.; Moraes, R.C.; Kaiser, S.; Ortega, G.G.; Fuentefria, A.M.; Silveira, G.P. In Vitro additive effect on griseofulvin and terbinafine combinations against multidrug-resistant dermatophytes. Braz. J. Pharm. Sci. 2018, 54, e17149. [Google Scholar] [CrossRef]
- Namdeo, A.G.; Sharma, A.; Yadav, K.N.; Gawande, R.; Mahadik, K.R.; Lopez-Gresa, M.P.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Metabolic characterization of Withania somnifera from different regions of India using NMR spectroscopy. Planta Med. 2011, 77, 1958–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.; Kaur, G. Aqueous extract from the Withania somnifera leaves as a potential anti-neuroinflammatory agent: A mechanistic study. J. Neuroinflam. 2016, 13, 193. [Google Scholar] [CrossRef] [Green Version]
- Jayaprakasam, B.; Zhang, Y.; Seeram, N.P.; Nair, M.G. Growth inhibition of human tumor cell lines by withanolides from Withania somnifera leaves. Life Sci. 2003, 74, 125–132. [Google Scholar] [CrossRef]
- Nefzi, A.; Aydi Ben Abdallah, R.; Jabnoun-Khiareddine, H.; Medimagh-Saïdana, S.; Haouala, R.; Daami-Remadi, M. Antifungal activity of aqueous and organic extracts from Withania somnifera L. against Fusarium oxysporum f. sp. radicis-lycopersici. J. Microb. Biochem. Technol. 2016, 8, 144–150. [Google Scholar] [CrossRef]
- Mwitari, P.G.; Ayeka, P.A.; Ondicho, J.; Matu, E.N.; Bii, C.C. Antimicrobial activity and probable mechanisms of action of medicinal plants of Kenya: Withania somnifera, Warbugia ugandensis, Prunus africana and Plectrunthus barbatus. PLoS ONE 2013, 8, e65619. [Google Scholar] [CrossRef]
- Uddandapu, P.K.; Naidu, K.C.; Rao, Y.V. In vitro anti-fungal activity of few medicinal plants of Visakhapatnam region against clinically isolated dermatophytes. Int. J. Bioassays. 2016, 5, 4573. [Google Scholar] [CrossRef]
- El Moussaoui, A.; Jawhari, F.Z.; Almehdi, A.M.; Elmsellem, H.; Fikri Benbrahim, K.; Bousta, D.; Bari, A. Antibacterial, antifungal and antioxidant activity of total polyphenols of Withania frutescens L. Bioorganic. Chem. 2019, 93, 103337. [Google Scholar] [CrossRef]
- Balkrishna, A.; Verma, S.; Mulay, V.P.; Gupta, A.; Haldar, S.; Varshney, A. Withania somnifera (L.) Dunal whole-plant extracts exhibited anti-sporotrichotic effects by destabilizing peripheral integrity of Sporothrix globosa yeast cells. PLoS Negl. Trop. Dis. 2022, 16, e0010484. [Google Scholar] [CrossRef]
- Mohsin, S.A.; Shaukat, S.; Nawaz, M.; Ur-Rehman, T.; Irshad, N.; Majid, M.; Hassan, S.S.u.; Bungau, S.; Fatima, H. Appraisal of selected ethnomedicinal plants as alternative therapies against onychomycosis: Evaluation of synergy and time-kill kinetics. Front. Pharmacol. 2022, 13, 1067697. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Chaturvedi, V.; Fothergill, A.; Rinaldi, M.G. Optimal testing conditions for determining MICs and minimum fungicidal concentrations of new and established antifungal agents for uncommon molds: NCCLS collaborative study. J. Clin. Microbiol. 2002, 40, 3776–3781. [Google Scholar] [CrossRef] [Green Version]
