

Improving Boron and Molybdenum Use Efficiencies in Contrasting Cultivars of Subirrigated Greenhouse-Grown Pot Chrysanthemums

Abstract
:
1. Introduction
2. Results
2.1. Growth of Two Chrysanthemum Cultivars Supplied with Moderate to Deficient Boron Levels
2.1.1. Summary of Significant Effects
2.1.2. Morphological Characteristics
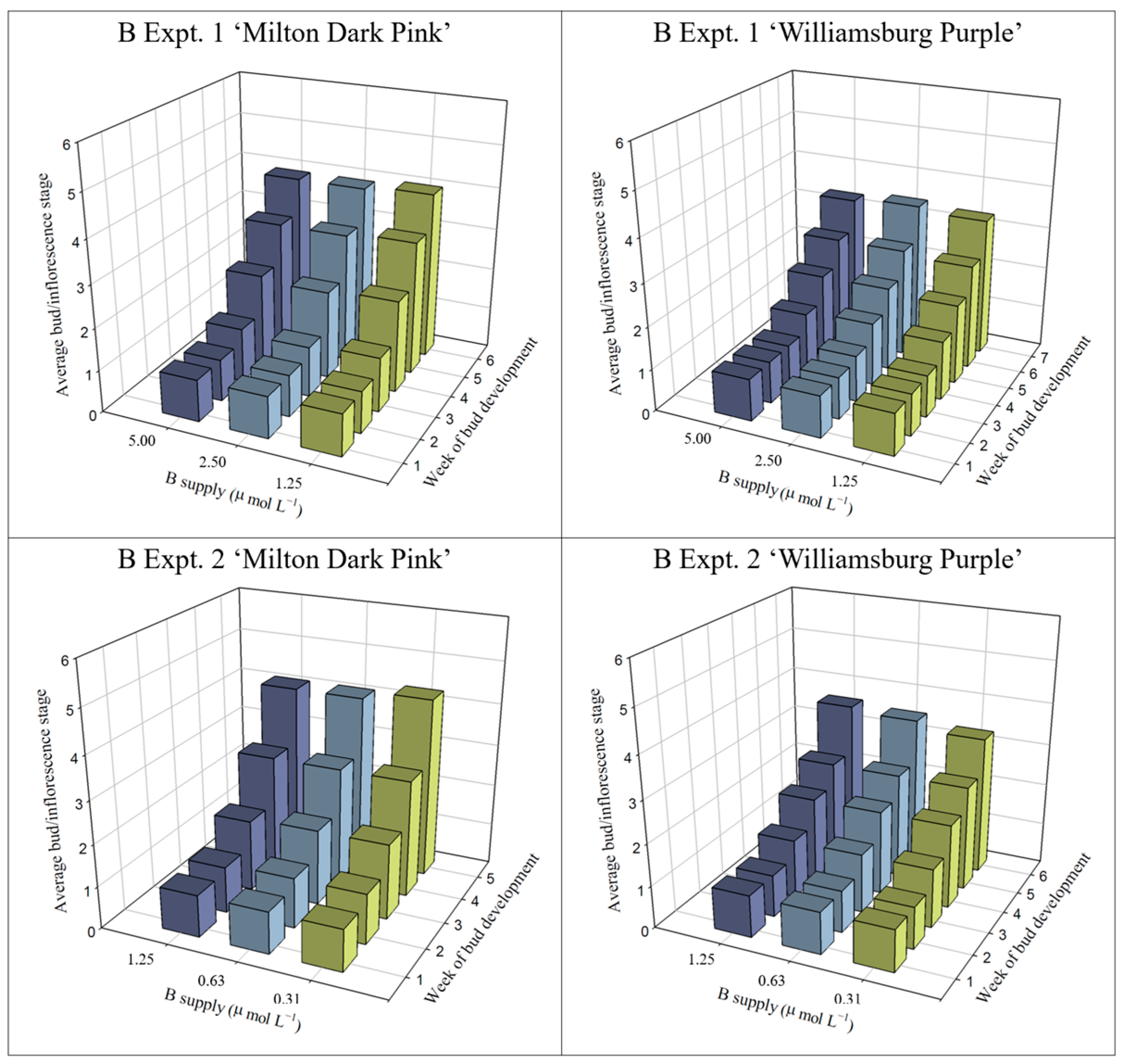
2.1.3. Bud and Inflorescence Development
2.1.4. Leaf Nutrient Composition at Bud Emergence
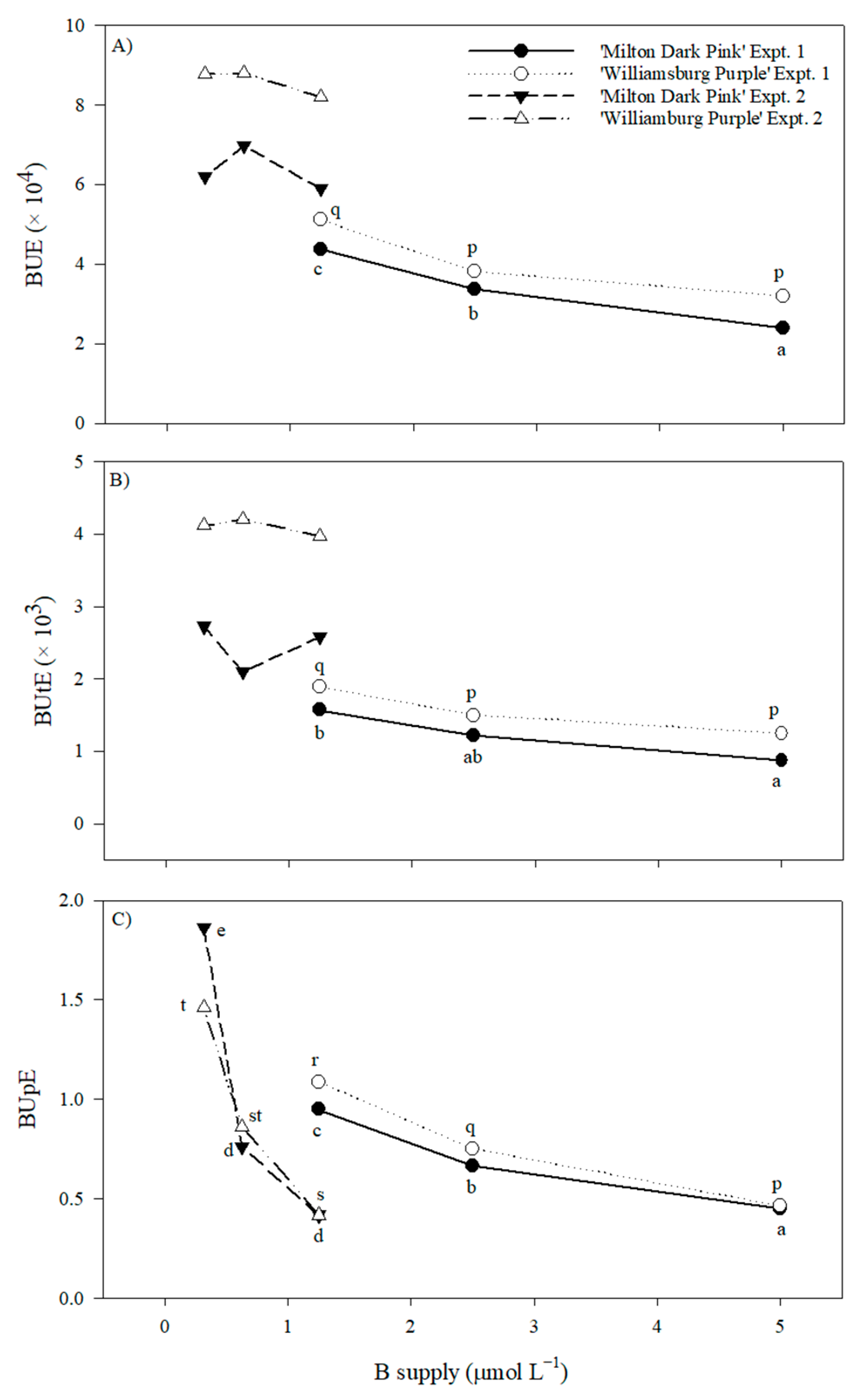
2.1.5. Nutrient Use Efficiency
2.2. Growth of Three Chrysanthemum Cultivars Supplied with Moderate Molybdenum Levels
2.2.1. Summary of Significant Effects
2.2.2. Morphological Characteristics
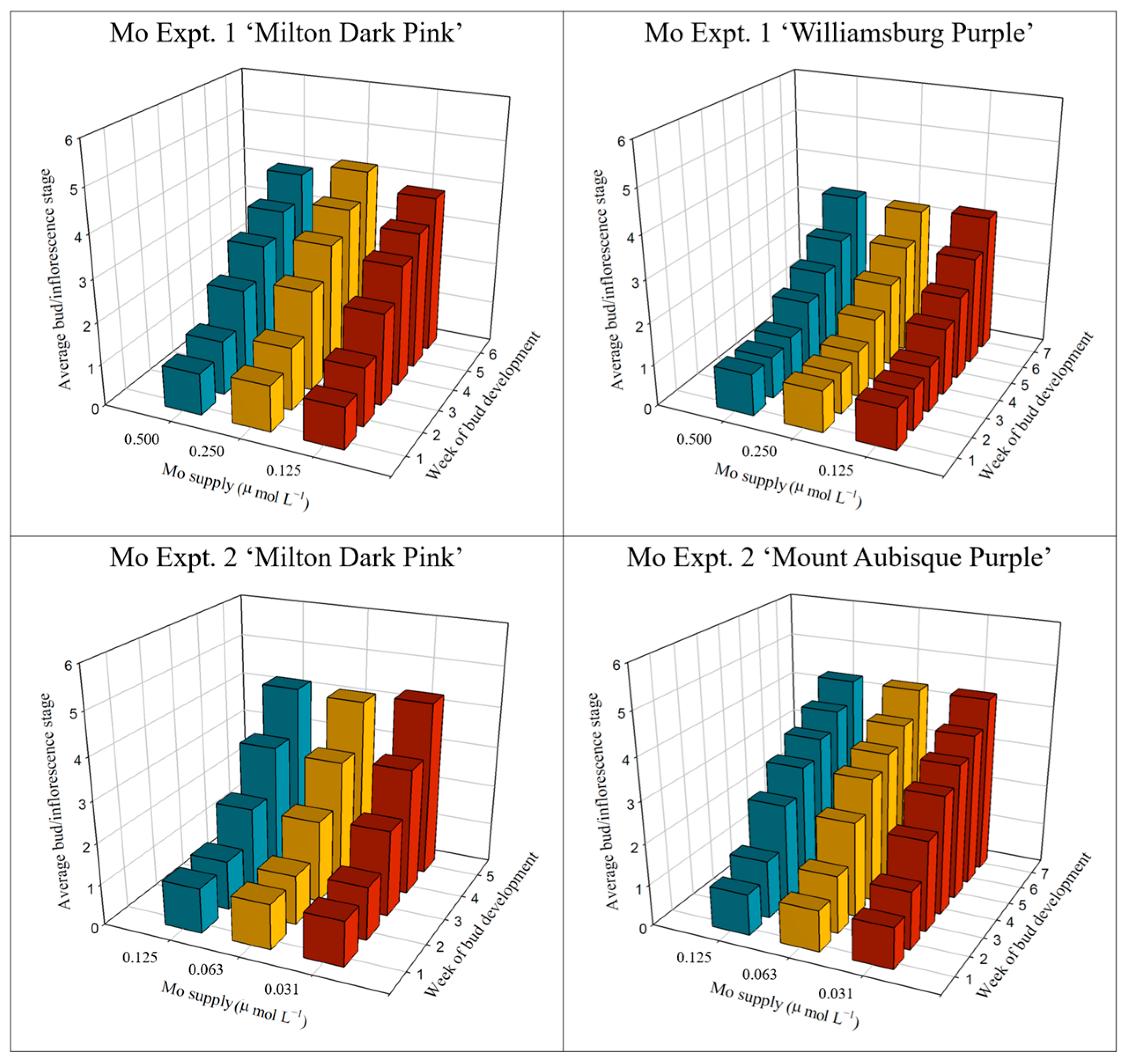
2.2.3. Bud and Inflorescence Development
2.2.4. Leaf Nutrient Composition
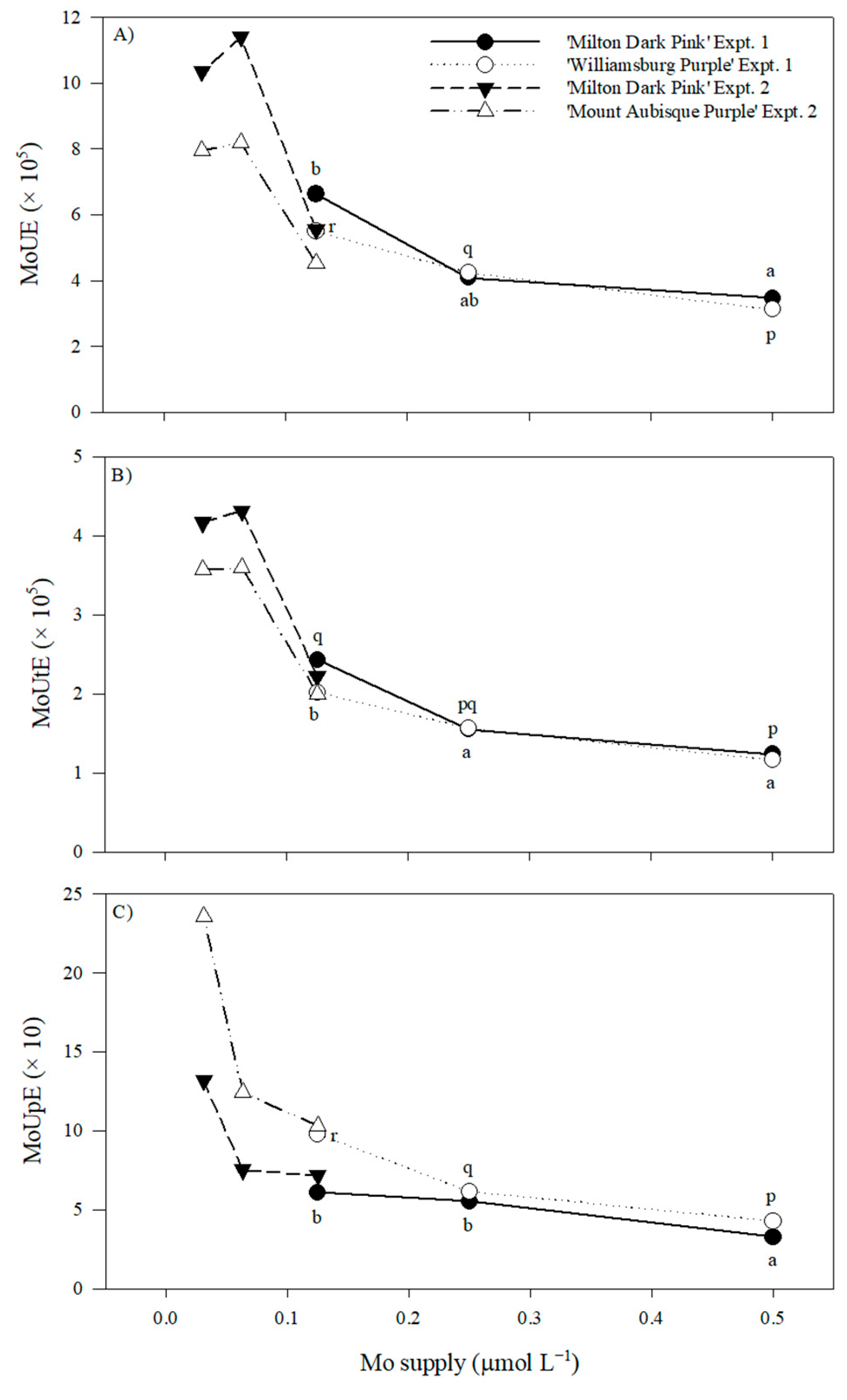
2.2.5. Nutrient Use Efficiency
3. Discussion
3.1. Optimization of Boron and Molybdenum Fertilization
3.2. Mechanisms for Improved Boron and Molybdenum Use Efficiencies
