
Effects of Drought Stress on Non-Structural Carbohydrates in Different Organs of Cunninghamia lanceolata
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effects of Drought Stress on NSCs in Aboveground Organs of Chinese Fir Plantlets
2.2. Effects of Drought Stress on NSCs in Belowground Organs of Chinese Fir Plantlets
2.3. Correlations between Soluble Sugar, Starch, and NSC Content in Different Organs of Chinese Fir Plantlets
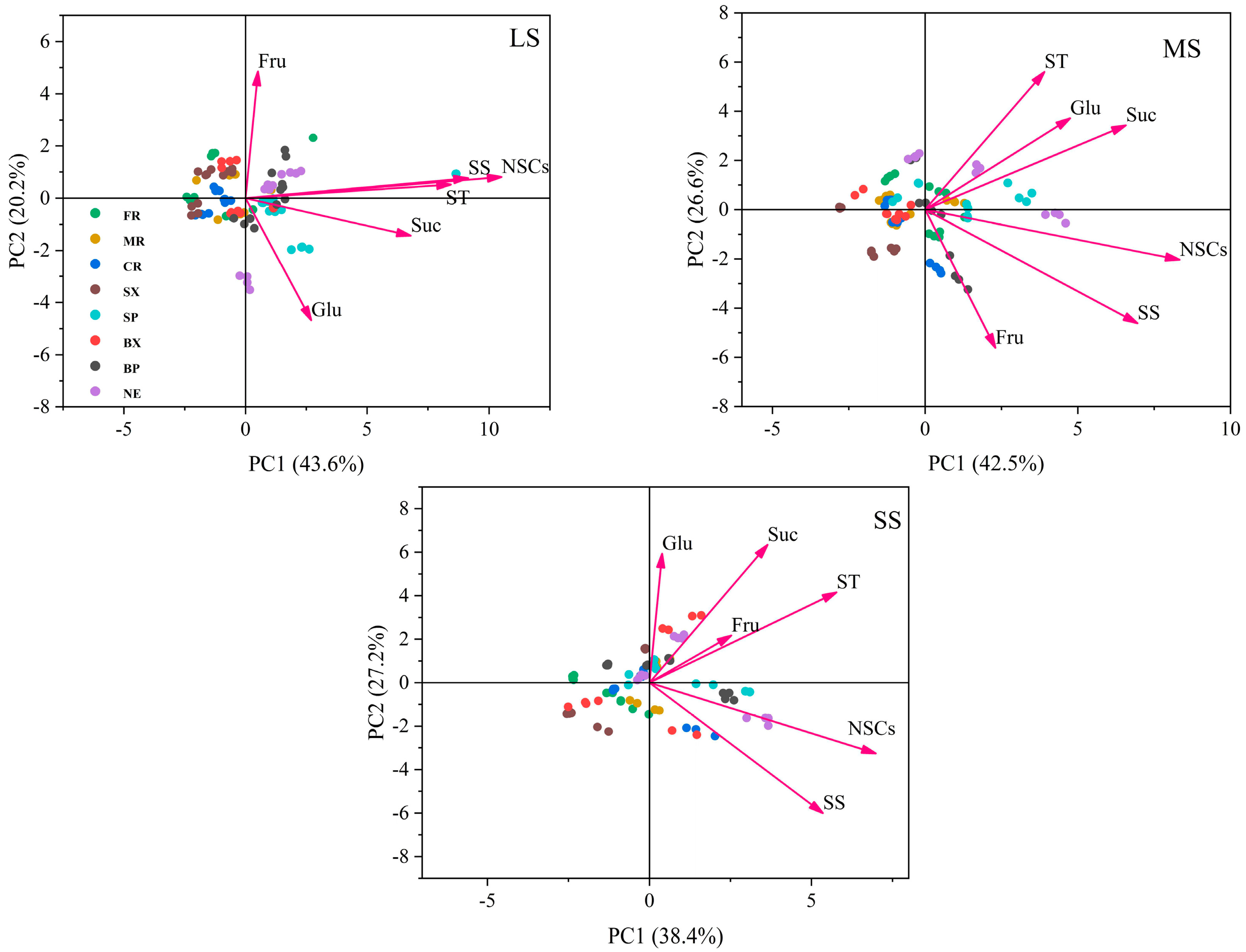
2.4. Principal Component Analysis (PCA) of NSCs in Different Organs of Chinese Fir
3. Discussion
3.1. Effects of Drought Stress on NSCs Distribution in Aboveground Organs
3.2. Effects of Drought Stress on NSCs Distribution in Belowground Roots
3.3. How Does NSCs Distribution in Different Organs Helps Alleviate Drought Stress?
4. Materials and Methods
4.1. Plant Material and Samplings
4.2. Measurement of Non-Structural Carbohydrate Content
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Auler, P.A.; Souza, G.M.; da Silva Engela, M.R.G.; do Amaral, M.N.; Rossatto, T.; da Silva, M.G.Z.; Braga, E.J.B. Stress memory of physiological, biochemical and metabolomic responses in two different rice genotypes under drought stress: The scale matters. Plant Sci. 2021, 311, 110994. [Google Scholar] [CrossRef]
- Abreha, K.B.; Enyew, M.; Carlsson, A.S.; Vetukuri, R.R.; Feyissa, T.; Motlhaodi, T.; Geleta, M. Sorghum in dryland: Morphological, physiological, and molecular responses of sorghum under drought stress. Planta 2022, 255, 20. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Lloret, F.; Martinez-Vilalta, J.; Serra-Diaz, J.M.; Ninyerola, M. Relationship between projected changes in future climatic suitability and demographic and functional traits of forest tree species in Spain. Clim. Chang. 2013, 120, 449–462. [Google Scholar] [CrossRef]
- Zhang, T.; Cao, Y.; Chen, Y.M.; Liu, G.B. Non-structural carbohydrate dynamics in Robinia pseudoacacia saplings under three levels of continuous drought stress. Trees 2015, 29, 1837–1849. [Google Scholar] [CrossRef]
- Sharma, A.; Zheng, B. Melatonin mediated regulation of drought stress: Physiological and molecular aspects. Plants 2019, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.T.; Bu, J.W.; Zhang, S.Y.; Jing, G.; Zhang, G.C.; Liu, X. Effects of drought stress on the photosynthetic physiological parameters of Populus × euramericana “Neva”. J. For. Res. 2019, 30, 409–416. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortalit. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhong, Y.Q.W.; Shangguan, Z.P. Elevated temperature exacerbates the effects of drought on the carbon and hydraulic characteristics of Robinia pseudoacacia seedlings. Agr. For. Meteorol. 