
Kalanchoe sp. Extracts—Phytochemistry, Cytotoxic, and Antimicrobial Activities
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Anticancer Study
2.1.1. Cytotoxic Activity
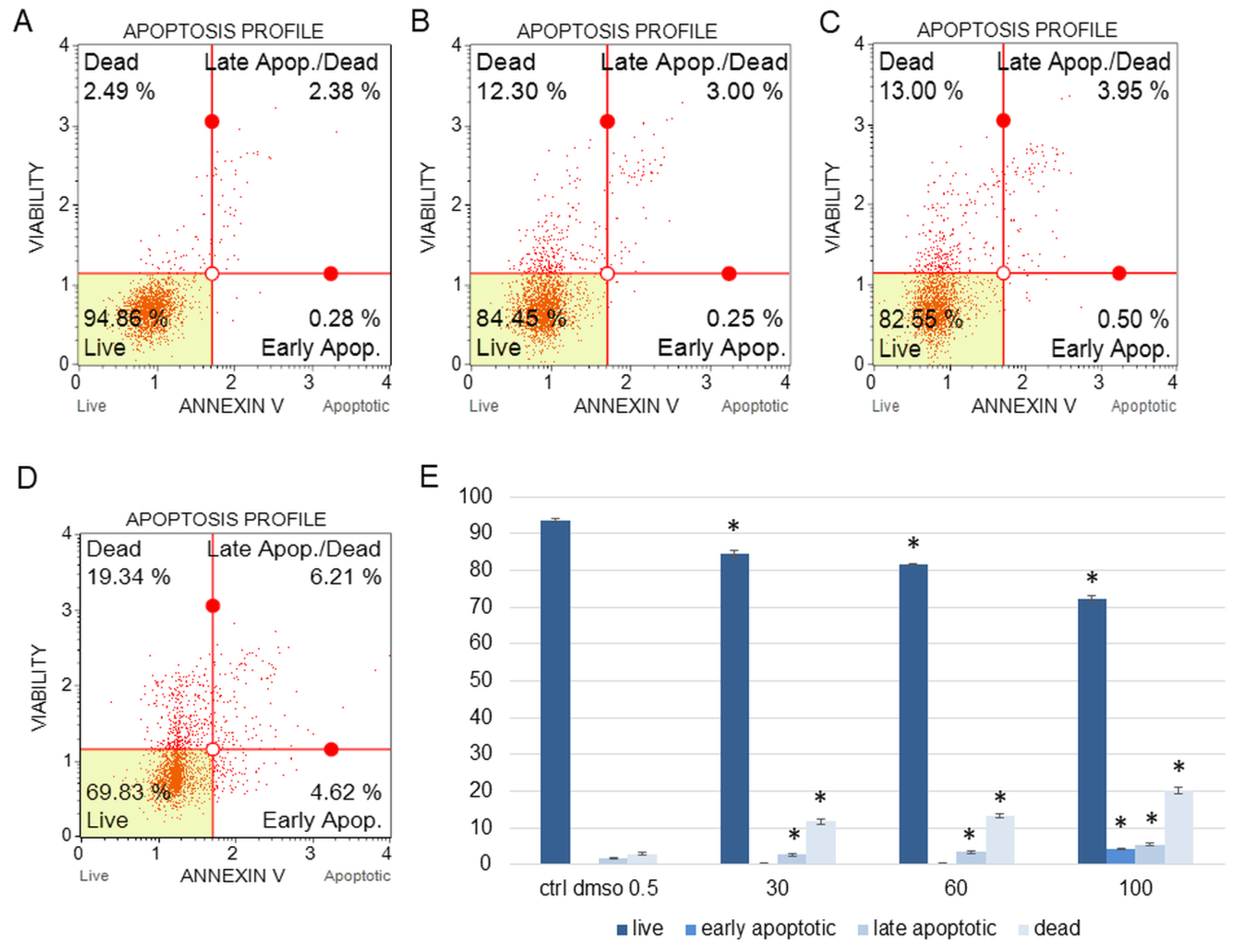
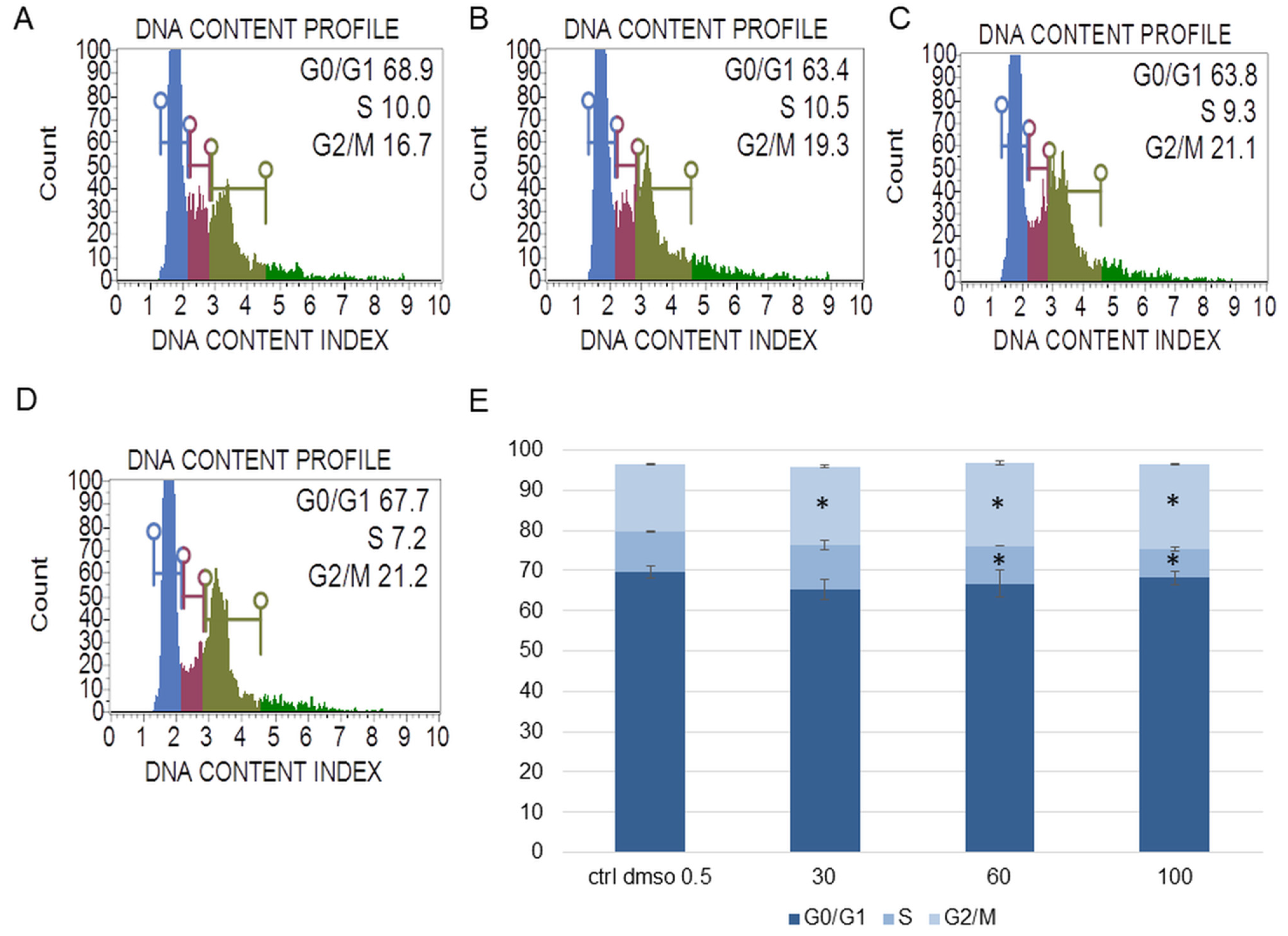
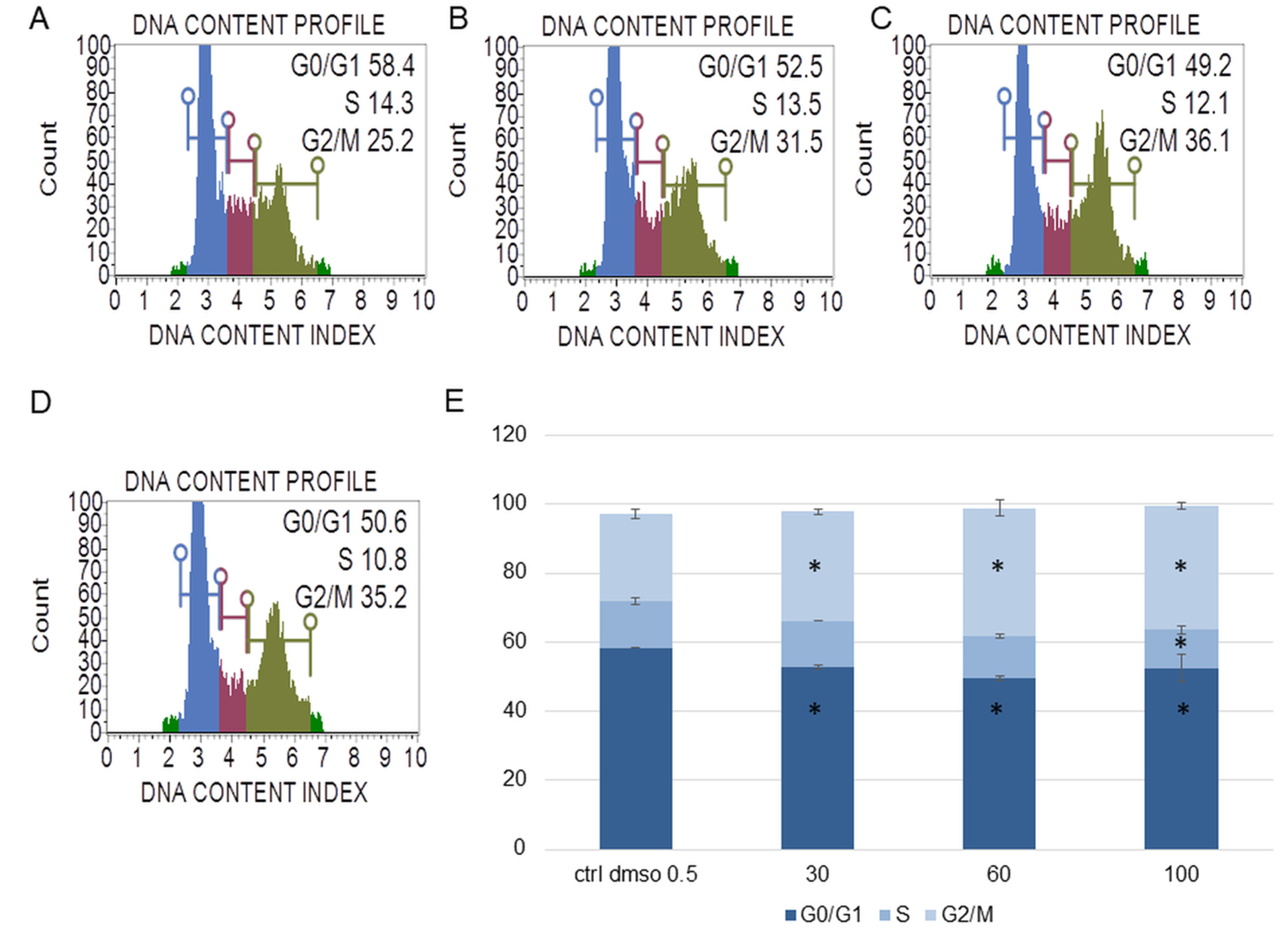
2.1.2. The Effect of the Water Fraction of K. blossfeldiana on Mitochondrial Membrane Potential (MMP), Oxidative Stress Level, and Cell Cycle Arrest in SKOV-3 and Hela Cells
2.2. Microbiological Study
2.3. Phytochemical Profiles of the Water Fractions of K. blossfeldiana and K. pinnata Obtained by LC-QTOF-MS
3. Discussion
4. Materials and Methods
4.1. Preparation of Plant Fractions
4.2. Anticancer Study
4.2.1. Cell Culture
4.2.2. Cytotoxic Assays
MTT Test
Annexin V and Dead Cell Assay
Mitochondrial Potential Assay
Oxidative Stress Assay
Cell Cycle Analysis
4.3. Microbiological Study
4.3.1. Microorganism Species
4.3.2. Antibacterial Assay
4.4. Antioxidant Tests
4.4.1. DPPH Assay
4.4.2. ABTS Assay
4.5. Phytochemical Study of K. blossfeldiana and K. pinnata Water Fractions
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selvakumari, E.; Muthukumaran, R.; Atchaya, S.; Kowsalya, S.; Elavarasan, N. Ethno pharmacological and phyto pharmacological perception on Kalanchoe (Crassulacea)—A comprehensive review. IJPSR 2022, 13, 1428–1440. [Google Scholar]
- Majaz, Q.A.; Tatiya, A.U.; Khurshid, M.; Nazim, S.; Siraj, S.; Allana, A. The miracle plant (Kalanchoe pinnata): A phytochemical and pharmacological review. Int. J. Ayurveda Res. 2011, 2, 1478–1482. [Google Scholar]
- Lans, C.A. Ethanomedicines used in trinidad and tobago for urinary problems and diabetes mellitus. J. Ethnobiol. 2006, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Sen, P.; Dollo, M.; Chaudhury, M.D.; Choudhury, D. Documentation and traditional herbal knowledge of Khamptis of Arunachal Pardesh. IJTK 2008, 7, 438–442. [Google Scholar]