3.3. Potential Interactions of Boron or Molybdenum with Other Elements
3.4. Prospects for the Floricultural Industry
4. Materials and Methods
4.1. Plant Growth Conditions
4.2. Integrated Pest Management
4.3. Data Collection
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- White, P.J. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 4–47. [Google Scholar] [CrossRef]
- White, P.J. Long-distance transport in the xylem and phloem. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Boston, MA, USA, 2012; pp. 49–70. [Google Scholar] [CrossRef]
- Sutton, W.J.; Bozzo, G.G.; Carlow, C.; Macdonald, W.N.; Shelp, B.J. Strategic timing and rate of sulphur fertilization improves sulphur use efficiency in subirrigated greenhouse-grown chrysanthemums. Can. J. Plant Sci. 2019, 99, 654–665. [Google Scholar] [CrossRef]
- Shelp, B.J.; Sutton, W.J.; Flaherty, E.J. Strategic timing and rate of phosphorus fertilization improves phosphorus-use efficiency in two contrasting cultivars of subirrigated greenhouse-grown chrysanthemum. Can. J. Plant Sci. 2020, 100, 264–275. [Google Scholar] [CrossRef]
- MacDonald, W.N.; Blom, T.J.; Tsujita, M.J.; Shelp, B.J. Review: Improving nitrogen use efficiency of potted chrysanthemum: Strategies and benefits. Can. J. Plant Sci. 2013, 93, 1009–1016. [Google Scholar] [CrossRef]
- Ferrarezi, R.; Weaver, G.M.; Van Iersel, M.W.; Testezlaf, R. Subirrigation: Historical overview, challenges, and future prospects. Horttechnology 2015, 25, 262–276. [Google Scholar] [CrossRef] [Green Version]
- Semananda, N.P.K.; Ward, J.D.; Myers, B.R. A semi-systematic review of capillary irrigation: The benefits, limitations, and opportunities. Horticulturae 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Syngenta Flowers. Pot Mum Culture Guide. 2015. Available online: https://www.syngentaflowers-us.com/sites/g/files/kgtney846/files/file_field/import/field_media_link/sites/g/files/zhg721/f/potmumculturalinformation_1.pdf (accessed on 3 November 2022).
