

Identification and Expression Analysis of the Isopentenyl Transferase (IPT) Gene Family under Lack of Nitrogen Stress in Oilseed (Brassica napus L.)


Abstract
:1. Introduction
2. Results and Analysis
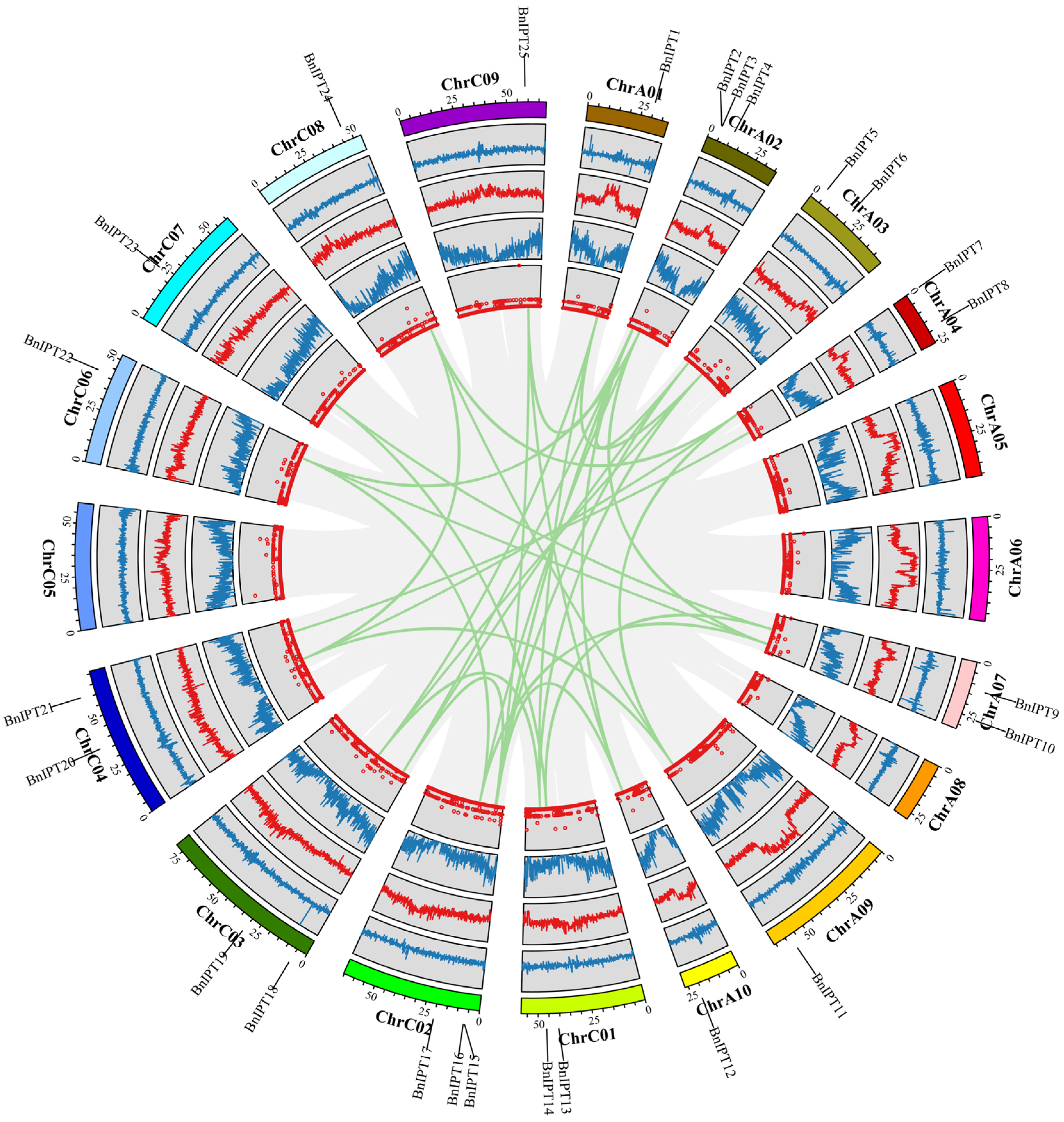
2.1. Identification and Chromosomal Localization of the BnIPT Gene Family
2.2. Prediction of Physicochemical Properties, Structure, and Subcellular Localization of BnIPT Protein
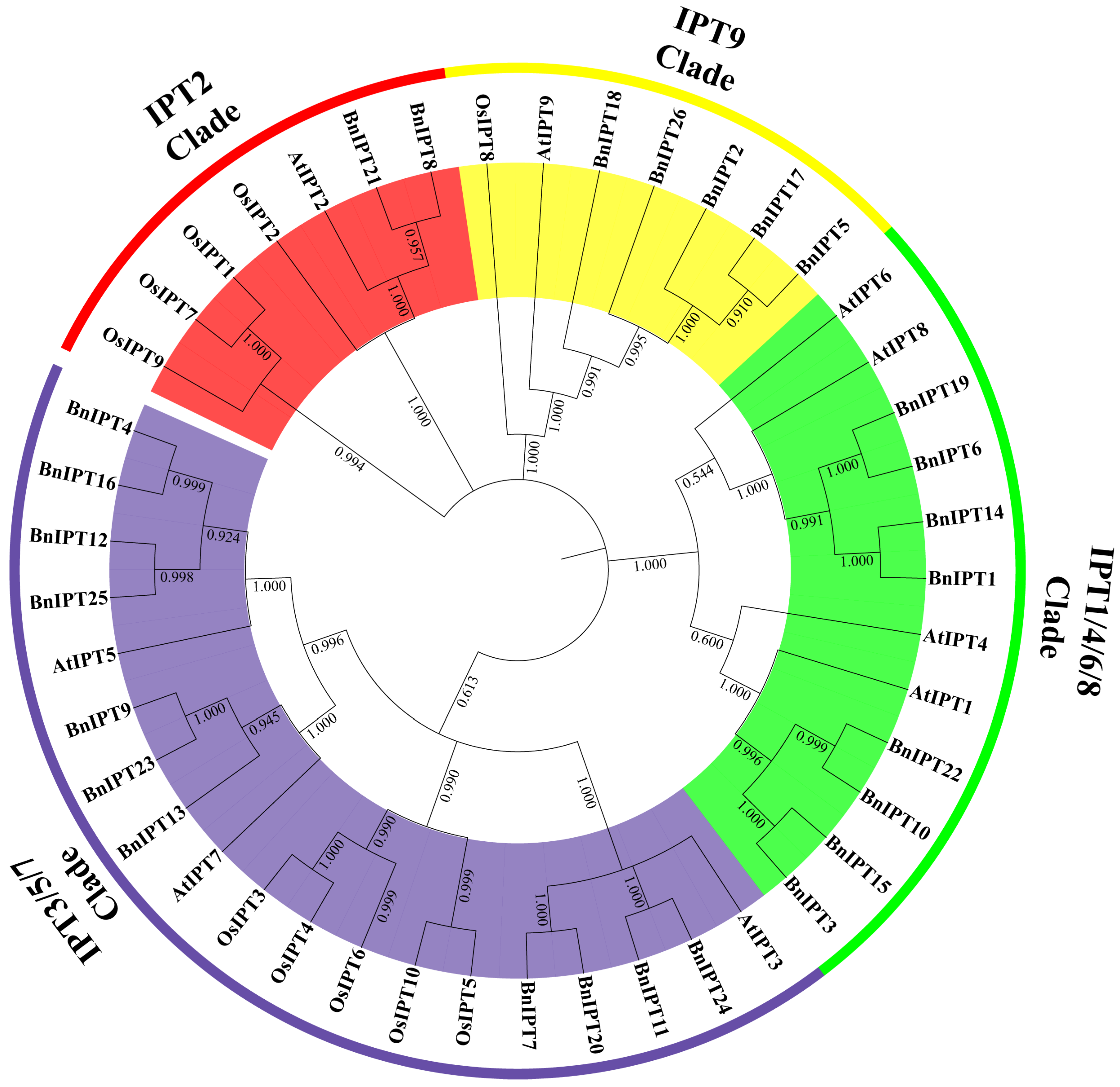
2.3. Phylogenetic Analysis of the IPT Family in B. napus
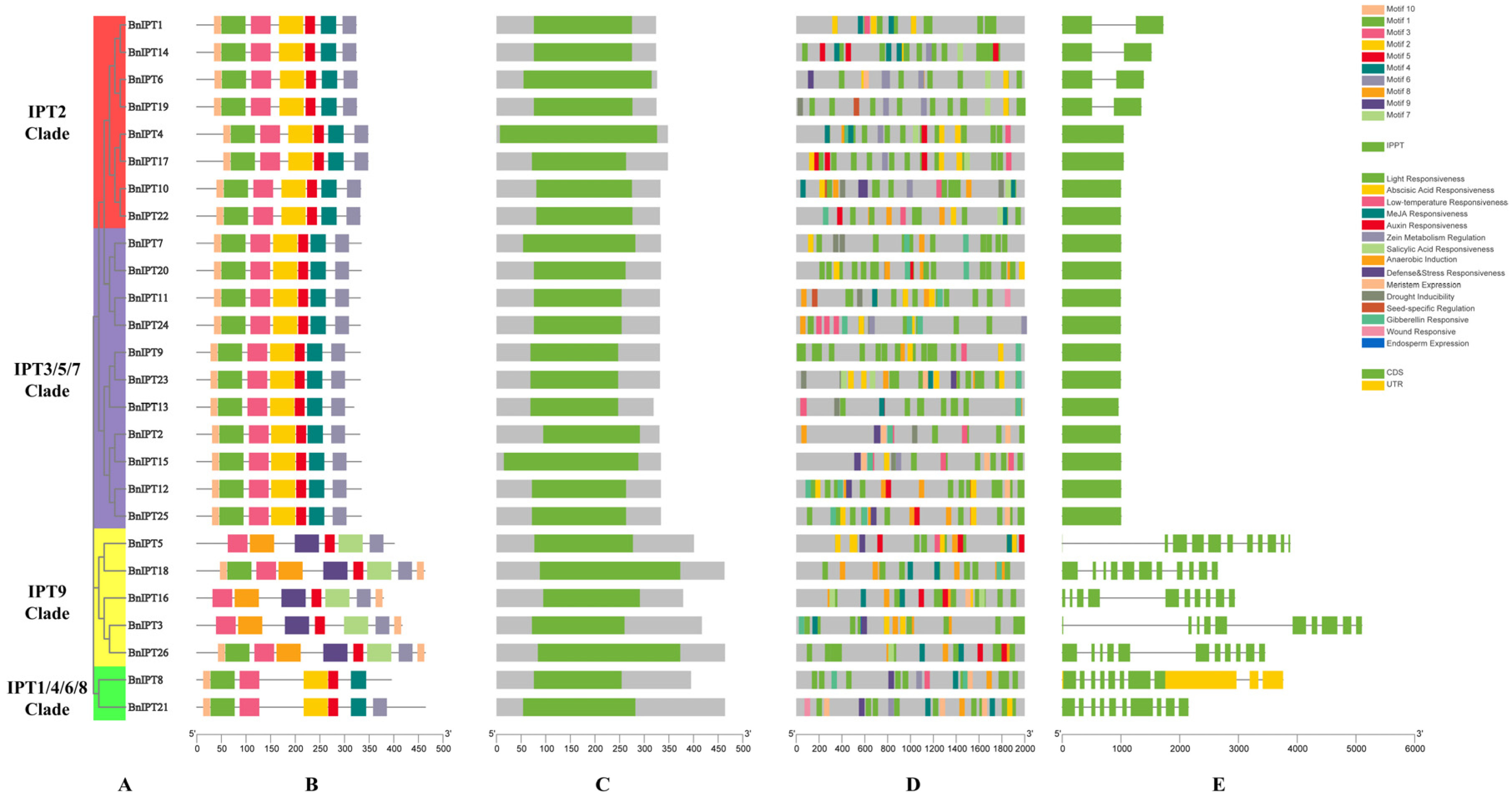
2.4. Analysis of Rape IPT Family Structure, Conserved Motifs, and Promoter Cis-Acting Elements
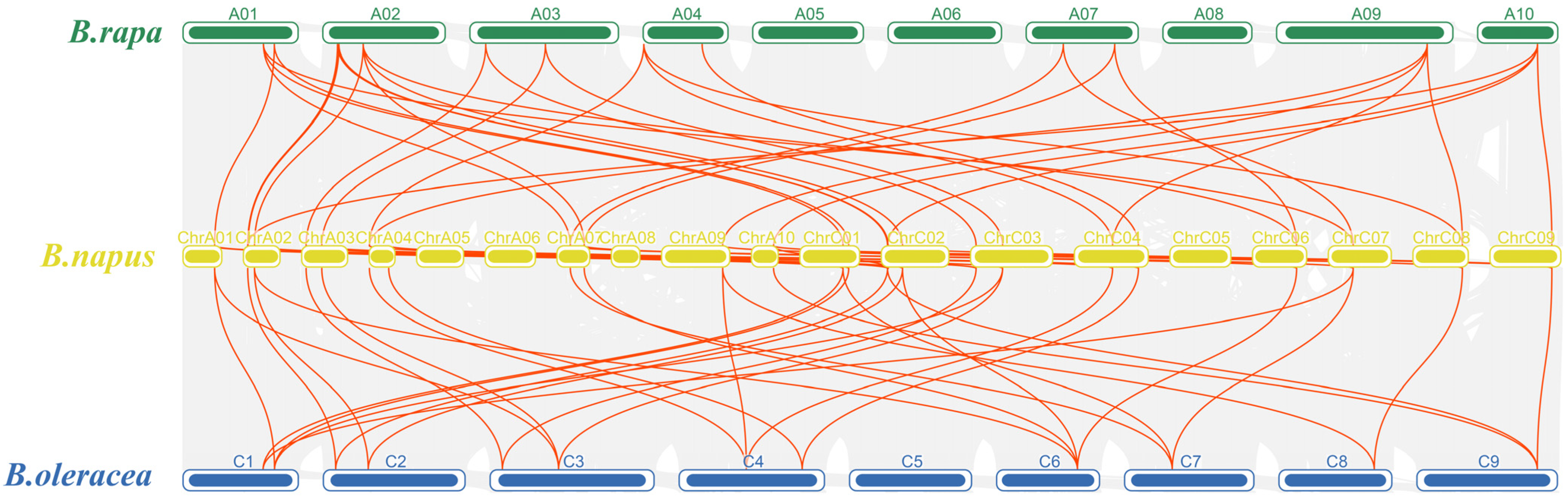
2.5. Collinearity Analysis of IPTs
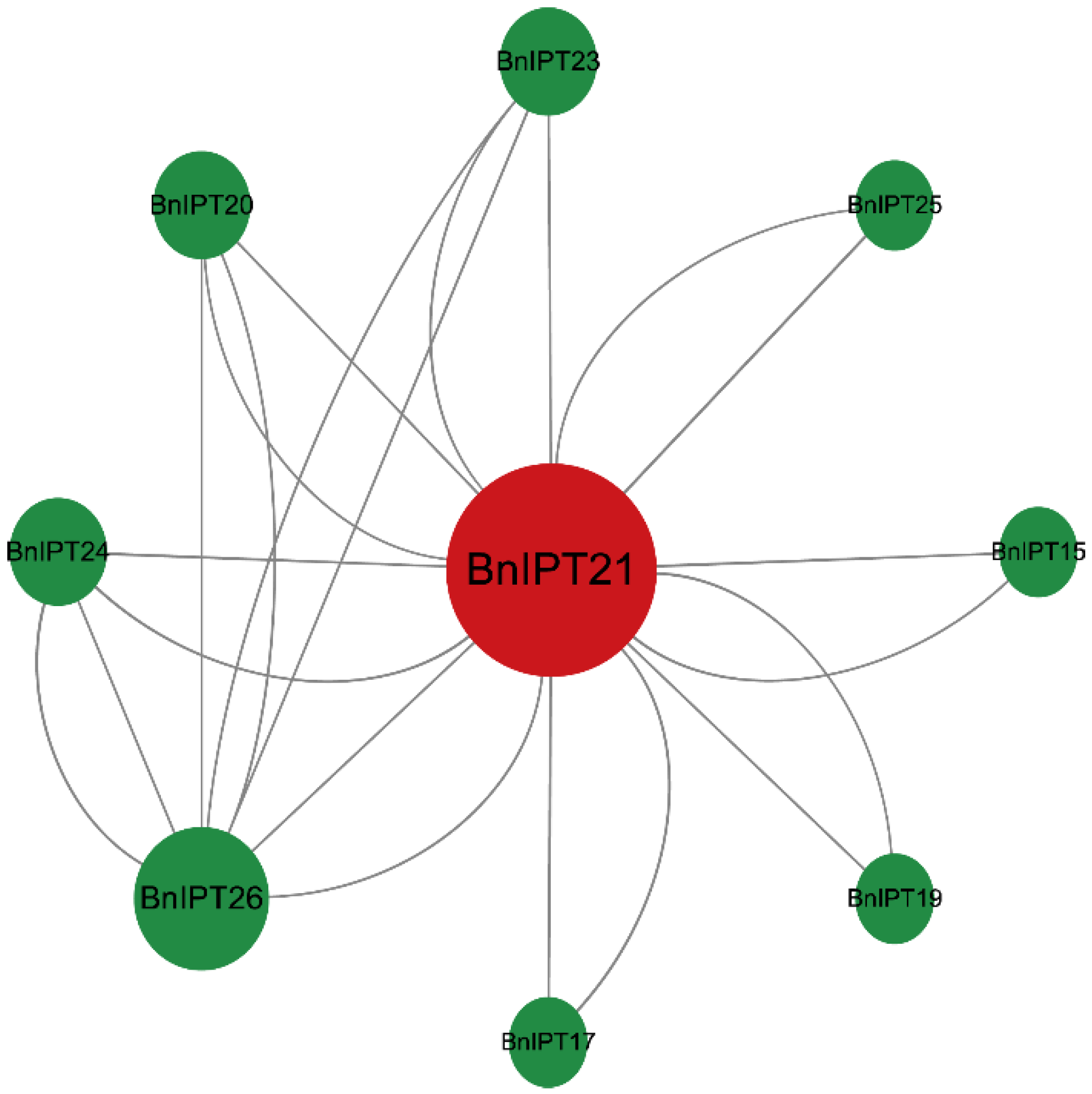
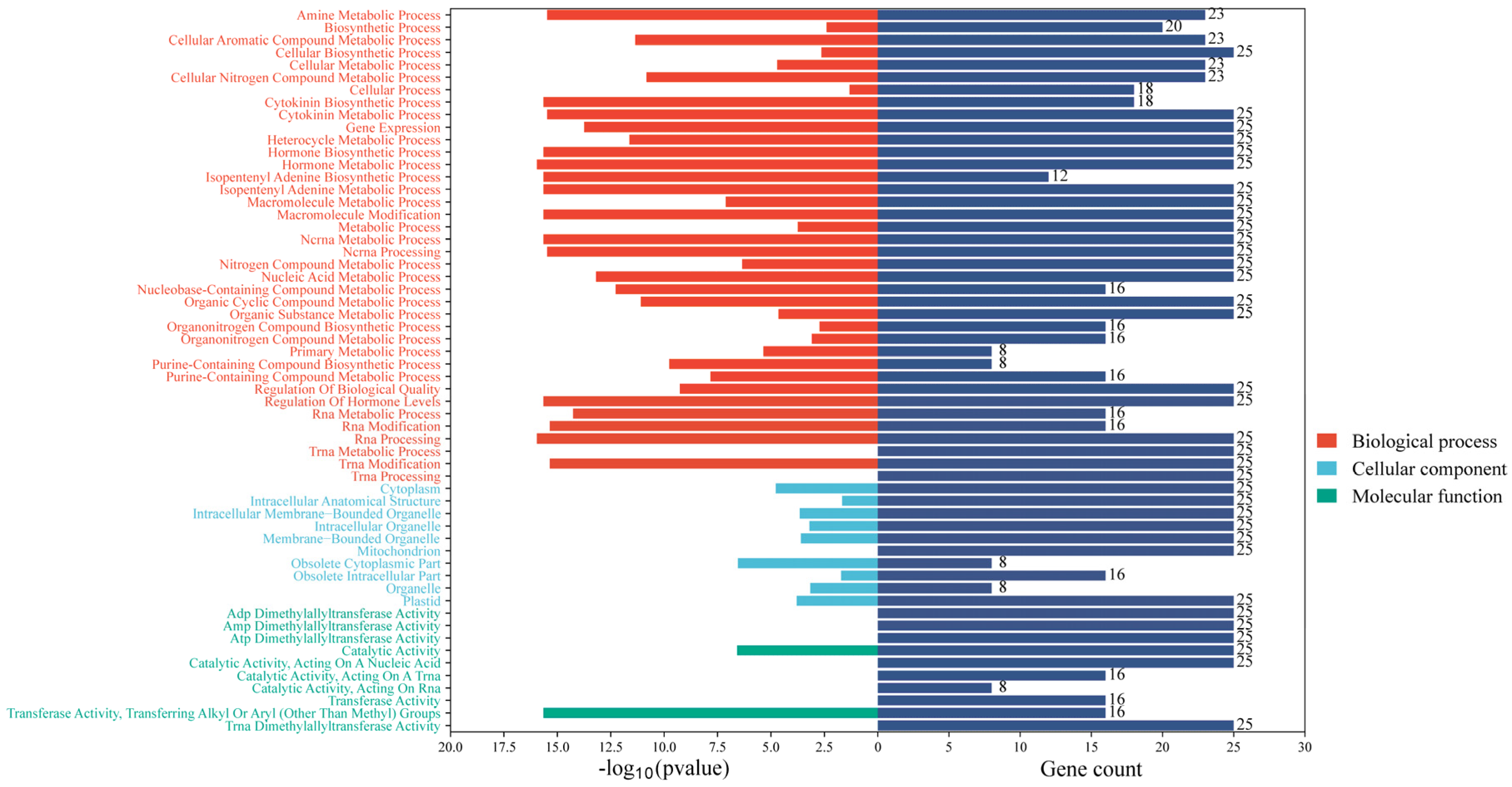
2.6. Protein–Protein Interaction Networks Analysis and GO Enrichment
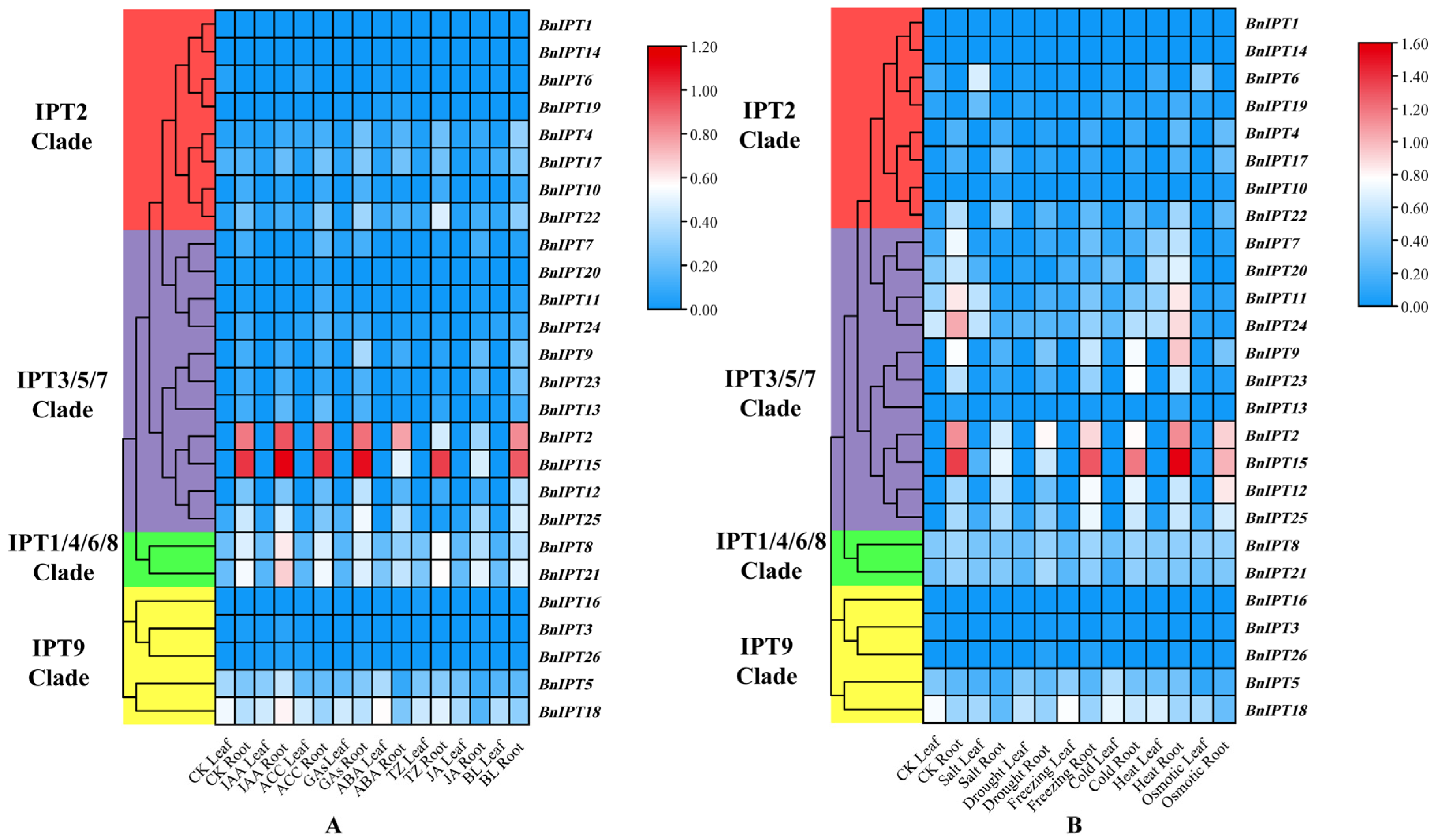
2.7. BnIPT Transcriptome Expression Pattern Analysis
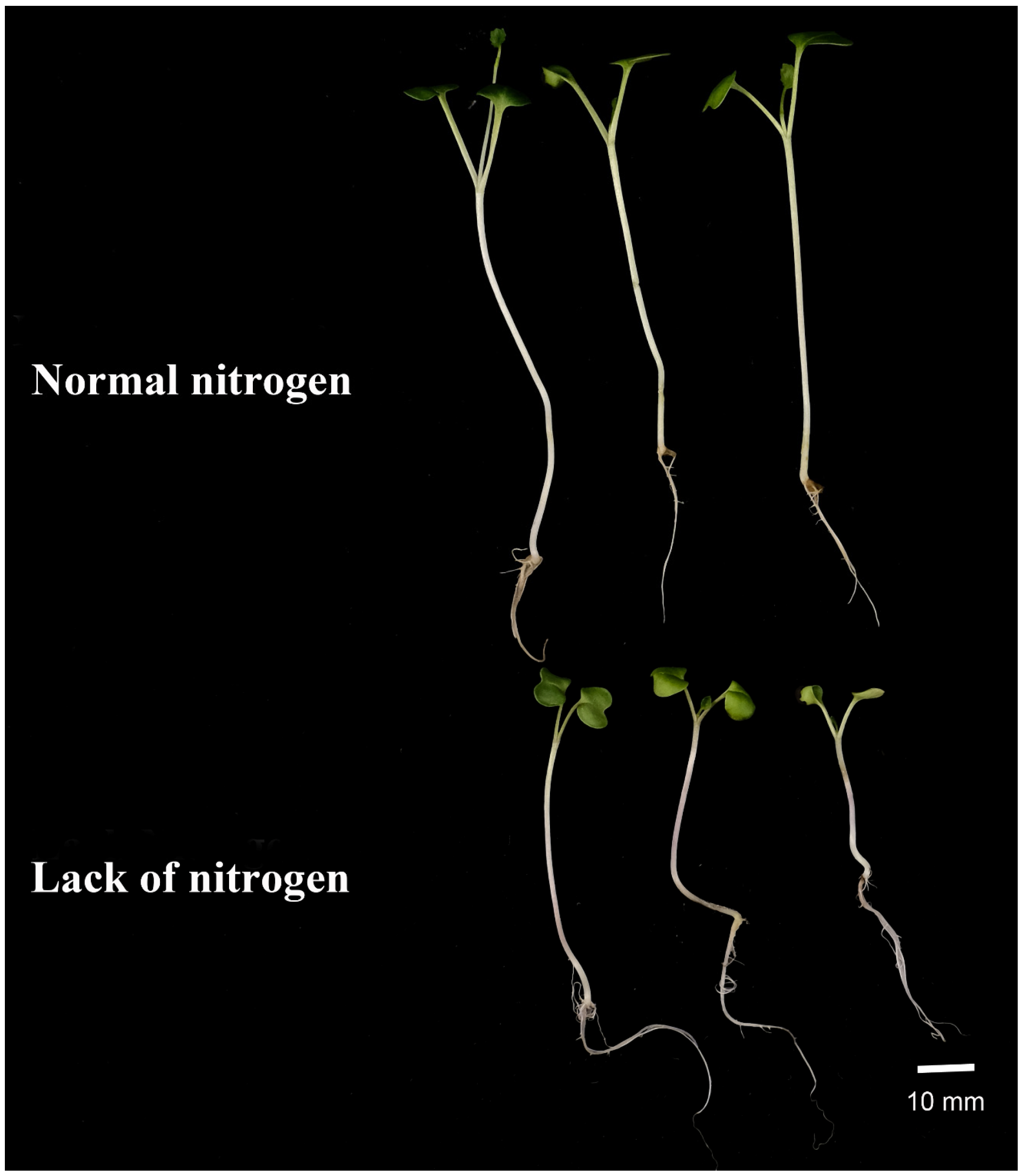
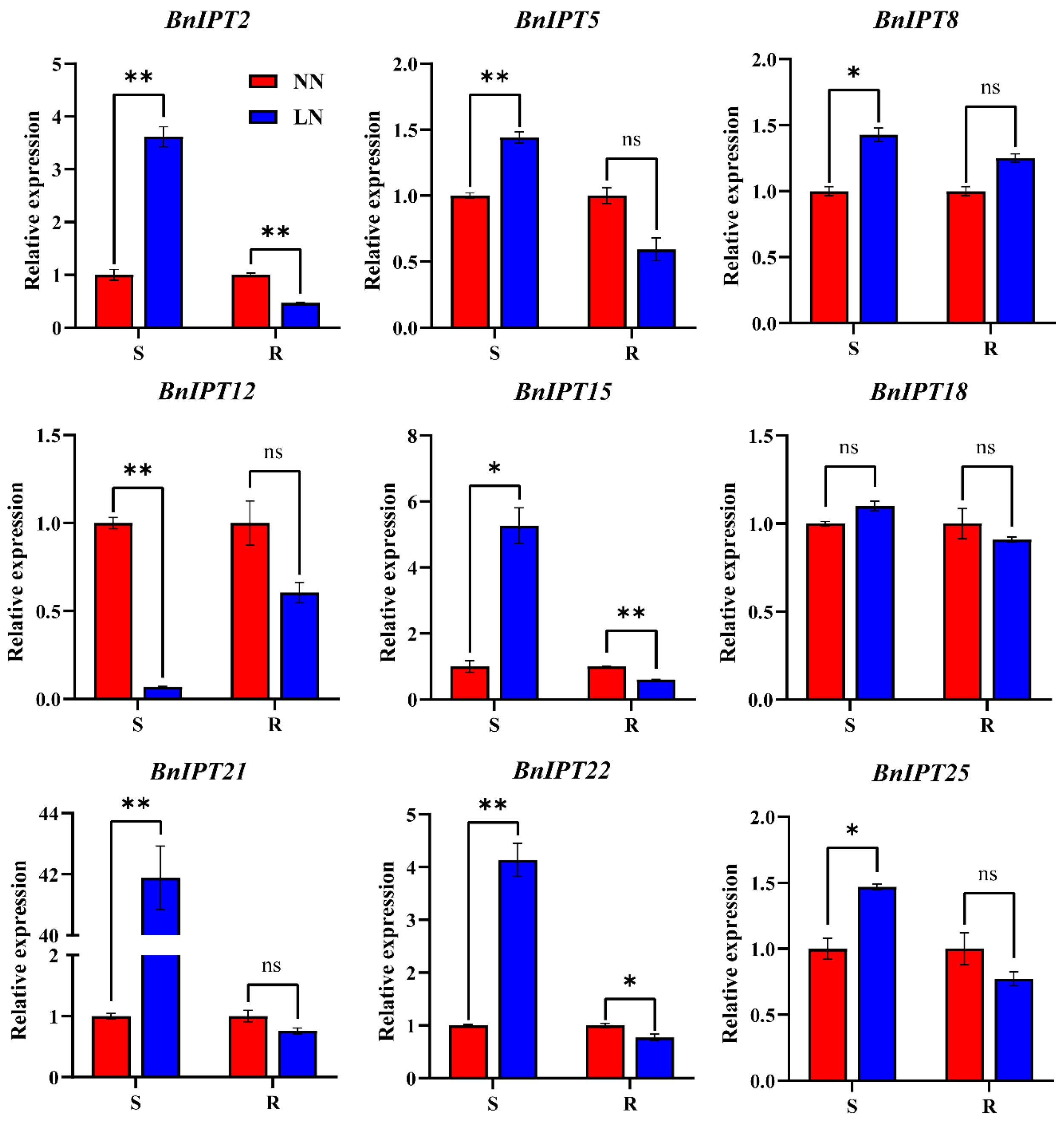
2.8. Expression Analysis of BnIPTs in Response to Lack of Nitrogen
3. Materials and Methods
3.1. Identification of BnIPT Family Members and Chromosome Mapping
3.2. Phylogenetic Analysis
3.3. Prediction of Gene Structures, Protein Conserved Motifs, and Promoter Cis-Acting Elements
3.4. IPT Genes Collinearity Analysis
3.5. Protein–Protein Interaction Networks and GO Enrichment Analysis of BnIPTs
3.6. Transcriptome Expression Pattern Analysis
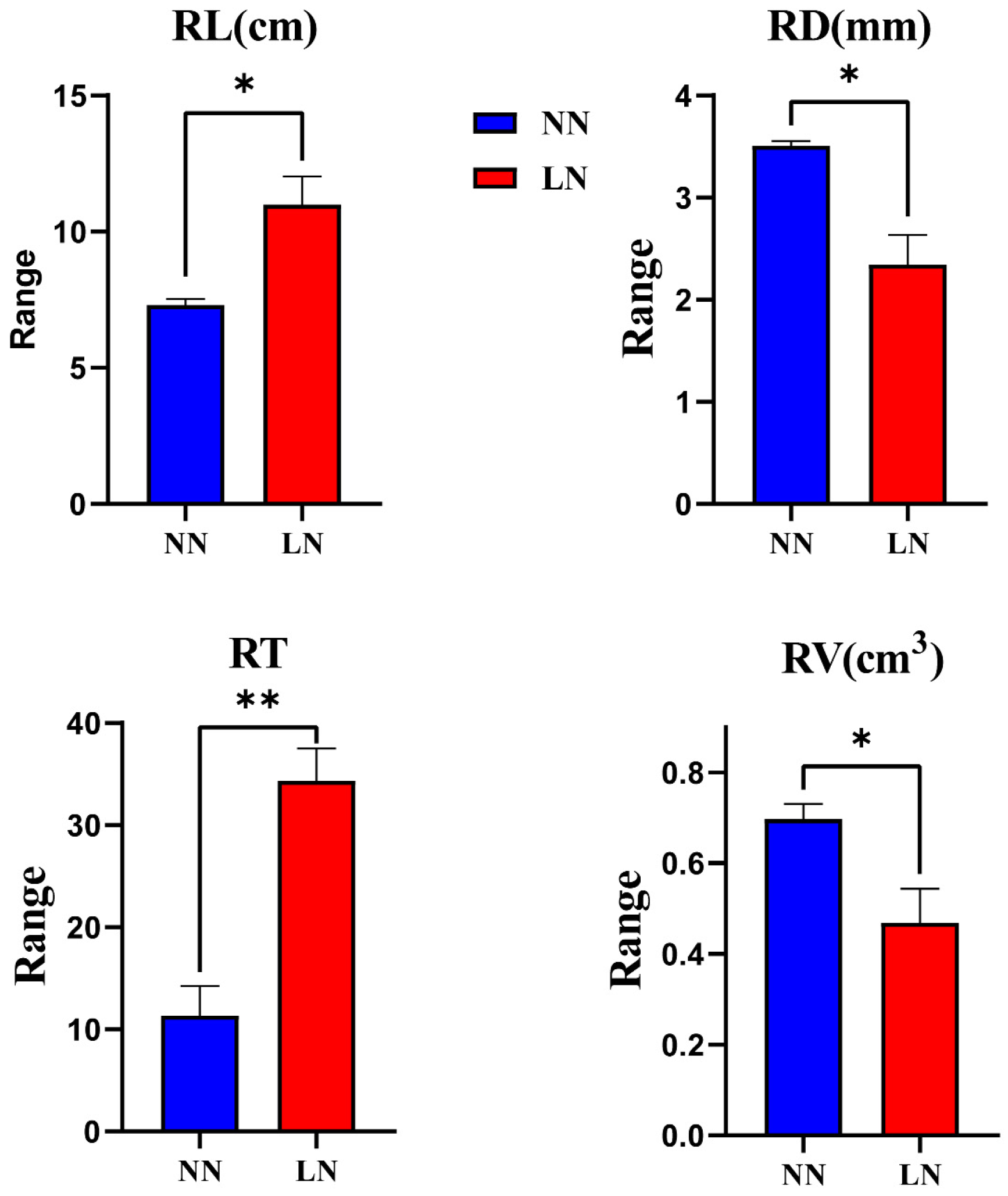
3.7. Plants Materials and Treatment
3.8. Relative Expression of BnIPT Genes under Different Nitrogen Levels
3.9. Statistical Analysis
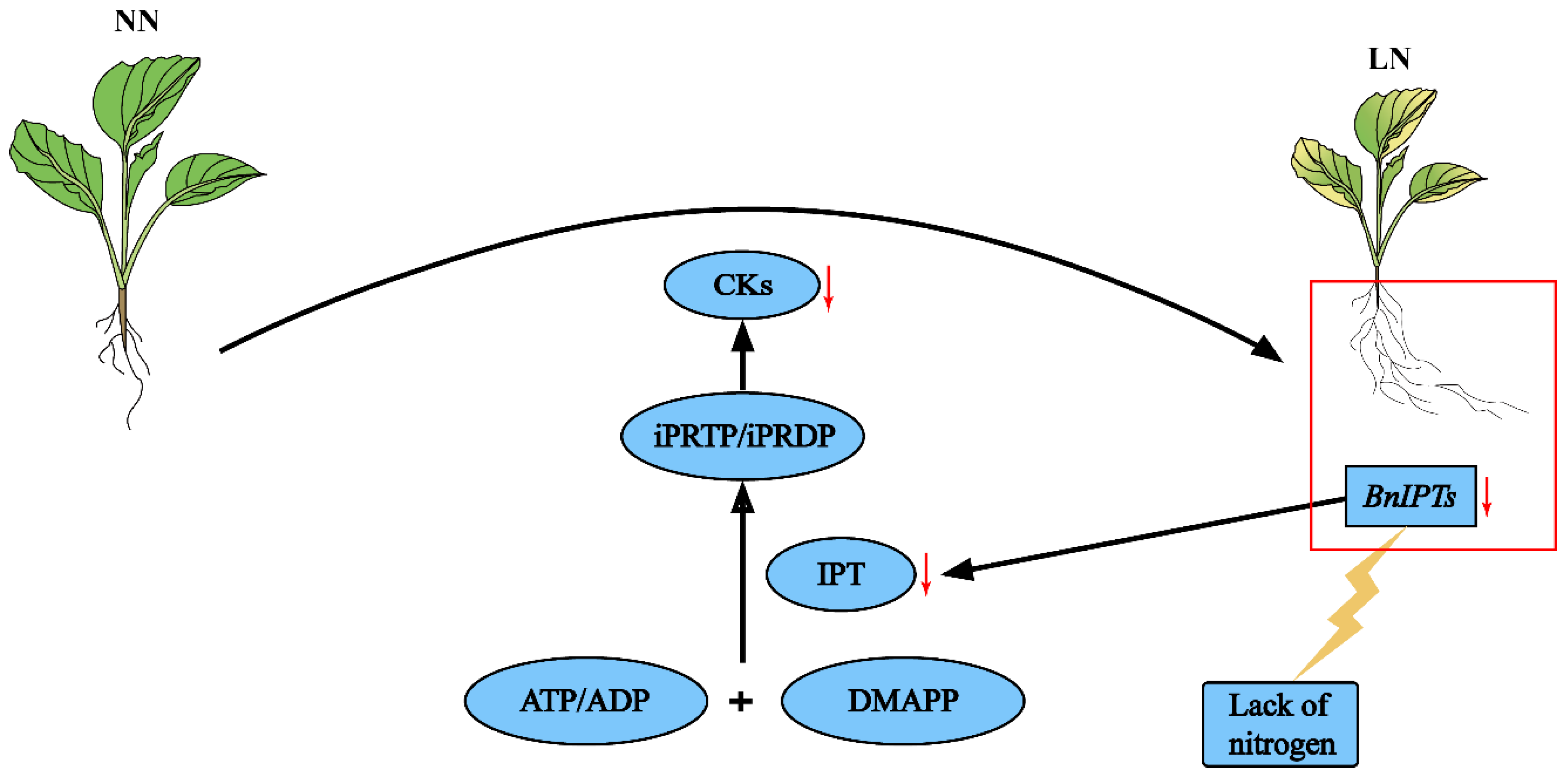
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waadt, R. Phytohormone signaling mechanisms and genetic methods for their modulation and detection. Curr. Opin. Plant. Biol. 2020, 57, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [PubMed]