- Milad, R.; El-Ahmady, S.; Singab, A.N. Genus Kalanchoe (Crassulaceae): A review of its ethnomedicinal, botanical, chemical and pharmacological properties. Eur. J. Med. Plants 2014, 4, 86–104. [Google Scholar] [CrossRef]
- Maurice, M. Handbook of African Medicinal Plant; CRC Press: London, UK; Tokyo, Japan, 1993. [Google Scholar]
- Kamboj, A.; Rathour, A.; Kaur, M. Bufadienolides and their medicinal utility: A review. Int. J. Pharm. Sci. 2013, 5, 20–27. [Google Scholar]
- Melero, C.P.; Medarde, M.; San Feliciano, A. A short review on cardiotonic steroids and their aminoguanidine analogues. Molecules 2000, 5, 51–81. [Google Scholar] [CrossRef] [Green Version]
- Schoner, W.; Scheiner-Bobis, G. Endogenous and exogenous cardiac glycosides: Their roles in hypertension, salt metabolism, and cell growth. Am. J. Physiol. Cell Physiol. 2007, 293, 509–536. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Stochmal, A. Bufadienolides of Kalanchoe species: An overview of chemical structure, biological activity and prospects for pharmacological use. Phytochem. Rev. 2017, 16, 1155–1171. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.A.; Muzitano, M.F.; Camargo, L.M.M.; Coutinho, M.A.S. Therapeutic Potential of Kalanchoe Species: Flavonoids and other secondary metabolites. Nat. Prod. Commun. 2008, 3, 2151–2164. [Google Scholar] [CrossRef] [Green Version]
- Shirobokov, V.P.; Evtushenko, A.I.; Lapchik, V.F.; Shirobokov, D.N.; Suptel, E.A. Antiviral activity of representatives of the family Crassulaceae. Antibiotiki 1981, 26, 897–900. [Google Scholar] [PubMed]
- Rossi-Bergmann, B.; Costa, S.S.; Borges, M.B.S.; Da Silva, S.A.; Noleto, G.R.; Souza, M.L.M.; Moraes, V.L.G. Immunosuppressive effect of the aqueous extract of Kalanchoe pinnata in mice. Phytother. Res. 1994, 8, 399–402. [Google Scholar] [CrossRef]
