
Bohemian Knotweed Reynoutria × bohemica Chrtek et Chrtková Seems Not to Rely Heavily on Allelopathy for Its Persistence in Invaded Sites in the Southwest Part of the Zagreb, Croatia


Abstract
:1. Introduction
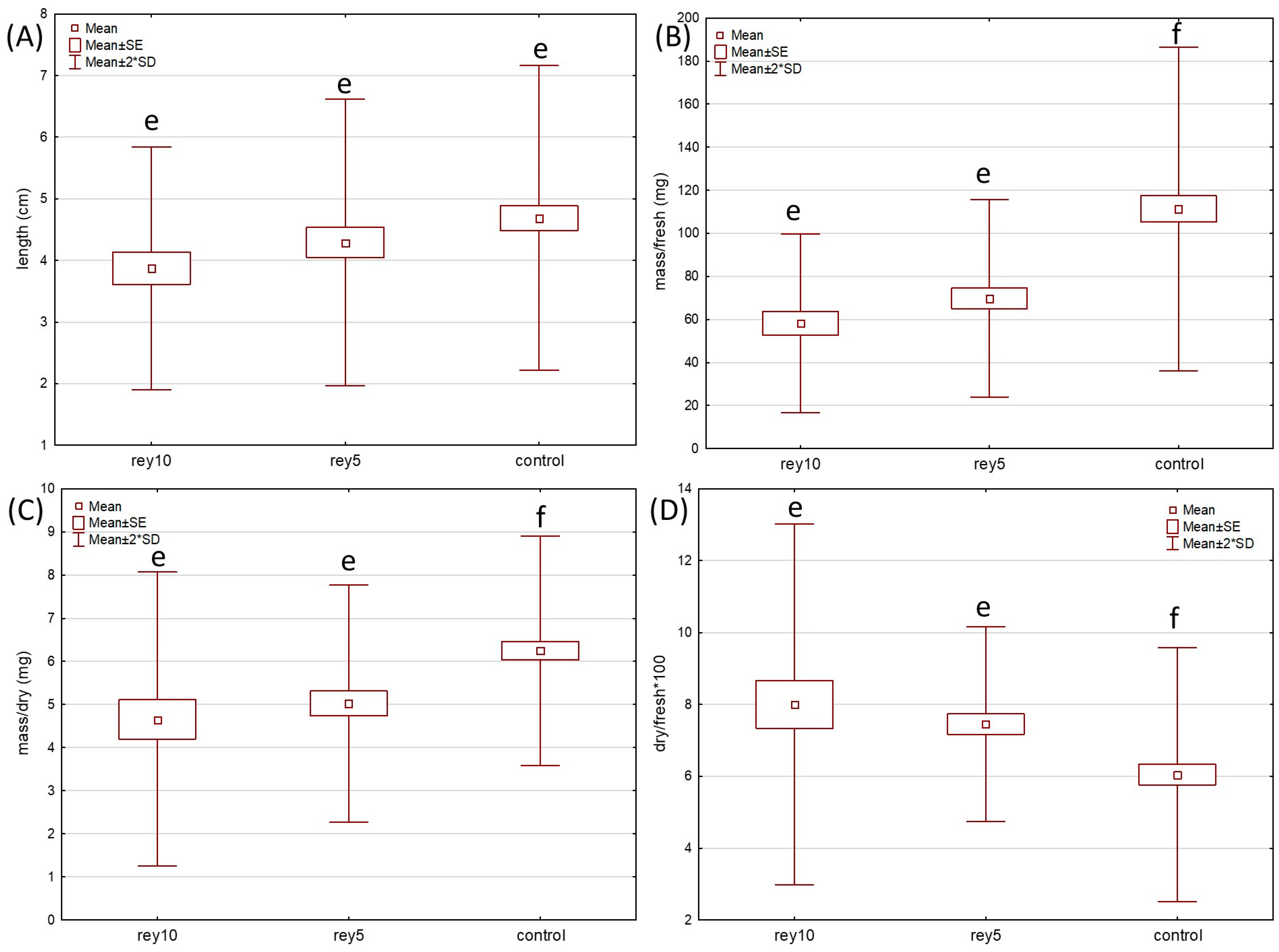
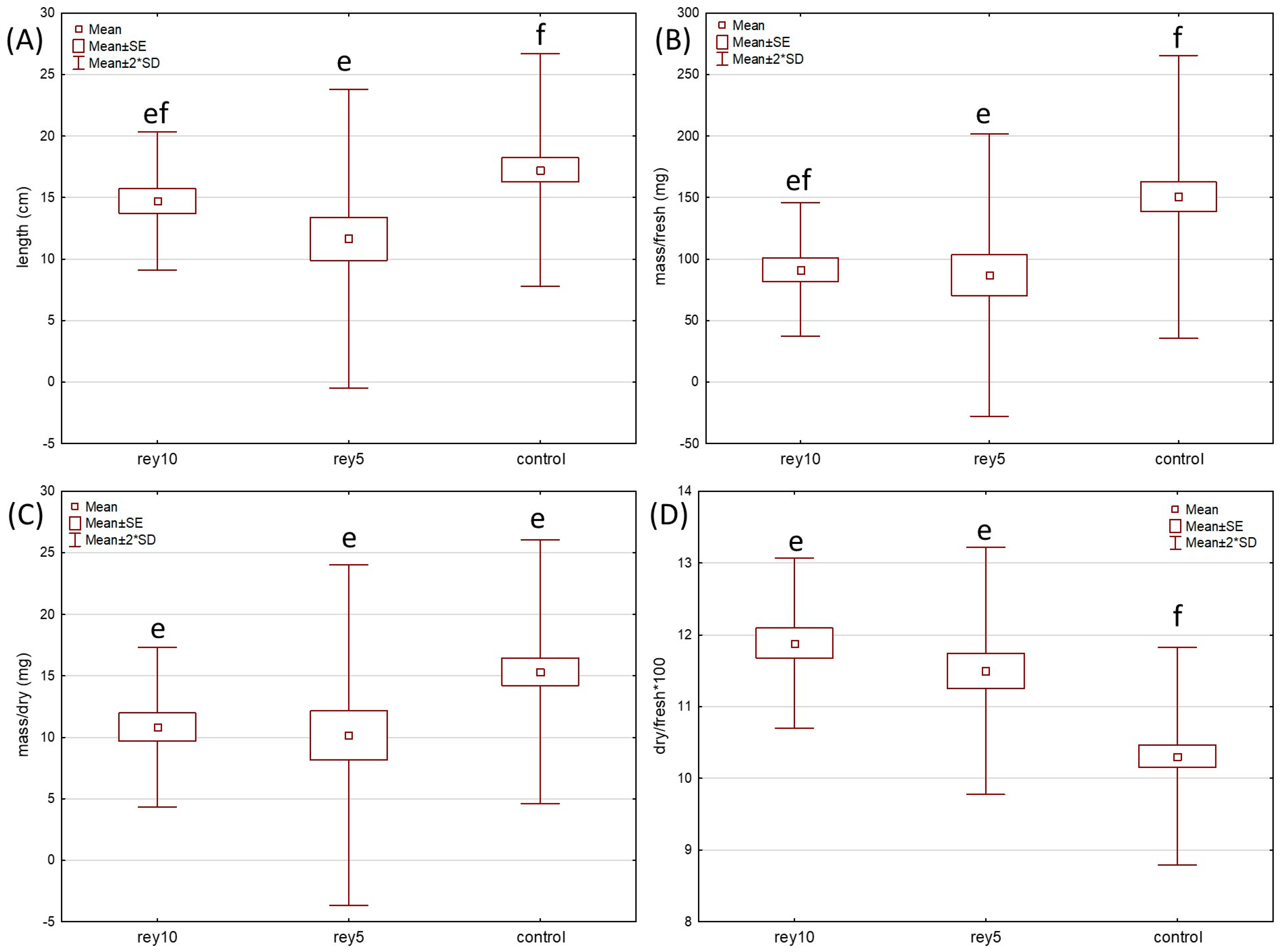
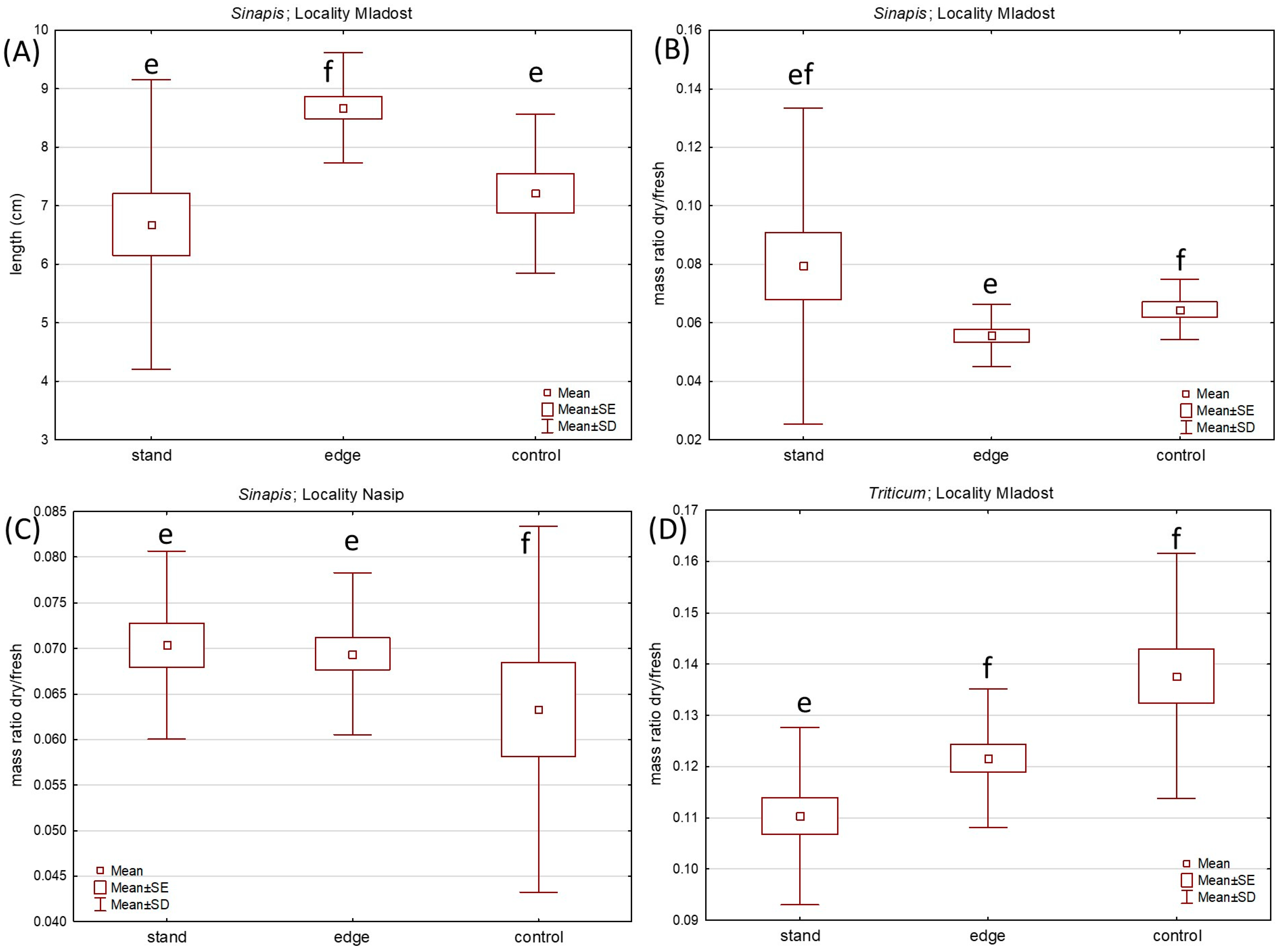
2. Results
2.1. Germination and Growth Test
2.2. Soil Chemical Characteristics Test
3. Discussion
4. Materials and Methods
4.1. Germination Test in Petri Dishes with Knotweed Leaf Exudate
4.2. Germination and Growth Tests in the Pots with Soil with Knotweed Leaf Exudate
4.3. Germination and Growth Tests in the Pots with Soil Sampled In Situ at the Knotweed-Invaded Sites
4.4. Chemical Characteristics of Soil Sampled In Situ at the Knotweed-Invaded Sites
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murrell, C.; Gerber, E.; Krebs, C.; Parepa, M.; Schanffner, U.; Bossdorf, O. Invasive knotweed affects native plants through allelopathy. Am. J. Bot. 2011, 98, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Scalera, R.; Genovesi, P.; Essl, F.; Rabitsch, W. The Impacts of Alien Species in Europe; EAA Technical Report; European Environment Agency: Luxembourg, 2012; pp. 1–113.