- Cortleven, A.; Leuendorf, J.E.; Frank, M.; Pezzetta, D.; Bolt, S.; Schmulling, T. Cytokinin action in response to abiotic and biotic stresses in plants. Plant Cell Environ. 2019, 42, 998–1018. [Google Scholar] [CrossRef] [PubMed]
- Mik, V.; Szucova, L.; Spichal, L.; Plihal, O.; Nisler, J.; Zahajska, L.; Dolezal, K.; Strnad, M. N9-Substituted N(6)-[(3-methylbut-2-en-1-yl)amino]purine derivatives and their biological activity in selected cytokinin bioassays. Bioorg. Med. Chem. 2011, 19, 7244–7251. [Google Scholar] [CrossRef]
- Wybouw, B.; De Rybel, B. Cytokinin—A Developing Story. Trends Plant Sci. 2019, 24, 177–185. [Google Scholar] [CrossRef]
- Golovko, A.; Sitbon, F.; Tillberg, E.; Nicander, B. Identification of a tRNA isopentenyltransferase gene from Arabidopsis thaliana. Plant Mol. Biol. 2002, 49, 161–169. [Google Scholar] [CrossRef]
- Gajdosova, S.; Spichal, L.; Kaminek, M.; Hoyerova, K.; Novak, O.; Dobrev, P.I.; Galuszka, P.; Klima, P.; Gaudinova, A.; Zizkova, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef]
- Hnatuszko-Konka, K.; Gerszberg, A.; Weremczuk-Jezyna, I.; Grzegorczyk-Karolak, I. Cytokinin Signaling and De Novo Shoot Organogenesis. Genes 2021, 12, 265. [Google Scholar] [CrossRef]
- Takei, K.; Sakakibara, H.; Sugiyama, T. Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 26405–26410. [Google Scholar] [CrossRef]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases. Plant Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Zhang, J.; He, Y.; Zhu, X.; Zhou, X.; Gong, H.; Yin, J.; Liu, Y. Silicon confers cucumber resistance to salinity stress through regulation of proline and cytokinins. Plant Physiol. Biochem. 2020, 156, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Beznec, A.; Faccio, P.; Miralles, D.J.; Abeledo, L.G.; Oneto, C.D.; Garibotto, M.B.; Gonzalez, G.; Moreyra, F.; Elizondo, M.; Ruiz, M.; et al. Stress-induced expression of IPT gene in transgenic wheat reduces grain yield penalty under drought. J. Genet. Eng. Biotechnol. 