- Singab, A.B.; El-Ahamdy, S.H.; Labib, R.M.; Fekry, S.S. Kalanchoe thrysiflora Harv. and Kalanchoe marmorata Baker; DNA profiling, biological guided fractionation of different extracts; isolation and identification of cytotoxic compounds. J. Appl. Pharm. Sci. 2012, 2, 215–220. [Google Scholar]
- Nahar, K.; Khan, M.G.U.; Rahman, M.S.; Begum, B.; Rashid, M.A. Antimicrobial and cytotoxic activities of Bryophyllum daigremontianum. J. Pharm. Sci. 2008, 7, 99–101. [Google Scholar] [CrossRef] [Green Version]
- Kamgang, R.; Mboumi, R.Y.; Fondjo, A.F.; Tagne, M.A.F.; Mengue N’dille, G.P.R.; Yonkeu, J.N. Antihyperglycaemic potential of the water-ethanol extract of Kalanchoe crenata (Crassulaceae). J. Nat. Med. 2008, 62, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Umbuzeiro-Valent, G.; Roubicek, D.A.; Haebisch, E.M. Mutagenic and antimutagenic evaluation of the juice of the leaves of Bryophyllum calycinum (Kalanchoe pinnata), a plant with antihistamine activity. Environ. Mol. Mutagen. 1999, 33, 325–327. [Google Scholar] [CrossRef]
- Gaind, K.N.; Singla, A.K.; Wallace, J.W. Flavonoid glycoside of Kalanchoe spathulata. Phytochemistry 1981, 20, 530–531. [Google Scholar] [CrossRef]
- Liu, K.C.S.; Yang, S.L.; Roberts, M.F.; Phillipson, J.D. Eupafolin rhamnosides from Kalanchoe gracilis. J. Nat. Prod. 1989, 52, 970–974. [Google Scholar] [CrossRef]
- Muzitano, M.F.; Cruz, E.A.; De Almedia, A.P.; Da Silva, S.A.G.; Kaiser, C.R.; Guette, C.; Rosibergmann, B.; Costa, S.S. Quercitrin: An antileishmanial flavonoid glycoside from Kalanchoe pinnata. Planta Med. 2006, 72, 81–83. [Google Scholar] [CrossRef]
- Singab, A.B.; El-Ahamdy, S.H.; Labib, R.M.; Fekry, S.S. Phenolics from Kalanchoe marmorata Baker, family Crassulaceae. Bull. Fac. Pharm. Cairo Univ. 2011, 49, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Stefanowicz-Hajduk, J.; Asztemborska, M.; Krauze-Baranowska, M.; Godlewska, S.; Gucwa, M.; Moniuszko-Szajwaj, B.; Stochmal, A.; Ochocka, J.R. Identification of flavonoids and bufadienolides and cytotoxic effects of Kalanchoe daigremontiana extracts on human cancer cell lines. Planta Med. 2020, 86, 238–239. [Google Scholar] [CrossRef]