- Bassett, I.; Paynter, Q.; Beggs, J.R. Invasive Alternanthera philoxeroides (alligator weed) associated with increased fungivore dominance in Coleoptera on decomposing leaf litter. Biol. Invasions 2011, 13, 1377–1385. [Google Scholar] [CrossRef]
- Kohyt, J.; Skubała, P. Communities of mites (Acari) in litter and soil under the invasive red oak (Quercus rubra L.) and native pedunculate oak (Q. robur L.). Biol. Lett. 2013, 50, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Inderjit; Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel Weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kaur, S.; Kohli, R.K. Novel weapon hypothesis for the successful establishment of invasive plants in alien environments. In Invasive Plant Ecology, 1st ed.; Jose, S., Singh, H.P., Batish, D.R., Kohli, R.K., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 19–28. [Google Scholar] [CrossRef]
- Callaway, R.M.; Cipollini, D.; Barto, K.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Stinson, K.; Klironomos, J. Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 2008, 89, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Inderjit; Evans, H.; Crocoll, C.; Bajpai, D.; Kaur, R.; Feng, Y.L.; Silva, C.; Treviño Carreón, J.; Valiente-Banuet, A.; Gershenzon, J.; et al. Volatile chemicals from leaf litter are associated with invasiveness of a neotropical weed in Asia. Ecology 2011, 92, 316–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Chen, X.; Ding, L.; Kong, C.H. Allelopathy and Allelochemicals in Grasslands and Forests. Forests 2023, 14, 562. [Google Scholar] [CrossRef]
- Meiners, S.J.; Kong, C.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant Ecol. 2012, 213, 1221–1227. [Google Scholar] [CrossRef]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Liu, Z.; Wang, Z.; Pang, W.; Zhang, L.; Wen, Z.; Zhao, Y.; Sun, J.; Wang, Z.; Yang, C. Effects of autotoxicity and allelopathy on seed germination and seedling growth in Medicago truncatula. Front. Plant Sci. 2022, 13, 908426. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Nilsson, M.C.; Gallet, C.; Zackrisson, O. An ecosystem-level perspective of allelopathy. Biol. Rev. 1998, 73, 305–319. [Google Scholar] [CrossRef]
- Jin, H.; Guo, H.; Yang, X.; Xin, A.; Liu, H.; Qin, B. Effect of allelochemicals, soil enzyme activity and environmental factors from Stellera chamaejasme L. on rhizosphere bacterial communities in the northern Tibetan Plateau. Arch. Agron. Soil Sci. 2022, 68, 547–560. [Google Scholar] [CrossRef]
- Kalisz, S.; Kivlin, S.N.; Bialic-Murphy, L. Allelopathy is pervasive in invasive plants. Biol. Invasions 2021, 23, 367–371. [Google Scholar] [CrossRef]
- Huang, F.; Lankau, R.; Peng, S. Coexistence via coevolution driven by reduced allelochemical effects and increased tolerance to competition between invasive and native plants. New Phytol. 2018, 218, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Flora Croatica Database Allochthonous Plants. Available online: https://hirc.botanic.hr/fcd/InvazivneVrste/ShowResults.aspx?hash=636268493 (accessed on 14 February 2023).