- ICL. Peter’s Professional 17-3-17 Peat-Lite Neutral Cal-Mag. 2022. Available online: https://icl-sf.com/uploads/USA/Product Sheets/OH/g99230_peters_pro_17-3-17_plncm.pdf (accessed on 18 November 2022).
- Master Plant-Prod Inc. Plant-Prod Solutions 17-5-17 Fusion Water Soluble Fertilizer. 2019. Available online: https://www.plantprod.com/product/plant-prod-solutions-17-5-17-fusion/ (accessed on 8 November 2022).
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Sonneveld, C.; Kreij, C. Nutrient solutions for vegetables and flowers grown in water or substrates. In Series Voedingsoplossingen Glastuinbouw, No 8, 6th ed.; Glasshouse Crops Research Station: Naaldwijk, The Netherlands, 1987; pp. 1–45. [Google Scholar]
- MacDonald, W.N.; Tsujita, M.J.; Blom, T.J.; Shelp, B.J. Impact of various combinations of nitrate and chloride on nitrogen remobilization in potted chrysanthemum grown in a subirrigation system. Can. J. Plant Sci. 2014, 94, 643–675. [Google Scholar] [CrossRef]
- Shelp, B.J.; Solntseva, I.; Sutton, W.J.; Lum, G.B.; Kessel, C.W. Optimizing supply and timing of nitrogen application for subirrigated potted chrysanthemums. Can. J. Plant Sci. 2017, 97, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Shelp, B.J.; Flaherty, E.J.; Duncan Stephens, S.; Donetz, A.J. Improving zinc and copper delivery for subirrigated greenhouse-grown pot chrysanthemums. Can. J. Plant Sci. 2021, 101, 268–273. [Google Scholar] [CrossRef]
- Duncan Stephens, S.; Flaherty, E.J.; Sutton, W.J.; Macdonald, W.N.; Shelp, B.J. Further optimization of macronutrient delivery for subirrigated greenhouse-grown chrysanthemums: Calcium and magnesium. Can. J. Plant Sci. 2021, 101, 129–134. [Google Scholar] [CrossRef]
- Donetz, A.J.; Flaherty, E.J.; Teeter-Wood, K.; Hoover, G.J.; Dayboll, C.; Shelp, B.J. Optimizing manganese and iron delivery for contrasting cultivars of subirrigated greenhouse-grown pot chrysanthemums. Can. J. Plant Sci. 2022, 102, 823–834. [Google Scholar] [CrossRef]
- Shelp, B.J. Physiology and biochemistry of boron in plants. In Boron and Its Role in Crop Production; Gupta, U.C., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 53–85. [Google Scholar]
- Blevins, D.G.; Lukaszewski, K.M. Boron in plant structure and function. Annu. Rev. Plant Biol. 1998, 49, 481–500. [Google Scholar] [CrossRef] [Green Version]
- Wimmer, M.A.; Eichert, T. Review: Mechanisms for boron deficiency-mediated changes in plant water relations. Plant Sci. 2013, 203–204, 25–32. [Google Scholar] [CrossRef]
- Tanaka, M.; Fujiwara, T. Physiological roles and transport mechanisms of boron: Perspectives from plants. Eur. J. Physiol. 2008, 456, 671–677. [Google Scholar] [CrossRef]
- U.S. Borax. Boron Deficiency in Chrysanthemums. 2023. Available online: https://agriculture.borax.com/crop-guides/flowers-and-ornamentals/chrysanthemum (accessed on 25 February 2021).