- Anderson, L.A.; Schultz, R.A.; Joubert, J.P.; Prozesky, L.; Kellerman, T.S.; Erasmus, G.L.; Procos, J. Krimpsiekte and acute cardiac glycoside poisoning in sheep caused by bufadienolides from the plant Kalanchoe lanceolata Forsk. Onderstepoort. J. Vet. Res. 1983, 50, 295–300. [Google Scholar]
- Wagner, H.; Fischer, M.; Lotter, H. Isolation and structure determination of daigremontianin, a novel bufadienolide from Kalanchoe daigremontiana. Planta Med. 1985, 2, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, T.; Haruna, M.; Yan, X.Z.; Chang, J.J.; Lee, K.H. Antitumor agents, Bryophyllin B, A novel potent cytotoxic bufadienolide from Bryophyllum pinnatum. J. Nat. Prod. 1989, 52, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Supratman, U.; Fujita, T.; Akiyama, K.; Hayashi, H. Insecticidal compounds from Kalanchoe daigremontiana × tubiflora. Phytochemistry 2001, 58, 311–314. [Google Scholar] [CrossRef]
- Wu, P.L.; Hsu, Y.L.; Wu, T.S.; Bastow, K.F.; Lee, K.H. Kalanchosides A-C, new cytotoxic bufadienolides from the aerial parts of Kalanchoe gracilis. Org. Lett. 2006, 8, 5207–5210. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.C.; Kuo, T.H.; Su, C.R.; Liou, M.J.; Wu, T.S. Cytotoxic principles and α-pyrone ring-opening derivatives of bufadienolides from Kalanchoe hybrida. Tetrahedron 2008, 64, 3392–3396. [Google Scholar] [CrossRef]
- Stefanowicz-Hajduk, J.; Hering, A.; Gucwa, M.; Hałasa, R.; Soluch, A.; Kowalczyk, M.; Stochmal, A.; Ochocka, J.R. Biological activities of leaf extracts from selected Kalanchoe species and their relationship with bufadienolides content. Pharm. Biol. 2020, 58, 732–740. [Google Scholar] [CrossRef]
- Chance, B.; Williams, G.R. The respiratory chain and oxidative phosphorylation. Adv. Enzymol. Relat. Subj. Biochem. 1956, 17, 65–134. [Google Scholar]