- Nikolić, T.; Mitić, B.; Boršić, I. Flora Hrvatske: Invazivne Biljke, 1st ed.; Alfa d.o.o.: Zagreb, Croatia, 2014; pp. 1–296. [Google Scholar]
- Brabec, J.; Pyšek, P. Establishment and survival of three invasive taxa of the genus Reynoutria (Polygonaceae) in mesic mown meadows: A field experimental study. Folia Geobot. 2000, 35, 27–42. [Google Scholar] [CrossRef]
- Urgenson, L.S.; Reichard, S.H.; Halpern, C.B. Community and ecosystem consequences of giant knotweed (Polygonum sachalinense) invasion into riparian forests of western Washington, USA. Biol. Conserv. 2009, 142, 1536–1541. [Google Scholar] [CrossRef]
- Brock, J.; Wade, M. Regeneration of Japanese knotweed (Fallopia japonica) from rhizome and stems: Observation from greenhouse trials. In Proceedings of the 9th International Symposium on the Biology of Weed, Dijon, France, 16–18 September 1992. [Google Scholar]
- Vastano, B.C.; Chen, Y.; Zhu, N.; Ho, C.T.; Zhou, Z.; Rosen, R.T. Isolation and Identification of Stilbenes in Two Varieties of Polygonum cuspidatum. J. Agric. Food Chem. 2000, 48, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Koce, J.D.; Soln, K. Phytotoxic effects of Fallopia japonica and F.x bohemica leaves. Phyton-Ann. REI Bot. 2018, 57, 47–58. [Google Scholar]
- Siemens, T.J.; Blossey, B. An evaluation of mechanisms growth and survival of two native species in invasive Bohemian knotweed (Fallopia x bohemica, Polygonaceae). Am. J. Bot. 2007, 94, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Gerber, E.; Krebs, C.; Murrell, C.; Moretti, M.; Rocklin, R.; Schaffner, U. Exotic invasive knotweeds (Fallopia spp.) negatively affect native plant and invertebrate assemblages in European riparian habitats. Biol. Conserv. 2008, 141, 646–654. [Google Scholar] [CrossRef]
- Kappes, H.; Lay, R.; Topp, W. Changes in different trophic levels of litter-dwelling macrofauna associated with giant knotweed invasion. Ecosystems 2007, 10, 734–744. [Google Scholar] [CrossRef]
- Maerz, J.C.; Blossey, B.; Nuzzo, V. Green frogs show reduced foraging success in habitats invaded by Japenese knotweed. Biodivers. Conserv. 2005, 14, 2901–2911. [Google Scholar] [CrossRef]
- Abgrall, C.; Forey, E.; Mignot, L.; Chauvat, M. Invasion by Fallopia japonica alters soil food webs through secondary metabolites. Soil Biol. Biochem. 2018, 127, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Neupert, M.; Margerie, P.; Forey, E.; Chauvat, M.; Bureau, F.; Aubert, M.; Prével, S.; Langlois, E.; Vincenot, L. The Best of BothWorlds? Hybridization Potentiates Exotic Bohemian Knotweed’s (Reynoutria × bohemica) Impacts on Native Plant and Faunal Communities. Biol. Life Sci. Forum 2021, 2, 20. [Google Scholar] [CrossRef]
- Tiébré, M.-S.; Saad, L.; Mahy, G. Landscape dynamics and habitat selection by the alien invasive Fallopia (Polygonaceae) in Belgium. Biodivers. Conserv. 2008, 17, 2357–2370. [Google Scholar] [CrossRef] [Green Version]
- Gillies, S.; Clements, D.R.; Grenz, J. Knotweed (Fallopia spp.) Invasion of North America Utilizes Hybridization, Epigenetics, Seed Dispersal (Unexpectedly), and an Arsenal of Physiological Tactics. Invasive Plant Sci. Manag. 2016, 9, 71–80. [Google Scholar] [CrossRef]
- Williams, F.; Eschen, R.; Harris, A.; Djeddour, D.; Pratt, C.; Shaw, R.S.; Varia, S.; Lamontagne-Godwin, J.D.; Thomas, S.E.; Murphy, S.T. The Economic Cost of Invasive Non-Native Species on Great Britain, 1st ed.; CABI Head Office: Wallingford, UK, 2010; pp. 33–40. [Google Scholar]
- Costello, K.E.; Scalera, R.; Nunes, A.L.; Smith, K. An Introduction to the EU Regulation on Invasive Alien Species; Publications Office of the European Union: Luxemburg, 2022; p. 22.