- Brown, P.H.; Shelp, B.J. Boron mobility in plants. Plant Soil 1997, 193, 85–101. [Google Scholar] [CrossRef]
- Marentes, E.; Shelp, B.J.; Vanderpool, R.A.; Spiers, G.A. Retranslocation of boron in broccoli and lupin during early reproductive growth. Physiol. Plant. 1997, 100, 389–399. [Google Scholar] [CrossRef]
- Shelp, B.J.; Kitheka, A.M.; Vanderpool, R.A.; Van Cauwenberghe, O.R.; Spiers, G.A. Xylem-to-phloem transfer of boron in broccoli and lupin during early reproductive growth. Physiol. Plant. 1998, 104, 533–540. [Google Scholar] [CrossRef]
- Huang, L.; Bell, R.W.; Dell, B. Boron supply into wheat (Triticum aestivum L. cv. Wilgoyne) ears whilst still enclosed within leaf sheaths. J. Exp. Bot. 2001, 52, 1731–1738. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Bell, R.W.; Dell, B. Evidence of phloem boron transport in response to interrupted boron supply in white lupin (Lupinus albus L. cv. Kiev Mutant) at the reproductive stage. J. Exp. Bot. 2008, 59, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Stangoulis, J.C.R.; Brown, P.H.; Bellaloui, N.; Reid, R.J.; Graham, R.D. The efficiency of boron utilisation in canola. Funct. Plant Biol. 2001, 28, 1109–1114. [Google Scholar] [CrossRef]
- Stangoulis, J.; Tate, M.; Graham, R.; Bucknall, M.; Palmer, L.; Boughton, B.; Reid, R. The mechanism of boron mobility in wheat and canola phloem. Plant Physiol. 2010, 153, 876–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, J.; Yamagami, M.; Noguchi, K.; Hayashi, H.; Fujiwara, T. Preferential translocation of boron to young leaves in Arabidopsis thaliana Regulated by the BOR1 Gene. Soil Sci. Plant Nutr. 2001, 47, 345–357. [Google Scholar] [CrossRef]
- Matoh, T.; Ochiai, K. Distribution and partitioning of newly taken-up boron in sunflower. Plant Soil 2005, 278, 351–360. [Google Scholar] [CrossRef]
- Hajiboland, R.; Bahrami-Rad, S.; Bastani, S.; Tolrà, R.; Poschenrieder, C. Boron re-translocation in tea (Camellia sinensis (L.) O. Kuntze) plants. Acta Physiol. Plant. 2013, 35, 2373–2381. [Google Scholar] [CrossRef]
- Du, W.; Pan, Z.Y.; Hussain, S.B.; Han, Z.X.; Peng, S.A.; Liu, Y.Z. Foliar supplied boron can be transported to roots as a boron-sucrose complex via phloem in citrus trees. Front. Plant Sci. 2020, 11, 250. [Google Scholar] [CrossRef]
- Shelp, B.J.; Vivekanandan, P.; Vanderpool, R.A.; Kitheka, A.M. Translocation and effectiveness of foliar-fertilized boron in broccoli plants of varying boron status. Plant Soil 1996, 183, 309–313. [Google Scholar] [CrossRef]
- Wu, X.; Riaz, M.; Yan, L.; Jiang, C. Distribution and mobility of foliar-applied boron (10B) in citrange rootstock under different boron conditions. J. Plant Growth Regul. 2020, 39, 575–582. [Google Scholar] [CrossRef]
- Bittner, F. Molybdenum metabolism in plants and crosstalk to iron. Front. Plant Sci. 2014, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Gupta, U.C.; Lipsett, J. Molybdenum in soils, plants, and animals. Adv. Agron. 1981, 34, 73–115. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S.D. The role of molybdenum in agricultural plant production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Chamizo-Ampudia, A.; Galván, A.; Fernández, E. Molybdenum metabolism in plants. Metallomics 2013, 5, 1191–1203. [Google Scholar] [CrossRef]
- Kannan, S.; Ramani, S. Studies on molybdenum absorption and transport in bean and rice. Plant Physiol. 1978, 62, 179–181. [Google Scholar] [CrossRef]
- Mauk, C.S.; Noodén, L.D. Regulation of mineral redistribution in pod-bearing soybean explants. J. Exp. Bot. 1992, 43, 1429–1440. [Google Scholar] [CrossRef]
- Ontario Ministry of Agriculture Food and Rural Affairs. Guide to Greenhouse Floriculture Production: Publication 370. Available online: https://files.ontario.ca/omafra-guide-to-greenhouse-floriculture-370-en-2022-11-04.pdf (accessed on 2 November 2022).
- Hill Laboratories. Crop Guide-Chrysanthemum. 2019. Available online: https://www.hill-laboratories.com/assets/Documents/Crop-Guides/3451v3-Crop-Guide-Chrysanthemum.pdf (accessed on 2 November 2022).
- Plank, O.C.; Granberry, D.M.; Phatak, S. Plant Analysis Handbook for Georgia. 2022. Available online: http://aesl.ces.uga.edu/publications/plant/contable.html#floral (accessed on 2 November 2022).
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Rooda van Eysinga, J.P.N.L.; Smilde, K.W. Nutritional Disorders in Chrysanthemums; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1980; p. 42. [Google Scholar]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.M.; Yvin, J.C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 317. [Google Scholar] [CrossRef] [Green Version]
- Government of Ontario. Nutrient Management Act, 2002, S.O. 2002, c. 4. Available online: https://www.ontario.ca/laws/statute/02n04 (accessed on 22 February 2023).
- Ontario Ministry of the Environment. Greenhouse Wastewater Monitoring Project (2010 and 2011). Available online: https://www.ontario.ca/page/greenhouse-wastewater-monitoring-project-2010-and-2011 (accessed on 22 February 2022).