2021, 19, 67. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gianfagna, T.; Huang, B. Proteomic changes associated with expression of a gene (ipt) controlling cytokinin synthesis for improving heat tolerance in a perennial grass species. J. Exp. Bot. 2010, 61, 3273–3289. [Google Scholar] [CrossRef] [PubMed]
- Prerostova, S.; Cerny, M.; Dobrev, P.I.; Motyka, V.; Hluskova, L.; Zupkova, B.; Gaudinova, A.; Knirsch, V.; Janda, T.; Brzobohaty, B.; et al. Light Regulates the Cytokinin-Dependent Cold Stress Responses in Arabidopsis. Front. Plant Sci. 2020, 11, 608711. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinin biosynthesis and transport for systemic nitrogen signaling. Plant J. 2021, 105, 421–430. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Zhou, Y.J.; Li, C.Q.; Zhu, K.Y.; Xu, Y.J.; Wang, W.L.; Liu, L.J.; Zhang, H.; Gu, J.F.; Wang, Z.Q.; et al. Post-anthesis moderate soil-drying facilitates source-to-sink remobilization of nitrogen via redistributing cytokinins in rice. Field Crops Res. 2022, 288, 108692. [Google Scholar] [CrossRef]
- Shibasaki, K.; Takebayashi, A.; Makita, N.; Kojima, M.; Takebayashi, Y.; Kawai, M.; Hachiya, T.; Sakakibara, H. Nitrogen Nutrition Promotes Rhizome Bud Outgrowth via Regulation of Cytokinin Biosynthesis Genes and an Oryza longistaminata Ortholog of FINE CULM 1. Front. Plant Sci. 2021, 12, 670101. [Google Scholar] [CrossRef]
- Beszterda, M.; Nogala-Kałucka, M. Current Research Developments on the Processing and Improvement of the Nutritional Quality of Rapeseed (Brassica napus L.). Eur. J. Lipid Sci. Technol. 2019, 121, 1800045. [Google Scholar] [CrossRef]
- Moggré, G.-J.; Alayon Marichal, M.; Sowersby, T.; Grosvenor, A.; Gathercole, J.; Moreno, T. Protein Recovery from New Zealand Oil Rapeseed (Brassica napus) Cake. Waste Biomass Valorization 2021, 13, 1135–1141. [Google Scholar] [CrossRef]