- Kettunen, M.; Genovesi, P.; Gollasch, S.; Pagad, S.; Starfinger, U.; Ten Brink, P.; Shine, C. Technical Support to EU Strategy on Invasive Species (IS)-Assessment of the Impacts of IS in Europe and the EU (Final Module Report for the European Commission); Institute for European Environmental Policy (IEEP): Bruxelles, Belgium, 2008; p. 131. [Google Scholar]
- Chrtek, J.; Chrtková, A. Reynoutria × bohemica, novy krizenec z celedi Rdesnovitych. Cas. Nar. Muz. 1983, 152, 120. [Google Scholar]
- Bailey, J.P.; Wisskirchen, R. The distribution and origins of Fallopia × bohemica (Polygonaceae) in Europe. Nord. J. Bot. 2004, 24, 173–199. [Google Scholar] [CrossRef]
- Vuković, N.; Šegota, V.; Alegro, A.; Koletić, N.; Rimac, A.; Dekanić, S. “Flying under the radar”–how misleading distributional data led to wrong appreciation of knotweeds invasion (Reynoutria spp.) in Croatia. BioInvasions Rec. 2019, 8, 175–189. [Google Scholar] [CrossRef]
- Mandák, B.; Pyšek, P.; Bímová, K. History of the invasion and distribution of Reynoutria taxa in the Czech Republic: A hybrid spreading faster than its parents. Preslia 2004, 76, 15–64. [Google Scholar]
- Parepa, M.; Fischer, M.; Krebs, C.; Bossdorf, O. Hybridization increases invasive knotweed success. Evol. Appl. 2014, 7, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.H.; Seiger, L.A. Japanese Knotwees. Biological Control of Invasive Plants in the Eastern United States, 1st ed.; USDA, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2002; pp. 159–166.
- Gioria, M.; Osborne, B. Similarities in the impact of three large invasive plant species on soil seed bank communities. Biol. Invasions 2010, 12, 1671–1683. [Google Scholar] [CrossRef]
- Inderjit; Callaway, R.M. Experimental designs for the study of allelopathy. Plant Soil 2003, 56, 1–11. [Google Scholar] [CrossRef]
- Moravcová, L.; Pyšek, P.; Jarošík, V.; Zákravský, P. Potential phytotoxic and shading effects of invasive Fallopia (Polygonaceae) taxa on the germination of dominant native species. NeoBiota 2011, 9, 31–47. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, K.O.; Rinella, M. Comparing susceptibility of eastern and western US grasslands to competition and allelopathy from spotted knapweed [Centaurea stoebe L. subsp. micranthos (Gugler) Hayek]. Plant Ecol. 2011, 212, 821–828. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. The ecological importance of allelopathy. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 25–45. [Google Scholar] [CrossRef]
- Orr, S.P.; Rudgers, J.A.; Clay, K. Invasive plants can inhibit native tree seedlings: Testing potential allelopathic mechanisms. Plant Ecol. 2005, 181, 153–165. [Google Scholar] [CrossRef]
- Kaur, R.; Callaway, R.M.; Inderjit. Soils and the conditional allelopathic effects of a tropical invader. Soil Biol. Biochem. 2014, 78, 316–325. [Google Scholar] [CrossRef]
- Gunarathne, R.; Perera, G. Does the invasion of Prosopis juliflora cause the die-back of the native Manilkara hexandra in seasonally dry tropical forests of Sri Lanka. Trop. Ecol. 2016, 57, 475–488. [Google Scholar]