- Cuenca-Estrella, M. Combinations of antifungal agents in therapy--what value are they? J. Antimicrob. Chemother. 2004, 54, 854–869. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Perfect, J.R. Use of Antifungal Combination Therapy: Agents, Order, and Timing. Curr. Fungal Infect. Rep. 2010, 4, 87–95. [Google Scholar] [CrossRef] [Green Version]
- McCarty, T.P.; Pappas, P.G. Antifungal Pipeline. Front. Cell. Infect. Microbiol. 2021, 11, 732223. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Ibe, C.; Munro, C.A. Fungal cell wall: An underexploited target for antifungal therapies. PLoS Pathog. 2021, 17, e1009470. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, T.A.; Komoto, T.T.; Massaroto, B.G.; Miranda, C.E.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complement. Altern. Med. 2013, 13, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, N.A.R.; Latgé, J.P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5, 3–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, C.; Anjos, J.; Walker, L.A.; Silva, B.M.; Cortes, L.; Mota, M.; Munro, C.A.; Gow, N.A.; Gonçalves, T. Modulation of Alternaria infectoria cell wall chitin and glucan synthesis by cell wall synthase inhibitors. Antimicrob. Agents Chemother. 2014, 58, 2894–2904. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Lee, H.; Ko, H.J.; Woo, E.R.; Lee, D.G. Fungicidal effect of isoquercitrin via inducing membrane disturbance. Biochim. Biophys. Acta Biomembr. 2015, 1848, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, A.; Veses, V.; Gow, N.A.R. Vacuole dynamics in fungi. Fungal Biol. Rev. 2010, 24, 93–105. [Google Scholar] [CrossRef]
- Aki, T.; Nara, A.; Uemura, K. Cytoplasmic vacuolization during exposure to drugs and other substances. Cell Biol. Toxicol. 2012, 28, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; De, S.; Das, S.; Mukherjee, S.; Sengupta Bandyopadhyay, S. Withaferin A Induces ROS-Mediated Paraptosis in Human Breast Cancer Cell-Lines MCF-7 and MDA-MB-231. PLoS ONE 2016, 11, e0168488. [Google Scholar] [CrossRef] [Green Version]
- Laguerre, M.; Decker, E.A.; Lecomte, J.; Villeneuve, P. Methods for evaluating the potency and efficacy of antioxidants. Curr. Opin. Clin. Nutr. Metab. Care. 2010, 13, 518–525. [Google Scholar] [CrossRef]
- Phongpaichit, S.; Nikom, J.; Rungjindamai, N.; Sakayaroj, J.; Hutadilok-Towatana, N.; Rukachaisirikul, V.; Kirtikara, K. Biological activities of extracts from endophytic fungi isolated from Garcinia plants. FEMS Immunol. Med. Microbiol. 2007, 51, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget. 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Singh, S. Evaluation of anti-inflammatory effect of Withania somnifera root on collagen-induced arthritis in rats. Pharm. Biol. 2014, 52, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Cao, C.M.; Gallagher, R.J.; Timmermann, B.N. Antiproliferative withanolides from several solanaceous species. Nat. Prod. Res. 2014, 28, 1941–1951. [Google Scholar] [CrossRef] [Green Version]