2020, 280, 107794. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wu, F.; Li, M.; Liang, D.Q.; Ding, G.J. Heteroblastic Foliage Affects the Accumulation of Non-Structural Carbohydrates and Biomass in Pinus massoniana (Lamb.) Seedlings. Forests 2021, 12, 1686. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, G.; Peng, S.; Bai, J.; Lu, Q.; Duan, B. Water relations and non-structural carbohydrate responses to the combined effects of defoliation and progressive drought in a dioecious tree. New For. 2021, 52, 605–619. [Google Scholar] [CrossRef]
- Sevanto, S.; Mcdowell, N.G.; Dickman, L.T.; Pangle, R.T.; Pockman, W. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Gao, J.; Zhang, Y.; Liu, Y.; Tang, M. Effects of ectomycorrhizal fungi (Suillus variegatus) on the growth, hydraulic function, and non-structural carbohydrates of Pinus tabulaeformis under drought stress. BMC Plant Biol. 2021, 21, 171. [Google Scholar] [CrossRef]
- Rosas, T.; Galiano, L.; Ogaya, R.; Peñuelas, J.; Martínez-Vilalta, J. Dynamics of non-structural carbohydrates in three Mediterranean woody species following long-term experimental drought. Front. Plant Sci. 2013, 4, 400. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.J.; Liu, W.Q.; Huang, F.; Hua, L.; Yu, F.; He, Q.; Qiu, Q. Spatial Patterns of Non-Structural Carbohydrates in Eucalyptus urophylla× E.grandis under Dry-Season Irrigation with Fertilization. Forests 2021, 12, 1049. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Burslem, D.F.; Caduff, A.; Tay, J.; Hector, A. Contrasting nonstructural carbohydrate dynamics of tropical tree seedlings under water deficit and variability. New Phytol. 2015, 205, 1083–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urrutia-Jalabert, R.; Lara, A.; Barichivich, J.; Vergara, N.; Rodriguez, C.G.; Piper, F.I. Low growth sensitivity and fast replenishment of non-structural carbohydrates in a long-lived endangered conifer after drought. Front. Plant Sci. 2020, 11, 905. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.P.; Zhang, W.D.; Li, R.S.; Xu, M.; Wang, S.L. Different responses of non-structural carbohydrates in above-ground tissues/organs and root to extreme drought and re-watering in Chinese fir (Cunninghamia lanceolata) saplings. Trees 2016, 30, 1863–1871. [Google Scholar] [CrossRef]