- Chen, L.B. Mitochondrial membrane potential in living cells. Ann. Rev. Cell Biol. 1988, 4, 155–181. [Google Scholar] [CrossRef]
- Pieczenik, S.R.; Neustadt, J. Mitochondrial dysfunction and molecular pathways of disease. Exp. Mol. Pathol. 2007, 83, 84–92. [Google Scholar] [CrossRef]
- Lemasters, J.J.; Qian, T.; He, L.; Kim, J.S.; Elmore, S.P.; Cascio, W.E.; Brenner, D.A. Role of mitochondrial inner membrane permeabilization in necrotic cell death, apoptosis, and autophagy. Antioxid. Redox. Signal. 2002, 4, 769–781. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [Green Version]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genestra, M. Oxyl radicals, redox-sensitive signalling cascades and antioxidants. Cell. Signal. 2007, 19, 1807–1819. [Google Scholar] [CrossRef]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.B.d.S.; Casanova, L.M.; Costa, S.S. Bioactive compounds from Kalanchoe genus potentially useful for the development of new drugs. Life 2023, 13, 646. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, H.; Si, N.; Ren, W.; Gao, B.; Li, Y.; Jian, Y.; Xu, M.; Zhao, H.; Bian, B. Metabolites profiling of 10 bufadienolides in human liver microsomes and their cytotoxicity variation in HepG2 cell. Anal. Bioanal. Chem. 2016, 408, 2485–2495. [Google Scholar] [CrossRef]
- Han, L.; Yuan, B.; Shimada, R.; Hayashi, H.; Si, N.; Zhao, H.Y.; Bian, B.; Takagi, N. Cytocidal effects of arenobufagin and hellebrigenin, two active bufadienolide compounds, against human glioblastoma cell line U-87. Int. J. Oncol. 2018, 53, 2488–2502. [Google Scholar] [CrossRef] [Green Version]
- Yeh, J.Y.; Huang, W.J.; Kan, S.F.; Wang, P.S. Effects of bufalin and cinobufagin on the proliferation of androgen dependent and independent prostate cancer cells. Prostate 2003, 54, 112–124. [Google Scholar] [CrossRef]