- Li, J.; Han, Z.; Xian, M. Exploration and application of agriculture-tourism technologies based on rape flowers in rural revitalization of China. Oil Crop Sci. 2022, 7, 122–126. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Akhtar, M.; Mukhtar, Z.; Saeed, N.A. Hazards of nitrogen fertilizers and ways to reduce nitrate accumulation in crop plants. Environ. Sci. Pollut. Res. Int. 2020, 27, 17661–17670. [Google Scholar] [CrossRef] [PubMed]
- Rathke, G.; Behrens, T.; Diepenbrock, W. Integrated nitrogen management strategies to improve seed yield, oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): A review. Agric. Ecosyst. Environ. 2006, 117, 80–108. [Google Scholar] [CrossRef]
- Sieling, K.; Kage, H. Efficient N management using winter oilseed rape. A review. Agron. Sustain. Dev. 2010, 30, 271–279. [Google Scholar] [CrossRef]
- Liu, Q.; Ren, T.; Zhang, Y.; Li, X.; Cong, R.; White, P.J.; Lu, J. Yield loss of oilseed rape (Brassica napus L.) under nitrogen deficiency is associated with under-regulation of plant population density. Eur. J. Agron. 2019, 103, 80–89. [Google Scholar] [CrossRef]
- Paul, P.; Aithal, P.; Ripu Ranjan Sinha, R.R.S.; Saavedra, M.R.; Aremu, P.S.B. Agro Informatics with its Various Attributes and Emergence: Emphasizing Potentiality as a Specialization in Agricultural Sciences—A Policy Framework. IRA Int. J. Appl. Sci. 2019, 14, 34–44. [Google Scholar] [CrossRef]
- Ton, L.B.; Neik, T.X.; Batley, J.J.G. The use of genetic and gene technologies in shaping modern rapeseed cultivars (Brassica napus L.). Genes 2020, 11, 1161. [Google Scholar] [CrossRef]