- Mincheva, T.; Barni, E.; Siniscalo, C. From plant traits to invasion success: Impacts of the alien Fallopia japonica (Houtt.) Ronse Decraene on two native grassland species. Plant Biosyst. 2016, 150, 1348–1357. [Google Scholar] [CrossRef] [Green Version]
- Parepa, M.; Schaffner, U.; Bossdorf, O. Sources and modes of action invasive knotweed allelopathy: The effects of leaf litter and trained soil on the germination and growth of native plants. NeoBiota 2012, 13, 15–30. [Google Scholar] [CrossRef]
- Thelen, G.C.; Vivanco, J.M.; Newingham, B.; Good, W.; Bais, H.P.; Landres, P.; Caesar, A.; Callaway, R.M. Insect herbivory stimulates allelopathic exudation by an invasive plant and the suppression of natives. Ecol. Lett. 2005, 8, 209–217. [Google Scholar] [CrossRef]
- Vrchotová, N.; Šerá, B. Allelopathic properties of knotweeds rhizome extracts. Plant Soil Environ. 2008, 54, 301–303. [Google Scholar] [CrossRef] [Green Version]
- Loydi, A.; Donath, T.W.; Eckstein, R.L.; Otte, A. Non-native species litter reduces germination and growth of resident forbs and grasses: Allelopathic, osmotic or mechanical effects? Biol. Invasions 2015, 17, 581–595. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Gruber, W.; Meerts, P. Invasion by Fallopia japonica increases topsoil mineral nutrient concentrations. Ecoscience 2007, 14, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Maurel, N.; Salmon, S.; Ponge, J.F.; Machon, N. Does the invasive Reynoutria japonica have an impact on soil and flora in urban wastelands? Biol. Invasions 2010, 12, 1709–1719. [Google Scholar] [CrossRef] [Green Version]
- Dassonville, N.; Vanderhoeven, S.; Vanparys, V.; Hayez, M.; Gruber, W.; Meerts, P. Impacts of Alien Invasive Plants on Soil Nutrients Are Correlated with Initial Site Conditions in NW Europe. Oecologia 2008, 157, 131–140. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Kaur, H.; Kaur, R.; Kaur, S.; Baldwin, I.T.; Inderjit. Taking ecological function seriously: Soil microbial communities can obviate allelopathic effects of released metabolites. PLoS ONE 2009, 4, e4700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, C.H.; Zhang, S.Z.; Li, Y.H.; Xia, Z.C.; Yang, X.F.; Meiners, S.J.; Wang, P. Plant neighbor detection and allelochemical response are driven by root-secreted signaling chemicals. Nat. Commun. 2018, 9, 3867. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Cheng, H.F.; Kong, C.H.; Meiners, S.J. Intra-specific kin recognition contributes to inter-specific allelopathy: A case study of allelopathic rice interference with paddy weeds. Plant Cell Environ. 2021, 44, 3709–3721. [Google Scholar] [CrossRef]
- Bímová, K.M.; Bohumil, M.; Kasparová, I. How does Reynoutria invasion fit the various theories of invisibility? J. Veg. Sci. 2004, 15, 495–504. [Google Scholar] [CrossRef]
- Siddiqui, S.; Bhardwaj, S.; Khan, S.S.; Meghvanshi, M.K. Allelopathic Effect of Different Concentration of Water Extract of Prosopsis juliflora Leaf on Seed Germination and Radicle Length of Wheat (Triticum aestivum Var-Lok-1). Am. Eurasian J. Sci. Res. 2009, 4, 81–84. [Google Scholar]
- Pannacci, E.; Pettorossi, D.; Tei, F. Phytotoxic effects of aqueous extracts of sunflower on seed germination and growth of Sinapis alba L., Triticum aestivum L. and Lolium multiflorum Lam. Allelopath. J. 2013, 32, 23–36. [Google Scholar] [CrossRef]