- Stefanowicz-Hajduk, J.; Gucwa, M.; Moniuszko-Szajwaj, B.; Stochmal, A.; Kawiak, A.; Ochocka, J.R. Bersaldegenin-1,3,5-orthoacetate induces caspase-independent cell death, DNA damage and cell cycle arrest in human cervical cancer HeLa cells. Pharm. Biol. 2021, 59, 54–65. [Google Scholar] [CrossRef]
- Akinpelu, D.A. Antimicrobial activity of Bryophyllum pinnatum leaves. Fitoterapia 2000, 71, 193–194. [Google Scholar] [CrossRef]
- Akinsulire, O.R.; Aibinu, I.E.; Adenipekun, T.; Adelowotan, T.; Odugbemi, T. In vitro antimicrobial activity of crude extracts from plants Bryophyllum pinnatum and Kalanchoe crenata. Afr. J. Tradit. Complement. Altern. Med. 2007, 4, 338–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmawan, A.; Fajriah, M.S. 3′,4′-Dimethoxy quercetin, a flavonol compound isolated from Kalanchoe pinnata. J. App. Pharm. Sci. 2013, 3, 088–090. [Google Scholar]
- Tatsimo, S.J.N.; Tamokou, J.D.; Havyarimana, L.; Csupor, D.; Forgo, P.; Hohmann, J.; Kuiate, J.R.; Tane, P. Antimicrobial and antioxidant activity of Kaempferol rhamnoside derivatives from Bryophyllum pinnatum. BMC Res. Notes 2012, 5, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, A.H.; Olsen, C.E.; Møller, B.L. Flavonoids in flowers of 16 Kalanchoe blossfeldiana varieties. Phytochemistry 2005, 66, 2829–2835. [Google Scholar] [CrossRef]
- Pryce, R.J. Gallic acid as a natural inhibitor of flowering in Kalanchoe blossfeldiana. Phytochemistry 1972, 11, 1911–1918. [Google Scholar] [CrossRef]
- Yun, Y.; Aisyah, L.; Saputra, T.R.; Hakim, A.; Herlina, T.; Julaeha, E.; Zainuddin, A.; Supratman, U. Phenolic compounds from the leaves of Kalanchoe blossfeldiana (Crassulaceae) plant. In Proceedings of the 9th Joint Conference on Chemistry, Semarang, Indonesia, 12–13 November 2014. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Available online: https://www.luminexcorp.com/eu/muse-cell-analyzer/#assays (accessed on 10 May 2023).