- Dong, T.; Duan, B.; Zhang, S.; Helena, K.; Lo, N.; Li, C. Growth, biomass allocation and photosynthetic responses are related to intensity of root severance and soil moisture conditions in the plantation tree Cunninghamia lanceolata. Tree Physiol. 2016, 7, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.H.; Wang, Y.X.; Li, Q.; Xiao, W.F.; Song, X.Z. Photosynthesis, ecological stoichiometry, and non-structural carbohydrate response to simulated nitrogen deposition and phosphorus addition in Chinese fir forests. Forests 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Li, S.B.; Zhou, L.L.; Addo-Danso, S.D.; Ding, G.C.; Sun, M.; Wu, S.P.; Lin, S.Z. Nitrogen supply enhances the physiological resistance of Chinese fir plantlets under polyethylene glycol (PEG)-induced drought stress. Sci. Rep. 2020, 10, 7509. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Zou, Z.G.; Zhou, L.L.; Zhang, M.X.; Lin, S.Z.; Li, S.B. Xylem hydraulic functional characteristics in different diameter roots and their correlations with anatomical structures of Chinese fir saplings. Chin. J. Appl. Environ. Biol. 2023, 29, 1–10. (In Chinese) [Google Scholar]
- Li, S.B.; Yang, L.; Huang, X.Y.; Zou, Z.G.; Zhang, M.X.; Guo, W.J.; Zhou, L.L. Mineral Nutrient Uptake, Accumulation, and Distribution in Cunninghamia lanceolata in Response to Drought Stress. Plants 2023, 12, 2140. [Google Scholar] [CrossRef] [PubMed]
- Li, S.B.; Zheng, R.P.; Zhou, L.L.; Zheng, D.; Huang, X.Y.; Wu, Y.L. The hydraulic architecture characteristics of different drought-tolerant Chinese fir plantlets. Chin. J. Appl. Environ. Biol. 2022, 28, 1571–1577. (In Chinese) [Google Scholar]
- Kono, Y.; Ishida, A.; Saiki, S.T.; Yoshimura, K.; Dannoura, M.; Yazaki, K.; Kimura, F.; Yoshimura, J.; Aikawa, S. Initial hydraulic failure followed by late-stage carbon starvation leads to drought-induced death in the tree Trema orientalis. Commun. Biol. 2019, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Lu, R.Y.; Xia, J.Y. Impacts of global environmental change drivers on non-structural carbohydrates in terrestrial plants. Funct. Ecol. 2020, 34, 1525–1536. [Google Scholar] [CrossRef]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef]
- Li, N.N.; He, N.P.; Yu, G.R.; Wang, Q.F.; Sun, J. Leaf non-structural carbohydrates regulated by plant functional groups and climate: Evidences from a tropical to cold-temperate forest transect. Ecol. Indic. 2016, 62, 22–31. [Google Scholar] [CrossRef]
- VonArx, G.; Arzac, A.; Fonti, P.; Frank, D.; Zweifel, R.; Rigling, A.; Galiano, L.; Gessler, A.; Olano, J.M. Responses of sapwood ray parenchyma and non-structural carbohydrates of Pinus sylvestris to drought and long-term irrigation. Funct. Ecol. 2017, 31, 1371–1382. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.K.; Lin, J.; Liu, Q.L.; Ai, Y.F.; Ke, Y.Q.; Chen, C.; He, H. Time-course of photosynthesis and non-structural carbon compounds in the leaves of tea plants (Camellia sinensis L.) in response to deficit irrigation. Agr. Water Manag. 2014, 144, 98–106. [Google Scholar] [CrossRef]