- Chen, H.; Wang, T.; He, X.; Cai, X.; Lin, R.; Liang, J.; Wu, J.; King, G.; Wang, X. BRAD V3.0: An upgraded Brassicaceae database. Nucleic Acids Res. 2022, 50, D1432–D1441. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Combet, C.; Blanchet, C.; Geourjon, C.; Deleage, G. NPS@: Network Protein Sequence Analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Nystrom, S.L.; McKay, D.J. Memes: A motif analysis environment in R using tools from the MEME Suite. PLoS Comput. Biol. 2021, 17, e1008991. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Zhang, Z. KaKs_Calculator 3.0: Calculating Selective Pressure on Coding and Non-coding Sequences. Genom. Proteom. Bioinform. 2022, 20, 536–540. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Hernandez-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Liu, D.; Yu, L.; Wei, L.; Yu, P.; Wang, J.; Zhao, H.; Zhang, Y.; Zhang, S.; Yang, Z.; Chen, G.; et al. BnTIR: An online transcriptome platform for exploring RNA-seq libraries for oil crop Brassica napus. Plant Biotechnol. J. 2021, 19, 1895–1897. [Google Scholar] [CrossRef]
- Wang, G.L.; Ding, G.D.; Xu, F.S.; Cai, H.M.; Zou, J.; Ye, X.S. Genotype differences in photosynthetic characteristics and nitrogen efficiency of new-type oilseed rape responding to low nitrogen stress. J. Agric. Sci. 2014, 153, 1030–1043. [Google Scholar] [CrossRef]
- Song, J.; Jiang, L.; Jameson, P.E. Expression patterns of Brassica napus genes implicate IPT, CKX, sucrose transporter, cell wall invertase, and amino acid permease gene family members in leaf, flower, silique, and seed development. J. Exp. Bot. 2015, 66, 5067–5082. [Google Scholar] [CrossRef]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Fan, G.; Hu, Q.; Zhou, Y.; Guan, M.; Tong, C.; Li, J.; Du, D.; Qi, C.; Jiang, L.; et al. The high-quality genome of Brassica napus cultivar ‘ZS11’ reveals the introgression history in semi-winter morphotype. Plant J. 2017, 92, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Vankova, R.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS ONE 2012, 7, e42411. [Google Scholar] [CrossRef]