- Chomnawang, M.T.; Surassmo, S.; Nukoolkarn, V.S.; Gritsanapan, W. Antimicrobial effects of Thai medicinal plants against acne-inducing bacteria. J. Ethnopharmacol. 2005, 101, 330–333. [Google Scholar] [CrossRef]
- Lekogo, B.M.; Coroller, L.; Mathot, A.G.; Mafart, P.; Leguerinel, I. Modelling the influence of palmitic, palmitoleic, stearic and oleic acids on apparent heat resistance of spores of Bacillus cereus NTCC 11145 and Clostridium sporogenes Pasteur 79.3. Int. J. Food Microbiol. 2010, 141, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Kula, M.; Majdan, M.; Radwanska, A.; Nasal, A.; Hałasa, R.; Głód, D.; Matkowski, A.; Krauze-Baranowska, M. Chemical composition and biological activity of the fruits from Lonicera caerulea var. edulis ‘Wojtek’. Acad. J. Med. Plants 2013, 1, 141–148. [Google Scholar]
- Kwon, S.H.; Wang, Z.; Hwang, S.H.; Kang, Y.H.; Lee, J.Y.; Lim, S.S. Comprehensive evaluation of the antioxidant capacity of Perilla frutescens leaves extract and isolation of free radical scavengers using step-wise HSCCC guided by DPPH-HPLC. Int. J. Food Prop. 2017, 20, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Bułakowska, A.; Sławiński, J.; Hałasa, R.; Hering, A.; Gucwa, M.; Ochocka, J.R.; Stefanowicz-Hajduk, J. An in vitro antimicrobial, anticancer and antioxidant activity of N–[(2–Arylmethylthio)phenylsulfonyl]cinnamamide derivatives. Molecules 2023, 28, 3087. [Google Scholar] [CrossRef] [PubMed]
- Krzyżanowska-Kowalczyk, J.; Pecio, Ł.; Mołdoch, J.; Ludwiczuk, A.; Kowalczyk, M. Novel phenolic constituents of Pulmonaria officinalis L. LC-MS/MS comparison of spring and autumn metabolite profiles. Molecules 2018, 23, 2277. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; Jüppner, J.; Bajdzienko, K.; Giavalisco, P. Protocol: A fast, comprehensive and reproducible one-step extraction method for the rapid preparation of polar and semi-polar metabolites, lipids, proteins, starch and cell wall polymers from a single sample. Plant Methods 2016, 12, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liudvytska, O.; Ponczek, M.B.; Ciesielski, O.; Krzyżanowska-Kowalczyk, J.; Kowalczyk, M.; Balcerczyk, A.; Kolodziejczyk-Czepas, J. Rheum rhaponticum and Rheum rhabarbarum extracts as modulators of endothelial cell inflammatory response. Nutrients 2023, 15, 949. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Ruttkies, C.; Schymanski, E.L.; Wolf, S.; Hollender, J.; Neumann, S. MetFrag relaunched: Incorporating strategies beyond in silico fragmentation. J. Cheminf. 2016, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted metabolomics strategies—Challenges and emerging directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K. b. Water Fraction | K. p. Water Fraction | K. d. Water Fraction * | K. b. Dichlorometh. Fraction | K. p. Dichlorometh. Fraction | K. d. Dichlorometh. Fraction * | Vinblastine Sulphate | |
|---|---|---|---|---|---|---|---|
| HeLa | 28.28 ± 2.76 | 79.91 ± 1.82 | >100 | >100 | >100 | 6.42 ± 0.34 | 0.005 ± 0.0004 |
| SKOV-3 | 32.51 ± 0.69 | 64.89 ± 1.85 | >100 | >100 | 32.10 ± 0.73 | 5.42 ± 0.16 | 0.008 ± 0.0003 |
| MCF-7 | >100 | >100 | >100 | >50 | >100 | 8.02 ± 0.13 | 0.007 ± 0.0008 |
| A375 | 49.44 ± 2.43 | >100 | >100 | 95.11 ± 6.35 | 49.7 ± 1.30 | 7.72 ± 0.44 | 0.008 ± 0.0003 |
| IC50 (µg/mL) | ||
|---|---|---|
| Water Fraction of K. blossfeldiana | Ascorbic Acid | |
| DPPH | 9.44 ± 0.06 | 13.1 ± 0.17 |
| ABTS | 3.17 ± 0.1 | 5.26 ± 0.04 |