- Azeem, M.; Hanif, M.; Mahmood, K.; Ameer, N.; Chughtai, F.R.S.; Abid, U. An insight into anticancer, antioxidant, antimicrobial, antidiabetic and anti-inflammatory effects of quercetin: A review. Polym. Bull. 2023, 80, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Kahlmeter, G.; Guinea, J.; Meletiadis, J.; Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). How to: Perform antifungal susceptibility testing of microconidia-forming dermatophytes following the new reference EUCAST method E.Def 11.0, exemplified by Trichophyton. Clin. Microbiol. Infect. 2021, 27, 55–60. [Google Scholar] [CrossRef]
- Van Dijck, P.; Sjollema, J.; Cammue, B.P.; Lagrou, K.; Berman, J.; d’Enfert, C.; Andes, D.R.; Arendrup, M.C.; Brakhage, A.A.; Calderone, R.; et al. Methodologies for in vitro and in vivo evaluation of efficacy of antifungal and antibiofilm agents and surface coatings against fungal biofilms. Microb. Cell 2018, 5, 300–326. [Google Scholar] [CrossRef] [PubMed]
- Hazen, K.C. Fungicidal versus fungistatic activity of terbinafine and itraconazole: An in vitro comparison. J. Am. Acad. Dermatol. 1998, 38, S37–S41. [Google Scholar] [CrossRef] [PubMed]
- Shaban, S.; Patel, M.; Ahmad, A. Improved efficacy of antifungal drugs in combination with monoterpene phenols against Candida auris. Sci. Rep. 2020, 10, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lana, D.F.; Donato, R.K.; Bündchen, C.; Guez, C.M.; Bergamo, V.Z.; de Oliveira, L.F.; Machado, M.M.; Schrekker, H.S.; Fuentefria, A.M. Imidazolium salts with antifungal potential against multidrug-resistant dermatophytes. J. Appl. Microbiol. 2015, 119, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; MacDougall, C.; Ostrosky-Zeichner, L.; Perfect, J.R.; Rex, J.H. Combination antifungal therapy. Antimicrob. Agents Chemother. 2004, 48, 693–715. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Ghannoum, M.; Isham, N.; Verma, A.; Plaum, S.; Fleischer, A., Jr.; Hardas, B. In vitro antifungal activity of naftifine hydrochloride against dermatophytes. Antimicrob. Agents Chemother. 2013, 57, 4369–4372. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.; Mota, M.; Barros, L.; Dias, M.I.; Ferreira, I.C.F.R.; Piedade, A.P.; Casadevall, A.; Gonçalves, T. Pyomelanin Synthesis in Alternaria alternata Inhibits DHN-Melanin Synthesis and Decreases Cell Wall Chitin Content and Thickness. Front. Microbiol. 2021, 12, 691433. [Google Scholar] [CrossRef] [PubMed]
- Breivik, O.N.; Owades, J.L. Yeast analysis, spectrophotometric semimicrodetermination of ergosterol in yeast. J. Agric. Food Chem. 1957, 5, 360–363. [Google Scholar] [CrossRef]
- Calheiros, D.; Dias, M.I.; Calhelha, R.C.; Barros, L.; Ferreira, I.C.F.R.; Fernandes, C.; Gonçalves, T. Antifungal Activity of Spent Coffee Ground Extracts. Microorganisms 2023, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Lockowandt, L.; Pinela, J.; Roriz, C.L.; Pereira, C.; Abreu, R.M.V.; Calhelha, R.C.; Alves, M.J.; Barros, L.; Bredol, M.; Ferreira, I.C.F.R. Chemical features and bioactivities of cornflower (Centaurea cyanus L.) capitula: The blue flowers and the unexplored non-edible part. Ind. Crops Prod. 2019, 128, 496–503. [Google Scholar] [CrossRef] [Green Version]