- Maguire, T.J.; Wellen, C.; Stammler, K.L.; Mundle, S.O.C. Increased nutrient concentrations in Lake Erie tributaries influenced by greenhouse agriculture. Sci. Total Environ. 2018, 633, 433–440. [Google Scholar] [CrossRef]
- Pereira, G.L.; Siqueira, J.A.; Batista-Silva, W.; Cardoso, F.B.; Nunes-Nesi, A.; Araújo, W.L. Boron: More than an essential element for land plants? Front. Plant Sci. 2021, 11, 610307. [Google Scholar] [CrossRef]
- Miwa, K.; Aibara, I.; Fujiwara, T. Arabidopsis thaliana BOR4 is upregulated under high boron conditions and confers tolerance to high boron. Soil Sci. Plant Nutr. 2014, 60, 349–355. [Google Scholar] [CrossRef]
- Wakuta, S.; Mineta, K.; Amano, T.; Toyoda, A.; Fujiwara, T.; Naito, S.; Takano, J. Evolutionary divergence of plant borate exporters and critical amino acid residues for the polar localization and boron-dependent vacuolar sorting of AtBOR1. Plant Cell Physiol. 2015, 56, 852–862. [Google Scholar] [CrossRef] [Green Version]
- Aibara, I.; Hirai, T.; Kasai, K.; Takano, J.; Onouchi, H.; Naito, S.; Fujiwara, T.; Miwa, K. Boron-dependent translational suppression of the borate exporter BOR1 contributes to the avoidance of boron toxicity. Plant Physiol. 2018, 177, 759–774. [Google Scholar] [CrossRef] [Green Version]
- Du, C.W.; Wang, Y.H.; Xu, F.S.; Yang, Y.H.; Wang, H.Y. Study on the physiological mechanism of boron utilization efficiency in rape cultivars. J. Plant Nutr. 2002, 25, 231–244. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Z.; Yang, L.; Wang, Z.; Shi, L.; Naran, R.; Azadi, P.; Xu, F. Differences in cell wall components and allocation of boron to cell walls confer variations in sensitivities of Brassica napus cultivars to boron deficiency. Plant Soil 2012, 354, 383–394. [Google Scholar] [CrossRef]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, I.; Muthukumar, B.; Hyeong, C.P.; Buchner, P.; Lahner, B.; Danku, J.; Zhao, K.; Lee, J.; Hawkesford, M.J.; Guerinot, M.L.; et al. Variation in molybdenum content across broadly distributed populations of Arabidopsis thaliana is controlled by a mitochondrial molybdenum transporter (MOT1). PLoS Genet. 2008, 4, e1000004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.Y.; Liu, H.; Zhu, Y.F.; Pinson, S.R.M.; Lin, H.X.; Guerinot, M.L.; Zhao, F.J.; Salt, D.E. Natural variation in a molybdate transporter controls grain molybdenum concentration in rice. New Phytol. 2019, 221, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Gasber, A.; Klaumann, S.; Trentmann, O.; Trampczynska, A.; Clemens, S.; Schneider, S.; Sauer, N.; Feifer, I.; Bittner, F.; Mendel, R.R.; et al. Identification of an Arabidopsis solute carrier critical for intracellular transport and inter-organ allocation of molybdate. Plant Biol. 2011, 13, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R.; Kruse, T. Cell biology of molybdenum in plants and humans. Biochim. Biophys. Acta 2012, 1823, 1568–1579. [Google Scholar] [CrossRef] [Green Version]
- Hale, K.L.; McGrath, S.P.; Lombi, E.; Stack, S.M.; Terry, N.; Pickering, I.J.; George, G.N.; Pilon-Smits, E.A.H. Molybdenum sequestration in Brassica species. A role for anthocyanins? Plant Physiol. 2001, 126, 1391–1402. [Google Scholar] [CrossRef] [Green Version]
- Steinke, D.R.; Majak, W.; Sorensen, T.S.; Parvez, M. Chelation of molybdenum in Medicago sativa (alfalfa) grown on reclaimed mine tailings. J. Agric. Food Chem. 2008, 56, 5437–5442. [Google Scholar] [CrossRef]
- Tejada-Jiménez, M.; Galván, A.; Fernández, E.; Llamas, Á. Homeostasis of the micronutrients Ni, Mo and Cl with specific biochemical functions. Curr. Opin. Plant Biol. 2009, 12, 358–363. [Google Scholar] [CrossRef]
- Heuwinkel, H.; Kirkby, E.A.; Bot, J.L.; Marschner, H. Phosphorus deficiency enhances molybdenum uptake by tomato plants. J. Plant Nutr. 1992, 15, 549–568. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Hara, T.; Sonada, Y. Effects of boron deficiencies and calcium supply on the calcium metabolism in tomato plant. Plant Soil 1986, 93, 223–230. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Hara, T.; Sonada, Y. Distribution of calcium and boron in the pectin fraction of tomato leaf cell wall. Plant Cell Physiol. 1986, 27, 729–732. [Google Scholar]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, K.L.; Tyerman, S.D.; Kaiser, B.N. Molybdate transport through the plant sulfate transporter SHST1. FEBS Lett. 2008, 582, 1508–1513. [Google Scholar] [CrossRef]
- Government of Ontario. Ontario Water Resources Act, R.S.O. 1990, c. O.40. Available online: https://www.ontario.ca/laws/statute/90o40#Top (accessed on 30 April 2010).
- Shelp, B.J.; Flaherty, E.J.; Sutton, W.J.; Schenck, L.M.; Aalbers, J. Commercial validation of a modified method for delivering low nitrogen, phosphorus and potassium inputs to greenhouse-grown subirrigated pot chrysanthemums. Can. J. Plant Sci. 2021, 101, 962–966. [Google Scholar] [CrossRef]
- Lévesque, V.; Doraisa, M.; Gravel, V.; Ménard, C.; Antoun, H.; Rochette, P.; Roy, S. The use of artificial wetlands to treat greenhouse effluents. Acta Hortic. 2011, 893, 1185–1192. [Google Scholar] [CrossRef]
- Agriculture and Agri-Food Canada. Statistical Overview of the Canadian Ornamental Industry. Available online: https://agriculture.canada.ca/en/sector/horticulture/reports/statistical-overview-canadian-ornamental-industry-2021 (accessed on 24 March 2021).