| Microorganism Species | K. blossfeldiana Water Fraction | K. pinnata Water Fraction | K. daigremontiana Water Fraction | K. blossfeldiana Dichlorometh. Fraction | K. pinnata Dichlorometh. Fraction | K. daigremontiana Dichlorometh. Fraction | Ampicillin | Ket. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MIC | |
| S. β-hemolyzing group A PCM465 | >1.0 | >1.0 | >2.0 | >2.0 | 4.5 | 9.0 | 5.0 | 5.0 | 2.0 | >2.0 | 2.0 | 2.0 | 3.13 × 10−4 | n.t. |
| S. β-hemolyzing group G | >1.0 | >1.0 | >2.0 | >2.0 | 9.0 | 9.0 | >5.0 | >5.0 | 2.0 | >2.0 | 2.0 | 2.0 | 1.6 × 10−4 | n.t. |
| C. diphtheriae | >1.0 | >1.0 | >2.0 | >2.0 | 4.5 | 9.0 | 5.0 | 5.0 | >2.0 | >2.0 | 2.0 | 2.0 | 1.6 × 10−4 | n.t. |
| S. aureus ATCC6538 | 0.032 | >1.0 | 0.064 | >2.0 | 2.25 | >9.0 | >5.0 | >5.0 | 2.0 | >2.0 | >2.0 | >2.0 | 8 × 10−5 | n.t. |
| S. epidermidis ATCC14990 | 0.016 | >1.0 | 0.032 | >2.0 | 0.064 | >9.0 | 0.064 | >5.0 | 2.0 | >2.0 | >2.0 | >2.0 | 3.13 × 10−4 | n.t. |
| H. pylori ATCC43504 | >1.0 | >1.0 | >2.0 | >2.0 | >9.0 | >9.0 | 1.25 | >5.0 | 15 | >15 | >2.0 | >2.0 | 3.2 × 10−3 | n.t. |
| C. acnes ATCC6919 | 0.5 | >0.5 | 1.0 | >1.0 | 4.5 | >4.5 | >2.5 | >2.5 | 15 | >15 | 1.0 | >1.0 | 0.032 | n.t. |
| S. equinus ATCC15351 | 0.5 | >0.5 | 1.0 | >1.0 | 4.5 | >4.5 | >2.5 | >2.5 | 15 | >15 | 1.0 | >1.0 | 0.016 | n.t. |
| C. bifermentans ATCC638 | 0.25 | >0.5 | 1.0 | >1.0 | 4.5 | >4.5 | 2.5 | >2.5 | 1 | >15 | 0.25 | >1.0 | 0.016 | n.t. |
| C. sporogenes ATCC19404 | 0.5 | >0.5 | 1.0 | >1.0 | 4.5 | >4.5 | 2.5 | >2.5 | 2 | >15 | 0.125 | >1.0 | <6.3 × 10−5 | n.t. |
| C. albicans ATCC10231 | >1.0 | >1.0 | >2.0 | >2.0 | 9.0 | 9.0 | 5.0 | 5.0 | 2.0 | >2.0 | 2.0 | 2.0 | n.t. | >0.125 |
| Compound Categories | K. blossfeldiana Water Fraction | K. pinnata Water Fraction | Total Number of Compounds in Each Group |
|---|---|---|---|
| Unidentified | 55 | 26 | 68 |
| Flavonol glycoside | 8 | 25 | 31 |
| Acyclic alcohol glycoside | 9 | 14 | 16 |
| Benzoic acid derivative | 8 | 12 | 14 |
| Phenylpropanoid derivative | 8 | 11 | 13 |
| Gallic acid derivative | 12 | 6 | 13 |
| Organic acid | 11 | 12 | 12 |
| Megastigmane glycoside | 4 | 10 | 10 |
| Flavanol | 7 | 0 | 7 |
| Dimeric proanthocyanidin | 6 | 2 | 6 |
| Acyclic nitrile glycoside | 3 | 5 | 5 |
| Phenol derivative | 2 | 1 | 2 |
| Aminoacid | 3 | 3 | 3 |
| Acyclic acid glycoside | 2 | 0 | 2 |
| Phenylethane glycoside | 2 | 0 | 2 |
| Monoterpene derivative | 2 | 0 | 2 |
| Iridoid glycoside | 1 | 1 | 2 |
| Carbohydrate | 1 | 1 | 1 |
| Acetophenone derivative | 1 | 1 | 1 |
| Sesquiterpenoid derivative | 1 | 1 | 1 |
| Dimeric iridoid derivative | 1 | 0 | 1 |
| Flavanone derivative | 0 | 1 | 1 |
| Bicyclo[3.1.1] glycoside | 1 | 0 | 1 |
| Dihydrochalcone derivative | 1 | 0 | 1 |
| Fatty acid glycoside | 1 | 0 | 1 |
| Flavonol | 0 | 1 | 1 |
| Lipid | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanowicz-Hajduk, J.; Hering, A.; Kowalczyk, M.; Hałasa, R.; Gucwa, M.; Ochocka, J.R. Kalanchoe sp. Extracts—Phytochemistry, Cytotoxic, and Antimicrobial Activities. Plants 2023, 12, 2268. https://doi.org/10.3390/plants12122268
Stefanowicz-Hajduk J, Hering A, Kowalczyk M, Hałasa R, Gucwa M, Ochocka JR. Kalanchoe sp. Extracts—Phytochemistry, Cytotoxic, and Antimicrobial Activities. Plants. 2023; 12(12):2268. https://doi.org/10.3390/plants12122268
Chicago/Turabian StyleStefanowicz-Hajduk, Justyna, Anna Hering, Mariusz Kowalczyk, Rafał Hałasa, Magdalena Gucwa, and J. Renata Ochocka. 2023. "Kalanchoe sp. Extracts—Phytochemistry, Cytotoxic, and Antimicrobial Activities" Plants 12, no. 12: 2268. https://doi.org/10.3390/plants12122268