- Ghosh, A.; Shah, M.N.A.; Jui, Z.S.; Saha, S.; Fariha, K.A.; Islam, T. Evolutionary variation and expression profiling of Isopentenyl transferase gene family in Arabidopsis thaliana L. and Oryza sativa L. Plant Gene 2018, 15, 15–27. [Google Scholar] [CrossRef]
- Tan, M.; Li, G.; Qi, S.; Liu, X.; Chen, X.; Ma, J.; Zhang, D.; Han, M. Identification and expression analysis of the IPT and CKX gene families during axillary bud outgrowth in apple (Malus domestica Borkh.). Gene 2018, 651, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Soleymani, F.; Paquet, E.; Viktor, H.; Michalowski, W.; Spinello, D. Protein-protein interaction prediction with deep learning: A comprehensive review. Comput. Struct. Biotechnol. J. 2022, 20, 5316–5341. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Hao, Z.G.; Miao, S.; Zhang, X.; Li, J.Q.; Guo, S.X.; Lee, Y.I. Profiles of Cytokinins Metabolic Genes and Endogenous Cytokinins Dynamics during Shoot Multiplication In Vitro of Phalaenopsis. Int. J. Mol. Sci. 2022, 23, 3755. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, F.; Su, Y.; Jiang, Q.; Yuan, Y.; Nie, X.; Zhou, Y.; Zhang, X.; Wang, Z.; Wang, F.; et al. MdIPT8, an isopentenyl transferase enzyme, enhances the resistance of apple to Colletotrichum gloeosporioides infection. Sci. Hortic. 2022, 303, 111245. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Lai, N.; Kisiala, A.B.; Emery, R.J.N. Isopentenyltransferases as master regulators of crop performance: Their function, manipulation, and genetic potential for stress adaptation and yield improvement. Plant Biotechnol. J. 2021, 19, 1297–1313. [Google Scholar] [CrossRef]
- Antoniadi, I.; Mateo-Bonmati, E.; Pernisova, M.; Brunoni, F.; Antoniadi, M.; Villalonga, M.G.; Ament, A.; Karady, M.; Turnbull, C.; Dolezal, K.; et al. IPT9, a cis-zeatin cytokinin biosynthesis gene, promotes root growth. Front. Plant Sci. 2022, 13, 932008. [Google Scholar] [CrossRef] [PubMed]