- Savi, T.; García González, A.; Herrera, J.C.; Forneck, A. Gas exchange, biomass and non-structural carbohydrates dynamics in vines under combined drought and biotic stress. BMC Plant Biol. 2019, 19, 408. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhao, P.; Ouyang, L.; Zhu, L.W.; Ni, G.Y.; Schäfer, K.V. Whole-plant water hydraulic integrity to predict drought-induced Eucalyptus urophylla mortality under drought stress. For. Ecol. Manag. 2020, 468, 118179. [Google Scholar] [CrossRef]
- Zheng, X.J.; Xu, G.Q.; Li, Y.; Wu, X. Deepening rooting depths improve plant water and carbon status of a xeric tree during summer drought. Forests 2019, 10, 592. [Google Scholar] [CrossRef] [Green Version]
- Galiano, L.; Timofeeva, G.; Saurer, M.; Siegwolf, R.; Martínez-Vilalta, J.; Hommel, R.; Gessler, A. The fate of recently fixed carbon after drought release: Towards unravelling C storage regulation in Tilia platyphyllos and Pinus sylvestris. Plant Cell Environ. 2017, 40, 1711–1724. [Google Scholar] [CrossRef]
- Trumbore, S.; Czimczik, C.I.; Sierra, C.A.; Muhr, J.; Xu, X. Non-structural carbon dynamics and allocation relate to growth rate and leaf habit in California oaks. Tree Physiol. 2015, 35, 1206–1222. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Martin, R.E. Spectroscopic remote sensing of non-structural carbohydrates in forest canopies. Remote Sens. 2015, 7, 3526–3547. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Germino, M.J.; Breshears, D.D.; Barron-Gafford, G.A.; Guardiola-Claramonte, M.; Zou, C.B.; Huxman, T.E. Nonstructural leaf carbohydrate dynamics of Pinus edulis during drought-induced tree mortality reveal role for carbon metabolism in mortality mechanism. New Phytol. 2013, 197, 1142–1151. [Google Scholar] [CrossRef]
- Luo, G.Y.; Li, J.M.; Guo, S.L.; Li, Y.L.; Jin, Z.X. Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities. Life 2022, 12, 1310. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling. Plant Cell Environ. 2011, 34, 514–524. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Salleo, S. Refilling embolized xylem conduits: Is it a matter of phloem unloading? Plant Sci. 2011, 180, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Li, W.B.; Hartmann, H.; Adams, H.D.; Zhang, H.X.; Jin, C.J.; Zhao, C.Y.; Guan, D.X.; Wang, A.Z.; Yuan, F.H.; Wu, J.B. The sweet side of global change–dynamic responses of non-structural carbohydrates to drought, elevated CO2 and nitrogen fertilization in tree species. Tree Physiol. 2018, 38, 1706–1723. [Google Scholar] [CrossRef] [PubMed]