- Xavier, V.; Finimundy, T.C.; Heleno, S.A.; Amaral, J.S.; Calhelha, R.C.; Vaz, J.; Pires, T.C.S.P.; Mediavilla, I.; Esteban, L.S.; Ferreira, I.C.F.R.; et al. Chemical and Bioactive Characterization of the Essential Oils Obtained from Three Mediterranean Plants. Molecules 2021, 26, 7472. [Google Scholar] [CrossRef] [PubMed]
- Mandim, F.; Barros, L.; Calhelha, R.C.; Abreu, R.M.V.; Pinela, J.; Alves, M.J.; Heleno, S.; Santos, P.F.; Ferreira, I.C.F.R. Calluna vulgaris (L.) Hull: Chemical Characterization, Evaluation of Its Bioactive Properties and Effect on the Vaginal Microbiota. Food Funct. 2019, 10, 78–89. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ESI–MSn [m/z (Relative Abondance %)] | ||||||
|---|---|---|---|---|---|---|
| Peak | Rt (min) | λmax (nm) | Precursor Ion [M–H]− [M–H]+ | MS2 | MS3 | Attempt to Identify |
| 1 | 1.86 | 291 | 191 | 111 (100); 173 (33) | - | Citric acid (C6H5O7) |
| - | - | - | ||||
| 2 | 4.83 | 269 | - | - | - | Phenylalanine (C9H11NO2) |
| 166 | 120 (100) | - | ||||
| 3 | 7.88 | 291, 324 | 353 | 191(100); 179 (15); 173 (7); 161 (9) | - | 5-O-Caffeoylquinic acid (C16H18O9) |
| - | - | - | ||||
| 4 | 8.40 | 291, 324 | 367 | 193(20); 191(10); 173(100); 135(9) | - | 4-O-Feruloylquinic acid (C17H20O9) |
| - | - | - | ||||
| 5 | 13.32 | 318 | 249 | - | - | Caffeoyl putrescine (C13H18N2O3) |
| 251 | 234 (100); 163 (5); 89 (10) | 234(100), 163 (25) | ||||
| 6 | 14.30 | 287, 320 | 771 | 609 (100) | 301(100) | Quercetin-O-dihexosyl-deoxyhexoside (C33H40O21) |
| 773 | 627 (50); 611 (75); 665 (100) | 303 (100) | ||||
| 7 | 23.51 | 286, 320 | 609 | 301(100) | - | Quercetin-O-hexosyl-deoxyhexoside (C27H30O16) |
| 611 | 465(40); 303(100) | - | ||||
| 8 | 25.10 | 290, 321 | - | - | - | Feruloyl tyramine (C18H19O4N) |
| 314 | 177(100); 145(10); 117(2) | 177(80); 145(100) | ||||
| 9 | 25.32 | 254, 29 lsh, 338 | 609 | 609(50); 301(100) | 301(100), 179(20); 151(15) | Quercetin-O-hexosyl-deoxyhexoside (C27H30O16) |
| 611 | - | - | ||||
| 10 | 26.45 | 282, 323 | - | - | - | Bis(dihydrocaffeoyl)spermidine (C25H35N3O6) |
| 474 | 474 (100); 455 (15); 222 (60); 192 (5); 165 (10) | - | ||||
| 11 | 27.12 | 284, 323 | - | - | - | Methoxyferuloyltyramine (C19H21NO5) |
| 344 | 177(100); 145 (10) | |||||
| 12 | 28.72 | 274, 350 | 593 | 285 (100) | - | Luteolin-O-hexosyl-deoxyhexoside (C27H30O15) |
| 595 | 449(40); 287(100) | - | ||||
| 13 | 30.13 | 235 | 501 547 | 483 (95); 315 (100) | - | Baimantuoluoside J (C30H44O9) |
| - | - | - | ||||
| 14 | 33.10 | 230 | - | - | - | Withanoside II isomer (C40H62O16) |
| 817 [M+NH4]+ | 799 (100); 637(40); 475(45) | 475 (100) | ||||
| 15 | 34.12 | 230 | - | - | - | Withanoside II isomer (C40H62O16) |
| 817 [M+NH4]+ | 799 (100); 637(30); 475(40) | 475 (100) | ||||
| 16 | 34.67 | - | - | - | - | Grossamide (C36H37N2O8) |
| 625 | 488 (10); 462 (100); 351 (5); 325 (30) | 325 (100) | ||||
| 17 | 35.92 | 231 | 549 [M+HCOO]− | 503 (100) | 361(100); 377(80); 467(75); 343(60) | Withanolide S (C28H40O8) |