- Glanz-Idan, N.; Lach, M.; Tarkowski, P.; Vrobel, O.; Wolf, S. Delayed Leaf Senescence by Upregulation of Cytokinin Biosynthesis Specifically in Tomato Roots. Front. Plant Sci. 2022, 13, 922106. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Meng, Z.; Wang, B.; Chen, M. Research Progress on the Roles of Cytokinin in Plant Response to Stress. Int. J. Mol. Sci. 2020, 21, 6574. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Peng, L.; Zhang, Y. Plant DNA Methylation Responds to Nutrient Stress. Genes 2022, 13, 992. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Hh | Ee | Cc | Protein Name | Hh | Ee | Cc |
|---|---|---|---|---|---|---|---|
| BnIPT1 | 45.06% | 12.04% | 42.90% | BnIPT14 | 45.68% | 11.42% | 42.90% |
| BnIPT2 | 46.83% | 11.78% | 41.39% | BnIPT15 | 49.40% | 11.98% | 38.62% |
| BnIPT3 | 47.00% | 13.43% | 39.57% | BnIPT16 | 51.45% | 8.44% | 40.11% |
| BnIPT4 | 46.55% | 11.21% | 42.24% | BnIPT17 | 42.82% | 13.79% | 43.39% |
| BnIPT5 | 48.63% | 10.22% | 41.15% | BnIPT18 | 45.36% | 13.61% | 41.04% |
| BnIPT6 | 45.40% | 11.96% | 42.64% | BnIPT19 | 45.85% | 12.31% | 41.85% |
| BnIPT7 | 51.50% | 11.08% | 37.43% | BnIPT20 | 49.40% | 11.98% | 38.62% |
| BnIPT8 | 51.39% | 10.13% | 38.48% | BnIPT21 | 52.80% | 8.84% | 38.36% |
| BnIPT9 | 47.59% | 12.65% | 39.76% | BnIPT22 | 45.48% | 11.45% | 43.07% |
| BnIPT10 | 46.85% | 11.71% | 41.44% | BnIPT23 | 46.69% | 12.35% | 40.96% |
| BnIPT11 | 49.10% | 10.54% | 40.36% | BnIPT24 | 51.51% | 10.24% | 38.25% |
| BnIPT12 | 50.60% | 11.38% | 38.02% | BnIPT25 | 48.80% | 11.08% | 40.12% |
| BnIPT13 | 51.10% | 12.23% | 36.68% | BnIPT26 | 44.61% | 12.07% | 43.32% |
| Chemical Reagent | Mother Liquor (g/L) | Normal Nitrogen Nutrient Solution (mL/L) 1 | Lack of Nitrogen Nutrition Solution (mL/L) |
|---|---|---|---|
| KNO3 | 102.00 | 5.00 | 0 |
| MgSO4·7H2O | 98.00 | 5.00 | 5.00 |
| KH2PO4 | 28.00 | 5.00 | 5.00 |
| Ca(NO3)2·4H2O | 236.00 | 0.50 | 0 |
| FeSO4·7H2O | 2.80 | 5.00 | 5.00 |
| EDTA-2Na | 3.72 | 5.00 | 5.00 |
| MnCl2·4H2O | 3.62 | 0.25 | 0.25 |