- Camisón, Á.; Ángela Martín, M.; Dorado, F.J.; Moreno, G.; Solla, A. Changes in carbohydrates induced by drought and waterlogging in Castanea sativa. Trees 2020, 34, 579–591. [Google Scholar] [CrossRef]
- Maguire, A.J.; Kobe, R.K. Drought and shade deplete nonstructural carbohydrate reserves in seedlings of five temperate tree species. Ecol. Evol. 2015, 5, 5711–5721. [Google Scholar] [CrossRef]
- Chen, J.; Dong, T.; Duan, B.; Korpelainen, H.; Niinemets, Ü.; Li, C. Sexual competition and N supply interactively affect the dimorphism and competiveness of opposite sexes in Populus cathayana. Plant Cell Environ. 2015, 38, 1285–1298. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Category | Factor | Fructose | Glucose | Sucrose | Soluble Sugar | Starch | NSCs |
|---|---|---|---|---|---|---|---|
| Aboveground | T | 170.361 ** | 52.083 ** | 157.013 ** | 105.696 ** | 515.943 ** | 48.310 ** |
| D | 33.455 ** | 26.781 ** | 74.731 ** | 5.358 ** | 123.806 ** | 4.048 * | |
| O | 39.500 ** | 387.895 ** | 700.743 ** | 28.792 ** | 375.512 ** | 64.856 ** | |
| T × D | 88.139 ** | 86.482 ** | 59.958 ** | 2.495 | 48.741 ** | 1.554 * | |
| T × O | 21.665 ** | 20.559 ** | 268.612 ** | 9.451 ** | 76.026 ** | 13.638 ** | |
| D ×O | 18.016 ** | 49.337 ** | 88.814 ** | 2.109 * | 12.880 ** | 2.027 * | |
| T × D × O | 20.793 ** | 37.245 ** | 139.497 ** | 1.020 | 48.076 ** | 2.226 ** | |
| Belowground | T | 116.350 ** | 915.967 ** | 425.071 ** | 18.620 ** | 425.071 ** | 2639.884 ** |
| D | 19.229 ** | 952.120 ** | 159.381 ** | 13.711 ** | 159.381 ** | 9.836 ** | |
| O | 2.987 | 206.071 ** | 166.591 ** | 10.933 ** | 166.591 ** | 3.584 ** | |
| T × D | 36.161 ** | 427.521 ** | 57.815 ** | 6.273 ** | 57.815 ** | 23.236 * | |
| T × O | 46.699 ** | 328.833 ** | 22.360 ** | 5.199 ** | 22.360 ** | 3.373 ** | |
| D × O | 51.705 ** | 113.767 ** | 54.996 ** | 5.458 ** | 54.996 ** | 6.403 ** | |
| T × D × O | 36.639 ** | 386.522 ** | 20.080 ** | 3.146 ** | 20.080 ** | 10.114 ** |
| Organs | Index | Stem Xylem | Stem Phloem | Branch Xylem | Branch Phloem | Needles | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fru | Glu | Suc | Fru | Glu | Suc | Fru | Glu | Suc | Fru | Glu | Suc | Fru | Glu | Suc | ||
| Fine root | Fru | 0.520 ** | −0.294 * | −0.243 | 0.060 | −0.363 * | 0.440 ** | 0.814 ** | −0.370 * | −0.351 ** | 0.638 ** | −0.419 ** | 0.695 ** | 0.621 ** | −0.054 | −0.015 |
| Glu | −0.321 * | 0.890 ** | 0.436 ** | 0.384 ** | 0.421 ** | 0.086 | 0.027 | 0.737 ** | 0.289 | −0.282 | 0.688 ** | 0.485 ** | −0.136 | −0.112 | −0.041 | |