| 522 [M+NH4]+ | 505 (3); 488 (100); 319 (95); 169 (15) | - | ||||
| 18 | 36.77 | 228 | 973 | 955 (100) | 831(831) | Ashwagandhanolide (C56H78O12S) |
| 975 504 | 469 (100); 301 (30); 283 (20); 265 (50) | 451(100) | ||||
| 19 | 38.98 | 230 | 990 | 943 (100) | 781(100) | Withanolide sulfoxide (C56H78O13S) |
| 996 [M+HCOO]− 504 | 469 (100) | - | ||||
| 20 | 40.21 | 234 | 827 | 781 (100) | 781(100); 764(10); 619(15) | Withanoside IV (C40H63O15) |
| 783 800 [M+NH4]+ | 621 (40); 459 (100); 441 (80): 423 (40); 405 (20) | 441(100); 423(30); 269 (10) | ||||
| 21 | 40.89 | 234 | 827 [M+HCOO]− | 781 (100) | 781(100); 764(10); 619(15) | Withanoside VI (C40H63O15) |
| 783 800 [M+NH4]+ | 621 (40); 459 (100); 441(80): 423(40); 405(20) | 441(100); 423(30) 169(20) | ||||
| 22 | 41.93 | 233 | 515 [M+HCOO]− | 469(100) | 451(100); 433(10); 327(60) | δ-lactol withanolide (type A) (C28H38O6) |
| 453 471 [M+H]+ 488 [M+NH4]+ 493 [M+Na]+ 941 [2M+H]+ 958 [2M+NH4]+ | 471(5); 435(100); 417(40); 399(15); 325 (10) | 417(100); 399(50); 323(70) | ||||
| 23 | 42.38 | 232 | 665 [M+HCOO]− | 619(100) | 439(100) | Coagulin Q/physagulin D (C34H53O10) |
| 621 | 459(100); 441(95); 423(40); 405(20) | 441(100); 423(40); 405(20) | ||||
| 24 | 43.43 | 230 | 515 [M+HCOO]− | 469(100) | 451(100); 433(10) 327(80) | δ-lactol withanolide (type A) (C28H38O6) |
| 488 [M+NH4]+ 453 493 [M+Na]+ 941 [2M+H]+ 958 [2M+NH4]+ | 471(5); 435(100); 417(40); 399(15) | 417(100); 399(50); | ||||
| 25 | 44.19 | 231 | 469 515 [M+HCOO]− | 343 (100) | 325(55); 307(40); 299(100); 273(10) | δ-lactone withanolide (type A) (C28H38O6) |
| 453 493 [M+Na]+ 941 [2M+H]+ 958 [2M+NH4]+ | 435(75); 417(100); 399(40); 267(15) | 399(100); 381(15); 289(20); 211(15) | ||||
| 26 | 44.56 | 228 | 753 | 591(100) | 429(100) | Withanamide B/C (C38H63N2O13) |
| 755 | 593 (100) | 431 (100) | ||||
| 27 | 46.32 | 229 | 515 [M+HCOO]− | 469 (100) | 451 (90); 345 (100) | δ-lactol withanolide (type A) (C28H38O6) |
| 493 [M+Na]+ 453 488 [M+NH4]+ 941 [2M+H]+ 958 [2M+NH4]+ | 453(95); 285(100); 169(70) | 267(100); 249(30); 201(20); 157(30) | ||||
| 28 | 47.67 | 235 | - | - | - | Withanoside II aglycone (C28H43O6) |
| 475 | 475(100); 457(5); 421 (5) | - | ||||
| 29 | 48.60 | 228 | 499 [M+HCOO]− | 453 (100); 451 (20) | 435 (100); 409 (75) | Withacoagulin isomer (C28H38O5) |
| 472 [M+NH4]+ 477 [M+Na]+ 926 [2M+NH4]+ | 453 (30); 437 (75); 267 (20); 169 (15) | - | ||||
| 30 | 49.21 | 228 | 499 [M+HCOO]− | 453 (100); 451 (25) | 435 (100); 409 (65) | Withacoagulin isomer (C28H38O5) |
| 472 [M+NH4]+ 455 [M+H]+ 477 [M+Na]+ 926 [2M+NH4]+ | 437 (20); 267 (5); 169 (2) | - | ||||
| Strain ID | Species | MIC | |||||
|---|---|---|---|---|---|---|---|
| WcCEE (mg/mL) | IT (µg/mL) | TB (µg/mL) | GR (µg/mL) | ||||
| MIC | MFC | MFC/MIC | |||||
| CV3 | T. soudanense | 3.12 | 3.12 | 1 | 0.031 | 0.06 | 4 |
| CV4 | T. soudanense | 6.25 | 6.25 | 1 | 0.062 | 0.03 | 2 |
| CV8 | T. soudanense | 6.25 | 6.25 | 1 | 0.015 | 0.13 | 1 |
| CV10 | T. soudanense | 6.25 | 6.25 | 1 | 0.062 | 0.007 | 1 |
| CV11 | T. soudanense | 6.25 | 6.25 | 1 | 0.062 | 0.007 | 1 |