- Radhamani, R.; Kannan, R.; Rakkiyappan, P. Leaf chlorophyll meter readings as an indicator for sugarcane yield under iron deficient typic Haplustert. Sugar Tech. 2016, 18, 61–66. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | B Supply (μmol L−1) | Shoot | Bud/Inflorescence | |||||
|---|---|---|---|---|---|---|---|---|
| Height (cm plant−1) | DM (g plant−1) | DM (g plant−1) | Bloom Dia. (cm plant−1) | Development (Stage) | Number (Total plant−1) | |||
| Experiment 1 | ||||||||
| ‘Milton Dark Pink’ | 5.00 | 33.79 | 5.44 | 1.99 | 5.86 | 3.9 | 34.1a | |
| 2.50 | 35.22 | 5.59 | 2.00 | 5.95 | 3.9 | 32.4ab | ||
| 1.25 | 34.79 | 5.21 | 1.87 | 5.91 | 4.0 | 31.4b | ||
| ‘Williamsburg Purple’ | 5.00 | 35.26 | 6.47 | 2.92 | 4.56 | 3.3 | 36.1 | |
| 2.50 | 34.38 | 6.39 | 2.83 | 4.58 | 3.4 | 34.8 | ||
| 1.25 | 35.74 | 6.10 | 2.58 | 4.60 | 3.3 | 31.7 | ||
| Experiment 2 | ||||||||
| ‘Milton Dark Pink’ | 1.25 | 23.3 | 2.98b | 1.30b | 6.39 | 4.1 | 25.6 | |
| 0.63 | 23.1 | 3.00b | 1.33b | 6.44 | 4.1 | 25.4 | ||
| 0.31 | 24.4 | 3.35a | 1.47a | 6.42 | 4.3 | 26.3 | ||
| ‘Williamsburg Purple’ | 1.25 | 23.6 | 4.20 | 2.04 | 5.48 | 3.5 | 23.8 | |
| 0.63 | 23.6 | 4.28 | 2.06 | 5.55 | 3.5 | 24.3 | ||
| 0.31 | 22.5 | 3.78 | 1.78 | 5.51 | 3.4 | 22.6 | ||
| Cultivar | B Supply (μmol L−1) | Leaf Concentration (% DM) | |||||
|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | ||
| Experiment 1 | |||||||
| ‘Milton Dark Pink’ | 5.00 | 5.32 | 0.91 | 5.49 | 1.60 | 0.55 | 0.29 |
| 2.50 | 5.33 | 0.91 | 5.30 | 1.64 | 0.56 | 0.29 | |
| 1.25 | 5.26 | 0.86 | 5.35 | 1.50 | 0.52 | 0.30 | |
| ‘Williamsburg Purple’ | 5.00 | 5.55 | 0.76 | 5.54 | 1.26 | 0.42 | 0.30 |
| 2.50 | 5.63 | 0.76 | 5.35 | 1.27 | 0.42 | 0.30 | |
| 1.25 | 5.67 | 0.78 | 5.43 | 1.28 | 0.42 | 0.31 | |
| Experiment 2 | |||||||
| ‘Milton Dark Pink’ | 1.25 | 6.29 | 1.01 | 6.43b | 1.70 | 0.70 | 0.30 |
| 0.63 | 6.44 | 1.02 | 6.60ab | 1.69 | 0.74 | 0.30 | |
| 0.31 | 6.51 | 1.02 | 6.95a | 1.71 | 0.73 | 0.29 | |
| ‘Williamsburg Purple’ | 1.25 | 6.45 | 0.92 | 6.51 | 1.26 | 0.50 | 0.27 |
| 0.63 | 6.52 | 0.90 | 6.50 | 1.26 | 0.49 | 0.28 | |
| 0.31 | 6.60 | 0.92 | 6.58 | 1.28 | 0.51 | 0.28 | |
| Cultivar | B Supply (μmol L−1) | Leaf Concentration (mg kg−1 DM) | |||||
|---|---|---|---|---|---|---|---|
| B | Mo | Cu | Zn | Mn | Fe | ||
| Experiment 1 | |||||||
| ‘Milton Dark Pink’ | 5.00 | 48.8 | 2.3 | 6.4 | 49.5 | 85.0 | 119.8 |
| 2.50 | 49.0 | 2.4 | 7.1 | 53.5 | 81.3 | 116.3 | |
| 1.25 | 46.5 | 2.4 | 5.7 | 42.8 | 74.0 | 94.5 | |
| ‘Williamsburg Purple’ | 5.00 | 42.0 | 4.6 | 5.6 | 35.3 | 66.5 | 91.0 |
| 2.50 | 42.3 | 4.8 | 6.5 | 44.5 | 76.0 | 102.5 | |
| 1.25 | 41.8 | 4.1 | 5.2 | 34.5 | 79.3 | 115.3 | |
| Experiment 2 | |||||||
| ‘Milton Dark Pink’ | 1.25 | 38.5a | 2.2 | 4.6 | 49.3 | 66.5 | 111.0 |
| 0.63 | 19.3b | 1.9 | 4.3 | 46.3 | 68.8 | 98.5 | |
| 0.31 | 11.3c | 2.0 | 4.3 | 43.0 | 71.8 | 109.5 | |
| ‘Williamsburg Purple’ | 1.25 | 33.8a | 4.3 | 3.0 | 30.3 | 101.8 | 102.5 |
| 0.63 | 19.5b | 4.2 | 2.9 | 28.5 | 110.5 | 104.0 | |
| 0.31 | 11.3c | 3.6 | 3.0 | 30.0 | 115.0 | 98.5 | |
| Cultivar | Mo Supply (μmol L−1) | Shoot | Bud/Inflorescence | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Height (cm plant−1) | DM (g plant−1) | SPAD value | DM (g plant−1) | Bloom Dia. (cm plant−1) | Development (Stage) | Number (Total plant−1) | ||||
| Experiment 1 | ||||||||||
| ‘Milton Dark Pink’ | 0.500 | 30.64 | 4.83 | 44.02ab | 1.74a | 5.58 | 3.9a | 35.5 | ||
| 0.250 | 32.36 | 5.69 | 44.74a | 2.17b | 5.72 | 4.2b | 36.8 | |||