| Suc | 0.487 ** | −0.418 ** | −0.333 ** | 0.355 * | −0.608 ** | 0.402 ** | 0.205 | −0.447 ** | −0.391 ** | 0.385 ** | −0.676 ** | 0.187 | 0.153 | 0.339 * | 0.162 | |
| Medium root | Fru | 0.167 | −0.076 | 0.106 | 0.566 ** | −0.060 | −0.179 | 0.179 | −0.247 | −0.386 ** | 0.492 ** | 0.006 | 0.252 | 0.795 ** | −0.577 ** | 0.626 ** |
| Glu | −0.509 ** | 0.028 | −0.242 | 0.213 | 0.437 ** | −0.058 | −0.069 | −0.023 | −0.222 | 0.215 | 0.008 | 0.024 | 0.317 * | 0.106 | 0.472 ** | |
| Suc | −0.036 | 0.498 ** | 0.182 | 0.451 ** | −0.223 | 0.007 | 0.388 ** | 0.422 ** | 0.360 * | −0.074 | −0.095 | 0.486 ** | −0.162 | 0.051 | −0.309 ** | |
| Coarse root | Fru | 0.225 | −0.216 | −0.070 | 0.270 | −0.441 ** | 0.158 | −0.142 | −0.202 | −0.551 ** | 0.098 | 0.101 | −0.055 | 0.201 | −0.123 | 0.474 |
| Glu | 0.783 ** | 0.230 | −0.133 | 0.211 | 0.254 | −0.414 ** | −0.098 | 0.315 * | 0.226 | −0.267 | 0.247 | 0.141 | −0.194 | 0.042 | 0.094 | |
| Suc | 0.356 * | 0.074 | 0.264 | 0.279 | 0.067 | −0.045 | 0.292 * | −0.025 | −0.256 | 0.466 ** | 0.225 | 0.322 ** | 0.745 ** | −0.626 ** | 0.424 ** | |
| Organs | Index | Stem Xylem | Stem Phloem | Branch Xylem | Branch Phloem | Needles | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SS | ST | NSCs | SS | ST | NSCs | SS | ST | NSCs | SS | ST | NSCs | SS | ST | NSCs | ||
| Fine root | SS | 0.126 | −0.158 | 0.067 | 0.092 | −0.334 * | −0.036 | 0.150 | −0.271 | 0.048 | 0.282 | −0.500 ** | 0.093 | 0.409 ** | −0.569 ** | 0.196 |
| ST | −0.428 ** | −0.230 | −0.527 ** | −0.054 | 0.332 * | 0.074 | −0.357 * | −0.087 | −0.406 ** | −0.383 ** | 0.432 ** | −0.222 | 0.246 | 0.124 | −0.202 | |
| NSCs | −0.016 | −0.250 | −0.114 | 0.072 | −0.192 | −0.002 | −0.026 | −0.252 | −0.125 | 0.163 | −0.359 * | 0.026 | 0.373 ** | −0.657 ** | 0.124 | |
| Medium root | SS | 0.619 ** | −0.294 * | 0.422 ** | 0.357 * | −0.237 | 0.199 | 0.181 | −0.007 | 0.175 | −0.580 ** | −0.601 ** | 0.183 | 0.400 ** | −0.781 ** | −0.114 |
| ST | −0.481 ** | 0.430 ** | −0.196 | −0.622 ** | 0.390 ** | −0.362 * | −0.558 ** | 0.458 ** | −0.221 | 0.054 | 0.303 * | 0.252 | −0.576 ** | 0.489 ** | −0.252 | |
| NSCs | 0.491 ** | −0.162 | 0.981 ** | 0.072 | −0.059 | 0.032 | −0.043 | 0.228 | 0.107 | 0.575 ** | −0.493 ** | 0.248 | 0.151 | −0.700 ** | −0.309 * | |
| Coarse root | SS | 0.256 | −0.260 | 0.173 | 0.147 | −0.420 ** | −0.013 | 0.235 | −0.268 | 0.147 | 0.497 ** | −0.634 ** | 0.284 | 0.574 ** | −0.797 ** | 0.301 * |
| ST | −0.340 * | 0.332 * | −0.236 | −0.413 ** | 0.281 | −0.338 * | −0.336 * | 0.259 | −0.257 | −0.529 ** | 0.389 ** | −0.420 ** | −0.560 ** | 0.371 ** | −0.461 ** | |