| CV12 | T. soudanense | 1.56 | 1.56 | 1 | 0.062 | 0.015 | 2 |
| CV15 | T. soudanense | 6.25 | 6.25 | 1 | 0.062 | 0.015 | 1 |
| CV20 | T. soudanense | 1.56 | 1.56 | 1 | 0.062 | 0.015 | 4 |
| CV24 | T. soudanense | 3.12 | 3.12 | 1 | 0.125 | 0.015 | 8 |
| CV30 | T. soudanense | 6.25 | 6.25 | 1 | 0.031 | 0.007 | 0.5 |
| CV42 | T. soudanense | 6.25 | 6.25 | 1 | 0.003 | 0.007 | 1 |
| CV45 | T. soudanense | 6.25 | 6.25 | 1 | 0.031 | 0.007 | 4 |
| CV47 | T. soudanense | 6.25 | 6.25 | 1 | 0.031 | 0.007 | 4 |
| CV50 | T. soudanense | 3.12 | 3.12 | 1 | 0.031 | 0.007 | 0.5 |
| CV52 | T. soudanense | 1.56 | 3.12 | 1 | 0.031 | 0.007 | 2 |
| CV 54 | T. soudanense | 3.12 | 3.12 | 1 | 0.015 | 0.003 | 4 |
| CV58 | T. soudanense | 6.25 | 6.25 | 1 | 0.125 | 0.015 | 2 |
| CV60 | T. soudanense | 6.25 | 6.25 | 1 | 0.015 | 0.125 | 2 |
| Esp16 | T. rubrum | 6.25 | 6.25 | 1 | 0.062 | 0.06 | 1 |
| Esp32 | T. rubrum | 6.25 | 6.25 | 1 | 0.125 | 0.02 | 1 |
| IMF-29 | T. rubrum | 3.12 | 6.25 | 2 | 0.25 | 0.015 | 1 |
| CV55 | T. rubrum | 6.25 | 6.25 | 1 | 0.007 | 0.03 | 4 |
| Esp14 | T. interdigitale | 6.25 | 6.25 | 1 | 0.031 | 0.06 | 0.5 |
| IMF-28 | T. interdigitale | 3.12 | 6.25 | 2 | 0.125 | 0.13 | 2 |
| IMF-35 | M. canis | 25 | 25 | 1 | 0.25 | 0.5 | 2 |
| PT01 | M. canis | 25 | 25 | 1 | 0.25 | 0.5 | 2 |
| MIC (mg/mL) and (μg/mL) | MIC of WcCEE (mg/mL) + IT (μg/mL) | MIC of WcCEE (mg/mL) + TB (μg/mL) | MIC of WcCEE (mg/mL) + GR (μg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain ID | Species | WcCEE | IT | TB | GR | WcCEE | IT | FICI | Observ. | WcCEE | TB | FICI | Observ. | WcCEE | GR | FICI | Observ. |
| CV8 | T. soudanense | 6.25 | 0.015 | 0.13 | 1 | 0.097 | 0.007 | 0.48 | SYN. * | 0.097 | 0.001 | 0.024 | SYN. | 1.56 | 0.125 | 0.375 | SYN. |
| CV4 | T. soudanense | 6.25 | 0.062 | 0.03 | 2 | 1.56 | 0.031 | 0.75 | ADD. ** | 0.781 | 0.007 | 0.351 | SYN. | 0.781 | 1 | 0.625 | ADD. |
| CV3 | T. soudanense | 3.12 | 0.031 | 0.06 | 4 | 0.097 | 0.007 | 0.26 | SYN. | 0.097 | 0.015 | 0.273 | SYN. | 0.39 | 1 | 0.375 | SYN. |
| Esp14 | T. interdigitale | 6.25 | 0.031 | 0.06 | 0.5 | 0.097 | 0.007 | 0.24 | SYN. | 0.097 | 0.015 | 0.257 | SYN. | 3.12 | 0.125 | 0.749 | ADD. |
| IMF28 | T. interdigitale | 3.12 | 0.125 | 0.13 | 2 | 1.56 | 0.062 | 1 | ADD. | 0.781 | 0.031 | 0.498 | SYN. | 1.56 | 1 | 1 | ADD. |
| Esp32 | T. rubrum | 6.25 | 0.125 | 0.02 | 1 | 0.097 | 0.062 | 0.51 | ADD. | 0.097 | 0.003 | 0.216 | SYN. | 0.097 | 0.5 | 0.516 | ADD. |
| Esp16 | T. rubrum | 6.25 | 0.062 | 0.06 | 1 | 0.195 | 0.062 | 1.03 | ADD. | 0.39 | 0.015 | 0.304 | SYN. | 1.56 | 1 | 1.25 | ADD. |
| CV55 | T. rubrum | 6.25 | 0.007 | 0.03 | 4 | 0.097 | 0.003 | 0.44 | SYN. | 0.097 | 0.007 | 0.241 | SYN. | 3.12 | 0.5 | 0.624 | ADD. |
| IMF35 | M. canis | 25 | 0.25 | 0.5 | 2 | 25 | 0.25 | 2 | ADD. | 6.25 | 0.062 | 0.374 | SYN. | 25 | 2 | 2 | ADD. |
| PT01 | M. canis | 25 | 0.25 | 0.5 | 2 | 6.25 | 0.25 | 1.25 | ADD. | 0.097 | 0.125 | 0.254 | SYN. | 25 | 2 | 2 | ADD. |
| MIC (mg/mL) | ||||
|---|---|---|---|---|
| Isolate | Initial | 5 Passages | 10 Passages | 15 Passages |