| 0.125 | 30.34 | 5.05 | 43.60b | 1.86ab | 5.47 | 3.9a | 36.5 | |||
| ‘Williamsburg Purple’ | 0.500 | 30.77 | 6.61 | 45.92 | 2.46 | 4.53 | 3.3 | 33.4 | ||
| 0.250 | 30.47 | 6.56 | 46.39 | 2.44 | 4.53 | 3.2 | 33.9 | |||
| 0.125 | 32.60 | 6.75 | 47.10 | 2.48 | 4.64 | 3.3 | 32.6 | |||
| Experiment 2 | ||||||||||
| ‘Milton Dark Pink’ | 0.125 | 29.71 | 4.96 | 43.16 | 1.99 | 6.64 | 4.2 | 31.0 | ||
| 0.063 | 30.22 | 4.95 | 43.70 | 1.92 | 6.72 | 4.1 | 29.5 | |||
| 0.031 | 29.61 | 4.92 | 43.13 | 1.99 | 6.82 | 4.3 | 29.7 | |||
| ‘Mount Aubisque Purple’ | 0.125 | 30.67 | 5.80 | 40.52a | 2.56 | 6.45 | 4.3 | 23.9 | ||
| 0.063 | 30.02 | 5.83 | 41.50b | 2.56 | 6.43 | 4.2 | 24.4 | |||
| 0.031 | 29.41 | 6.08 | 40.31a | 2.75 | 6.39 | 4.3 | 24.6 | |||
| Cultivar | Mo Supply (μmol L−1) | Leaf Concentration (% DM) | |||||
|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | ||
| Experiment 1 | |||||||
| ‘Milton Dark Pink’ | 0.500 | 5.03 | 0.66 | 5.50 | 1.40 | 0.55 | 0.30 |
| 0.250 | 4.83 | 0.57 | 5.38 | 1.34 | 0.52 | 0.30 | |
| 0.125 | 4.82 | 0.63 | 5.34 | 1.31 | 0.51 | 0.29 | |
| ‘Williamsburg Purple’ | 0.500 | 4.87 | 0.54 | 4.89 | 1.09 | 0.35 | 0.27 |
| 0.250 | 4.97 | 0.59 | 4.98 | 1.12 | 0.36 | 0.28 | |
| 0.125 | 5.09 | 0.60 | 5.11 | 1.12 | 0.37 | 0.28 | |
| Experiment 2 | |||||||
| ‘Milton Dark Pink’ | 0.125 | 5.08 | 0.73 | 5.58 | 1.19 | 0.44 | 0.43 |
| 0.063 | 5.17 | 0.69 | 5.65 | 1.16 | 0.42 | 0.42 | |
| 0.031 | 5.12 | 0.72 | 5.53 | 1.19 | 0.45 | 0.42 | |
| ‘Mount Aubisque Purple’ | 0.125 | 4.92 | 0.72a | 5.99 | 1.03 | 0.40 | 0.50 |
| 0.063 | 5.09 | 0.65b | 6.10 | 1.02 | 0.40 | 0.49 | |
| 0.031 | 5.02 | 0.69ab | 6.00 | 1.04 | 0.42 | 0.48 | |
| Cultivar | Mo Supply (μmol L−1) | Leaf Concentration (mg kg−1 DM) | |||||
|---|---|---|---|---|---|---|---|
| Mo | B | Cu | Zn | Mn | Fe | ||
| Experiment 1 | |||||||
| ‘Milton Dark Pink’ | 0.500 | 1.4 | 64.8 | 3.4 | 23.3 | 75.3 | 70.8 |
| 0.250 | 1.2 | 63.5 | 3.2 | 27.5 | 66.3 | 70.3 | |
| 0.125 | 1.5 | 62.5 | 3.2 | 23.3 | 66.5 | 82.0 | |
| ‘Williamsburg Purple’ | 0.500 | 3.7 | 48.8 | 2.7 | 21.3 | 60.5 | 75.3 |
| 0.250 | 3.4 | 50.3 | 2.9 | 23.5 | 58.0 | 74.8 | |
| 0.125 | 2.9 | 48.8 | 3.0 | 21.0 | 57.3 | 73.5 | |
| Experiment 2 | |||||||
| ‘Milton Dark Pink’ | 0.125 | 1.6a | 72.5a | 3.8 | 34.1 | 81.3 | 87.1 |
| 0.063 | 1.3b | 71.3ab | 4.3 | 33.6 | 78.9 | 81.0 * | |
| 0.031 | 1.1c | 70.6b | 4.3 | 34.1 | 73.6 | 83.1 * | |
| ‘Mount Aubisque Purple’ | 0.125 | 1.6a | 70.8a | 3.5 | 28.0 | 74.5 | 84.5 |
| 0.063 | 1.1b | 70.5a | 3.5 | 29.3 | 79.3 | 80.8 | |
| 0.031 | 1.0b | 69.0b | 3.5 | 29.0 | 77.8 | 84.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teeter-Wood, K.R.; Flaherty, E.J.; Donetz, A.J.; Hoover, G.J.; MacDonald, W.N.; Wolyn, D.J.; Shelp, B.J. Improving Boron and Molybdenum Use Efficiencies in Contrasting Cultivars of Subirrigated Greenhouse-Grown Pot Chrysanthemums. Plants 2023, 12, 2348. https://doi.org/10.3390/plants12122348
Teeter-Wood KR, Flaherty EJ, Donetz AJ, Hoover GJ, MacDonald WN, Wolyn DJ, Shelp BJ. Improving Boron and Molybdenum Use Efficiencies in Contrasting Cultivars of Subirrigated Greenhouse-Grown Pot Chrysanthemums. Plants. 2023; 12(12):2348. https://doi.org/10.3390/plants12122348
Chicago/Turabian StyleTeeter-Wood, Katherine R., Edward J. Flaherty, Alyna J. Donetz, Gordon J. Hoover, William N. MacDonald, David J. Wolyn, and Barry J. Shelp. 2023. "Improving Boron and Molybdenum Use Efficiencies in Contrasting Cultivars of Subirrigated Greenhouse-Grown Pot Chrysanthemums" Plants 12, no. 12: 2348. https://doi.org/10.3390/plants12122348