| ZnSO4·7H2O | 0.44 | 0.25 | 0.25 |
| CuSO4·5H2O | 0.16 | 0.25 | 0.25 |
| H3BO3 | 5.72 | 0.25 | 0.25 |
| Na2MoO4·4H2O | 0.18 | 0.25 | 0.25 |
| KCl | 75.50 | 0 | 5.00 |
| CaCl2 | 111.00 | 4.50 | 5.00 |
| Gene Name | Forward Primer (5′→3′) | Reverse Primer (3′→5′) | Amplicon Length (bp) |
|---|---|---|---|
| BnIPT2 | CCGACAAGATCCAAGTCTACAA | AAATCCTCGTGAGTGTCTTCC | 118 |
| BnIPT5 | AGGCCTTGATGTTGGATCAG | CCTTGAGAGGGATGCAGAATAT | 90 |
| BnIPT8 | GCGAAGGTGGTCGTGATAAT | TTGCGTCGGCGTTTATGA | 103 |
| BnIPT9 | CTCAACACTCTCGGCTAACAA | AGGAAACGTCGACCCAAATAA | 137 |
| BnIPT12 | GTCTAGACATCGTCACCAACAA | TAATCCTCCGCCGTGAAATC | 109 |
| BnIPT15 | GGAGAACACTCACGACGATTT | AGAATTAGAACCACCGGCTATG | 115 |
| BnIPT18 | GGTGACTAGCAGTAGCGTATTC | TCGAAGTGTTCCCGTTTAGAG | 131 |
| BnIPT21 | CGGAATCGAGGAAGAGAAGATG | GAAGTGAGACGCCAAGTCTAC | 106 |
| BnIPT22 | CCGTCTTTCTCCTCACATTCTC | GCTCCTAAGATGACGACGATTT | 101 |
| BnIPT25 | ACAAAGTCACTCCAGAGGAAAG | AGTGCTTCACGCTGGTAATC | 106 |
| BnActin7 | TGGGTTTGCTGGTGACGAT | TGCCTAGGACGACCAACAATACT | 63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wan, H.; Zhu, W.; Dai, X.; Yu, Y.; Zeng, C. Identification and Expression Analysis of the Isopentenyl Transferase (IPT) Gene Family under Lack of Nitrogen Stress in Oilseed (Brassica napus L.). Plants 2023, 12, 2166. https://doi.org/10.3390/plants12112166
Chen J, Wan H, Zhu W, Dai X, Yu Y, Zeng C. Identification and Expression Analysis of the Isopentenyl Transferase (IPT) Gene Family under Lack of Nitrogen Stress in Oilseed (Brassica napus L.). Plants. 2023; 12(11):2166. https://doi.org/10.3390/plants12112166
Chicago/Turabian StyleChen, Jingdong, Heping Wan, Wenhui Zhu, Xigang Dai, Yi Yu, and Changli Zeng. 2023. "Identification and Expression Analysis of the Isopentenyl Transferase (IPT) Gene Family under Lack of Nitrogen Stress in Oilseed (Brassica napus L.)" Plants 12, no. 11: 2166. https://doi.org/10.3390/plants12112166