- Bączek, P.; Halarewicz, A. Allelopathic Effect of Black Cherry (Prunus serotina Ehrh.) on Early Growth of White Mustard (Sinapis alba L.) and Common Buckwheat (Fagopyrum esculentum Moench): Is the Invader a Threat to Restoration of Fallow Lands? Agronomy 2022, 12, 2103. [Google Scholar] [CrossRef]
- Ercisli, S.; Esitken, A.; Turkkal, C.; Orhan, E. The allelopathic effects of juglone and walnut leaf extracts on yield, growth, chemical and PNE compositions of strawberry cv. Fern. Plant Soil Environ. 2005, 51, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Abdul Raoof, K.M.; Siddiqui, M.B. Allelopathic effect of aqueous extracts of different parts of Tinospora cordifolia (Willd.) Miers on some weed plants. J. Agric. Ext. Rural Dev. 2012, 4, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Szabó, K.; Szabó, P.; Halbritter, R.A. Allelopathic Effects of Cirsium arvense (L.) Scop. in Hungary. Bulg. J. Agric. Sci. 2015, 21, 1012–1021. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Exudate Concentrations | Control | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Replication | 0.01 g/mL (PD) | 0.03 g/mL (PD) | 0.05 g/mL (PD) | 0.05 g/mL (Pot) | 0.1 g/mL (Pot) | C (PD) | C (Pot) | |||||||
| S | T | S | T | S | T | S | T | S | T | S | T | S | T | |
| 1 | 19 | 11 | 20 | 14 | 17 | 4 | 9 | 5 | 7 | 1 | 20 | 15 | 12 | 6 |
| 2 | 20 | 10 | 19 | 13 | 19 | 8 | 7 | 4 | 0 | 3 | 20 | 17 | 12 | 8 |
| 3 | 20 | 12 | 18 | 13 | 19 | 7 | 7 | 3 | 7 | 4 | 19 | 17 | 14 | 9 |
| % germinated | 98.3 | 55.0 | 95.0 | 66.7 | 91.7 | 31.7 | 45.1 | 24.0 | 27.5 | 15.7 | 98.3 | 96.1 | 74.5 | 45.1 |
| % of the control | 100 | 57.2 | 96.6 | 69.4 | 93.3 | 33.0 | 60.5 | 53.2 | 36.9 | 34.8 | ||||
| Locality | Position | pH | Soil Organic Matter (%) | Humus (%) |
|---|---|---|---|---|
| Mladost | control | 7.69 * | 11.33 | 8.74 |
| Mladost | edge | 7.81 | 7.19 | 5.33 |
| Mladost | stand | 7.90 * | 7.50 | 6.90 |
| Kajzerica | control | 7.92 | 8.79 | 7.97 |
| Kajzerica | edge | 7.75 | 8.62 | 7.78 |
| Kajzerica | stand | 7.87 | 8.19 | 7.69 |
| Nasip | control | 7.88 | 4.92 | 4.38 |
| Nasip | edge | 7.86 | 4.88 | 5.26 |
| Nasip | stand | 8.00 | 4.07 | 4.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levačić, D.; Perković, L.; Vuković, N.; Jelaska, S.D. Bohemian Knotweed Reynoutria × bohemica Chrtek et Chrtková Seems Not to Rely Heavily on Allelopathy for Its Persistence in Invaded Sites in the Southwest Part of the Zagreb, Croatia. Plants 2023, 12, 2222. https://doi.org/10.3390/plants12112222
Levačić D, Perković L, Vuković N, Jelaska SD. Bohemian Knotweed Reynoutria × bohemica Chrtek et Chrtková Seems Not to Rely Heavily on Allelopathy for Its Persistence in Invaded Sites in the Southwest Part of the Zagreb, Croatia. Plants. 2023; 12(11):2222. https://doi.org/10.3390/plants12112222
Chicago/Turabian StyleLevačić, Damjana, Lucia Perković, Nina Vuković, and Sven D. Jelaska. 2023. "Bohemian Knotweed Reynoutria × bohemica Chrtek et Chrtková Seems Not to Rely Heavily on Allelopathy for Its Persistence in Invaded Sites in the Southwest Part of the Zagreb, Croatia" Plants 12, no. 11: 2222. https://doi.org/10.3390/plants12112222