| NSCs | 0.153 | −0.161 | 0.102 | −0.049 | −0.310 * | −0.183 | 0.061 | −0.078 | 0.035 | 0.329 * | −0.500 ** | 0.154 | 0.387 ** | −0.796 ** | 0.095 | |
| Organ | Index | Stem Phloem | Branch Phloem | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fru | Glu | Suc | SS | ST | NSCs | Fru | Glu | Suc | SS | ST | NSCs | ||
| Stem xylem | Fru | 0.015 | −0.412 ** | 0.226 | 0.129 | 0.464 ** | 0.370 ** | 0.555 ** | −0.191 | 0.085 | −0.224 | 0.052 | −0.201 |
| Glu | 0.176 | 0.486 ** | −0.144 | −0.096 | −0.125 | −0.211 | −0.398 ** | 0.696 ** | 0.257 | −0.092 | −0.467 ** | −0.222 | |
| Suc | 0.128 | 0.243 | −0.175 | −0.051 | −0.141 | −0.124 | −0.279 | 0.600 ** | −0.032 | −0.017 | −0.134 | −0.054 | |
| SS | −0.390 ** | 0.138 | −0.323 * | 0.351 * | −0.593 ** | 0.080 | −0.231 | −0.192 | −0.335 ** | 0.583 ** | 0.068 | 0.581 ** | |
| ST | 0.449 ** | 0.373 | −0.052 | −0.265 | 0.241 | −0.164 | 0.008 | 0.479 ** | 0.189 | −0.427 ** | −0.047 | −0.425 ** | |
| NSCs | −0.258 | 0.174 | −0.363 ** | 0.281 | −0.548 ** | 0.027 | −0.244 | −0.036 | −0.291 * | 0.472 ** | 0.057 | 0.471 ** | |
| Branch xylem | Fru | −0.064 | −0.280 | 0.209 | 0.075 | 0.303 * | 0.232 | 0.492 ** | −0.176 | 0.710 ** | −0.290 * | 0.221 | −0.216 |
| Glu | 0.040 | 0.444 ** | −0.247 | −0.008 | −0.352 * | −0.185 | −0.452 ** | 0.631 ** | 0.127 | 0.114 | −0.459 ** | −0.022 | |
| Suc | −0.220 | 0.356 * | −0.533 ** | 0.257 | −0.761 ** | −0.107 | −0.448 ** | 0.203 | −0.218 | 0.542 ** | −0.100 | 0.494 ** | |
| SS | −0.478 ** | 0.371 ** | −0.011 | 0.279 | −0.428 ** | 0.085 | −0.099 | 0.188 | −0.019 | 0.366 ** | −0.149 | 0.310 * | |
| ST | 0.324 * | 0.160 | 0.023 | −0.139 | 0.240 | −0.028 | 0.104 | 0.454 ** | 0.404 ** | −0.436 ** | −0.267 | −0.496 | |
| NSCs | −0.207 | 0.370 ** | 0.110 | 0.151 | −0.190 | 0.066 | 0.006 | 0.353 * | 0.179 | 0.037 | −0.246 | −0.034 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Guo, W.; Yang, L.; Zou, Z.; Zhang, X.; Addo-Danso, S.D.; Zhou, L.; Li, S. Effects of Drought Stress on Non-Structural Carbohydrates in Different Organs of Cunninghamia lanceolata. Plants 2023, 12, 2477. https://doi.org/10.3390/plants12132477
Huang X, Guo W, Yang L, Zou Z, Zhang X, Addo-Danso SD, Zhou L, Li S. Effects of Drought Stress on Non-Structural Carbohydrates in Different Organs of Cunninghamia lanceolata. Plants. 2023; 12(13):2477. https://doi.org/10.3390/plants12132477
Chicago/Turabian StyleHuang, Xiaoyan, Wenjuan Guo, Li Yang, Zhiguang Zou, Xinyang Zhang, Shalom Daniel Addo-Danso, Lili Zhou, and Shubin Li. 2023. "Effects of Drought Stress on Non-Structural Carbohydrates in Different Organs of Cunninghamia lanceolata" Plants 12, no. 13: 2477. https://doi.org/10.3390/plants12132477