| T. rubrum CV55 | 6.25 | 6.25 | 6.25 | 6.25 |
| T. interdigitale Esp14 | 6.25 | 6.25 | 6.25 | 12.5 * |
| T. soudanense CV3 | 6.25 | 6.25 | 6.25 | 6.25 |
| WcCEE | Positive Control | |
|---|---|---|
| Antioxidant activity | ||
| TBARS (IC50, mg/mL) | 2.1 ± 0.2 | 0.0091 ± 0.0003 |
| OxHLIA (IC50, mg/mL) | 0.49 ± 0.03 | 0.0218 ± 0.0003 |
| CAA (% inhibition) | 60 ± 5 | 95 ± 5 |
| Anti-inflammatory activity (IC50, µg/mL) | ||
| NO production inhibition | 7 ± 1 | 6.3±0.4 |
| Cytotoxic activity (GI50, µg/mL) | ||
| AGS | 47 ± 4 | 1.20 ± 0.03 |
| CaCo2 | 63 ± 4 | 1.20 ± 0.02 |
| MCF-7 | 27 ± 2 | 1.00 ± 0.02 |
| NCI-H460 | 19 ± 1 | 1.20± 0.02 |
| Vero | >400 | 1.40 ± 0.06 |
| PLP2 | >400 | 1.4 ± 0.1 |
| Code | Species | Anatomical Site | Origin |
|---|---|---|---|
| CV3 | T. soudanense | Body | Cape Verde |
| CV4 | T. soudanense | Hand | Cape Verde |
| CV8 | T. soudanense | Nail | Cape Verde |
| CV10 | T. soudanense | Leg | Cape Verde |
| CV11 | T. soudanense | Body | Cape Verde |
| CV12 | T. soudanense | Arm | Cape Verde |
| CV15 | T. soudanense | Face | Cape Verde |
| CV20 | T. soudanense | Body | Cape Verde |
| CV24 | T. soudanense | Leg | Cape Verde |
| CV30 | T. soudanense | Body | Cape Verde |
| CV42 | T. soudanense | Arm | Cape Verde |
| CV45 | T. soudanense | Body | Cape Verde |
| CV47 | T. soudanense | Body | Cape Verde |
| CV50 | T. soudanense | Toenail | Cape Verde |
| CV52 | T. soudanense | Hand | Cape Verde |
| CV54 | T. soudanense | Nail | Cape Verde |
| CV58 | T. soudanense | Face | Cape Verde |
| CV60 | T. soudanense | Scalp | Cape Verde |
| Esp16 | T. rubrum | Nail | Spain |
| Esp32 | T. rubrum | Nail | Spain |
| IMF-29 | T. rubrum | Nk | Portugal |
| CV55 | T. rubrum | Nail | Cape Verde |
| Esp14 | T. interdigitale | Nail | Spain |
| IMF-28 | T. interdigitale | Nk | Portugal |
| IMF-35 | M. canis | Nk | Portugal |
| PT01 | M. canis | Arm | Portugal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, E.E.M.; Figueirinha, A.; Rodrigues, L.; Pinela, J.; Calhelha, R.C.; Barros, L.; Fernandes, C.; Salgueiro, L.; Gonçalves, T. The Chemical Profile, and Antidermatophytic, Anti-Inflammatory, Antioxidant and Antitumor Activities of Withania chevalieri A.E. Gonç. Ethanolic Extract. Plants 2023, 12, 2502. https://doi.org/10.3390/plants12132502
Correia EEM, Figueirinha A, Rodrigues L, Pinela J, Calhelha RC, Barros L, Fernandes C, Salgueiro L, Gonçalves T. The Chemical Profile, and Antidermatophytic, Anti-Inflammatory, Antioxidant and Antitumor Activities of Withania chevalieri A.E. Gonç. Ethanolic Extract. Plants. 2023; 12(13):2502. https://doi.org/10.3390/plants12132502
Chicago/Turabian StyleCorreia, Edmilson Emanuel Monteiro, Artur Figueirinha, Lisa Rodrigues, José Pinela, Ricardo C. Calhelha, Lillian Barros, Chantal Fernandes, Lígia Salgueiro, and Teresa Gonçalves. 2023. "The Chemical Profile, and Antidermatophytic, Anti-Inflammatory, Antioxidant and Antitumor Activities of Withania chevalieri A.E. Gonç. Ethanolic Extract" Plants 12, no. 13: 2502. https://doi.org/10.3390/plants12132502